
Effective Components and Molecular Mechanism of Agarwood Essential Oil Inhalation and the Sedative and Hypnotic Effects Based on GC-MS-Qtof and Molecular Docking

 ,
,
Abstract
:1. Introduction
2. Results
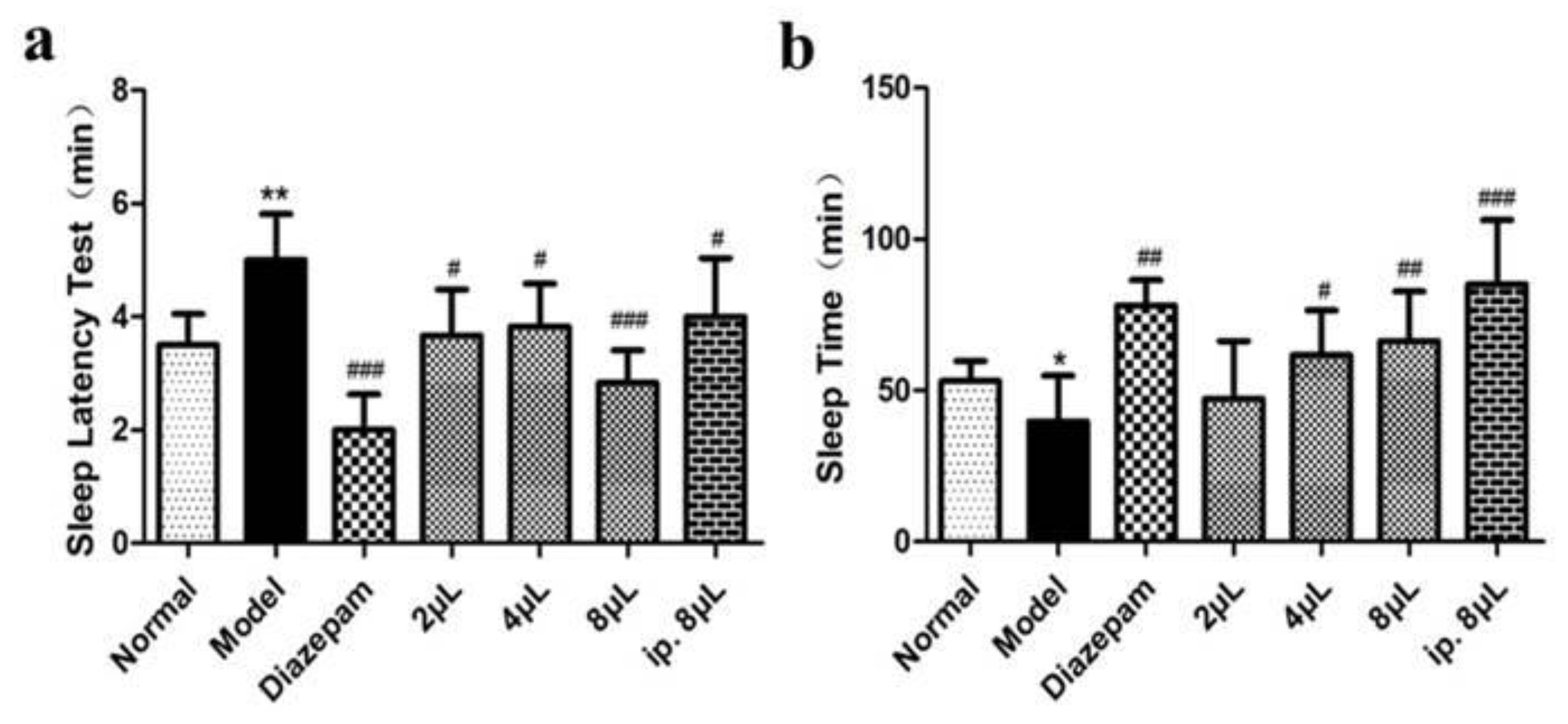
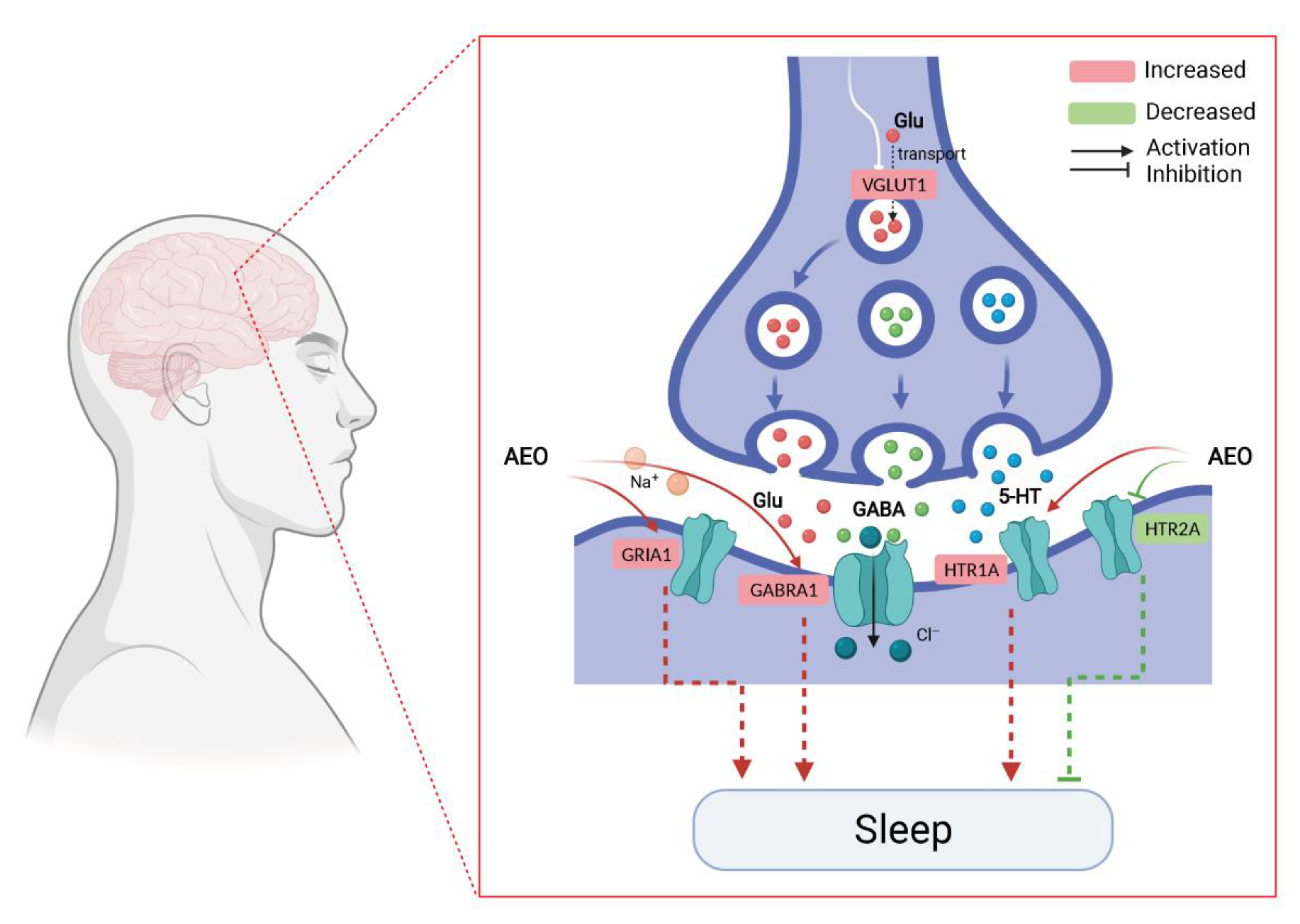
2.1. AEO Affects Sleep
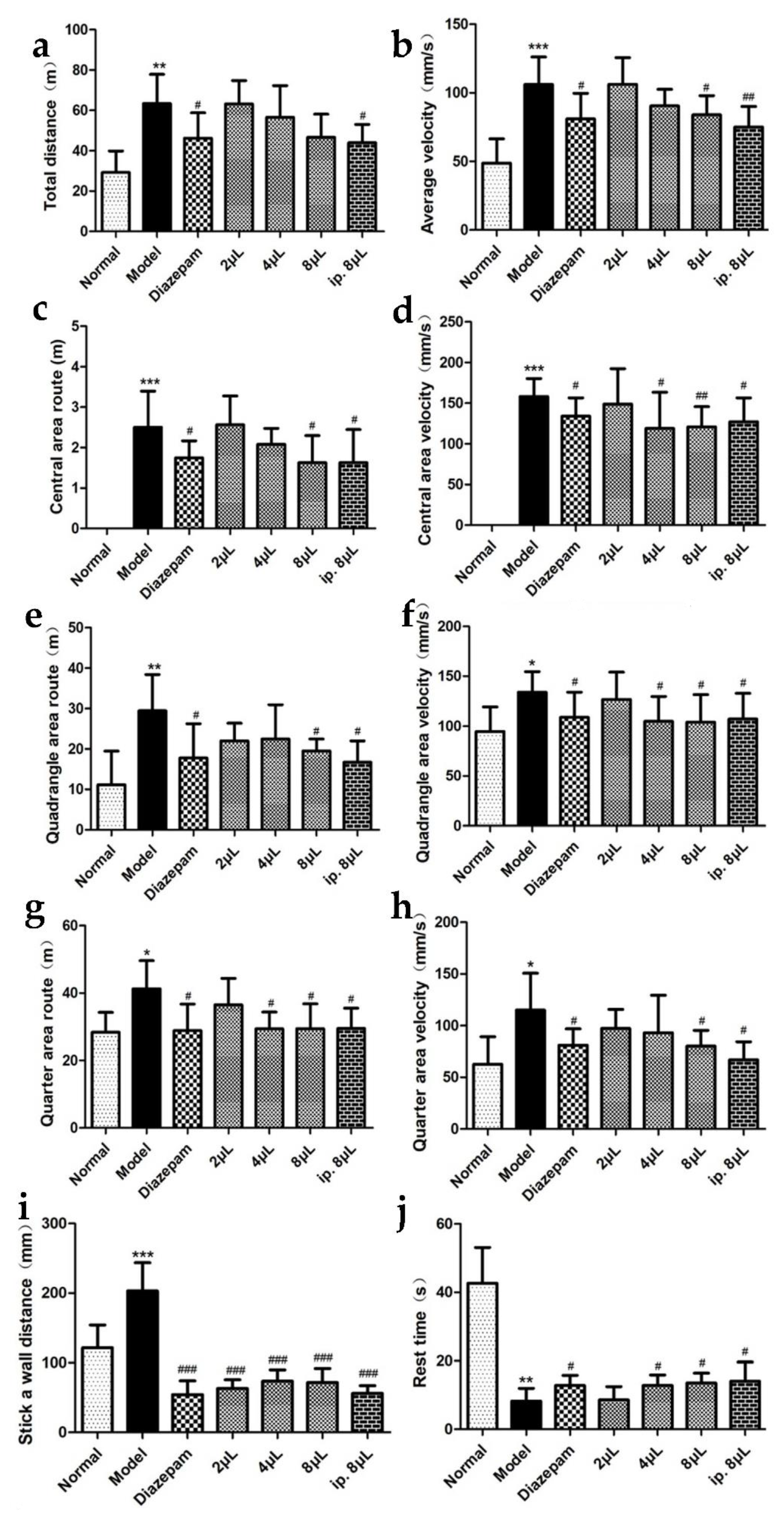
2.2. AEO Affects Autonomous Activities
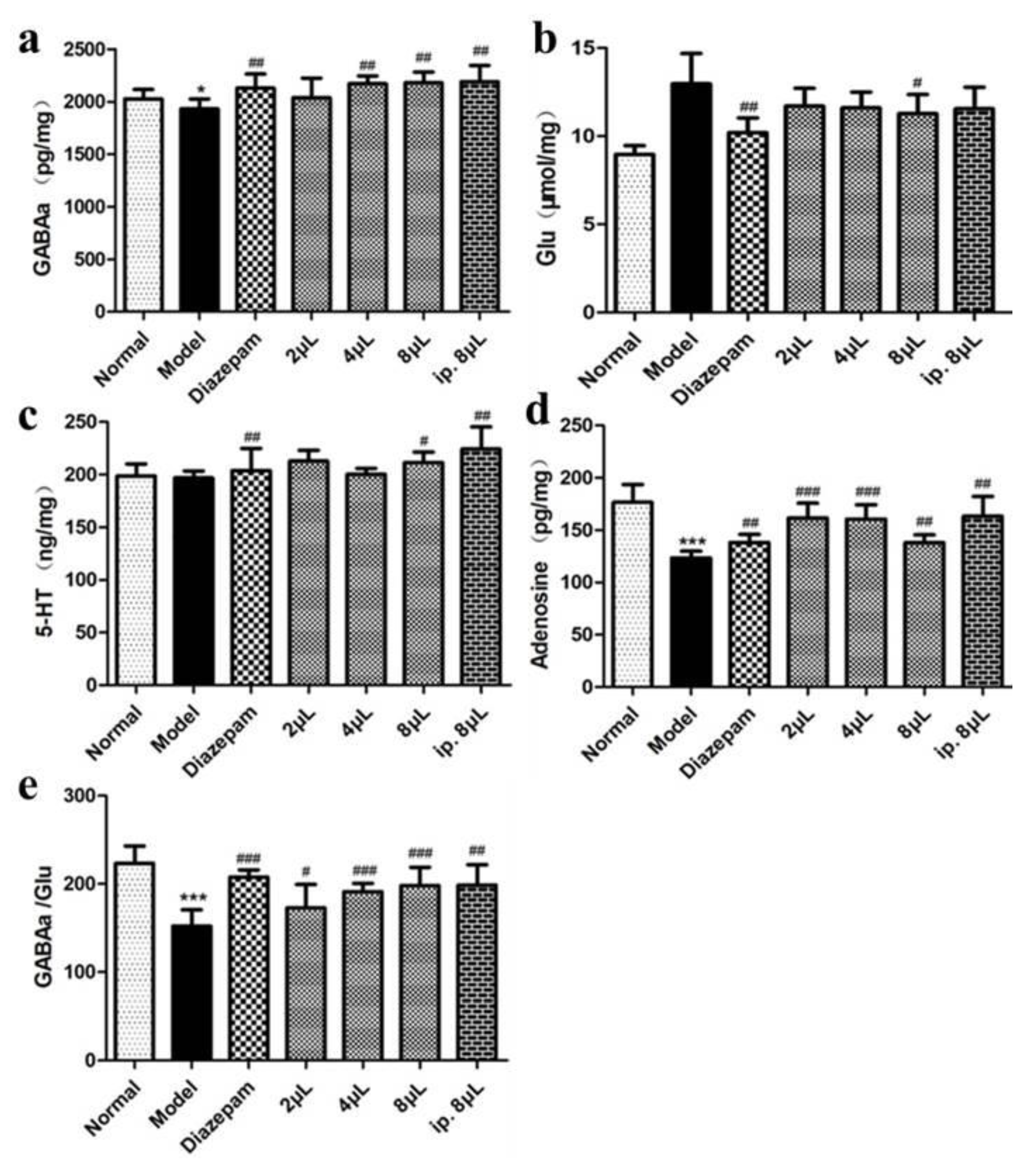
2.3. AEO Affects Neurotransmitter Levels
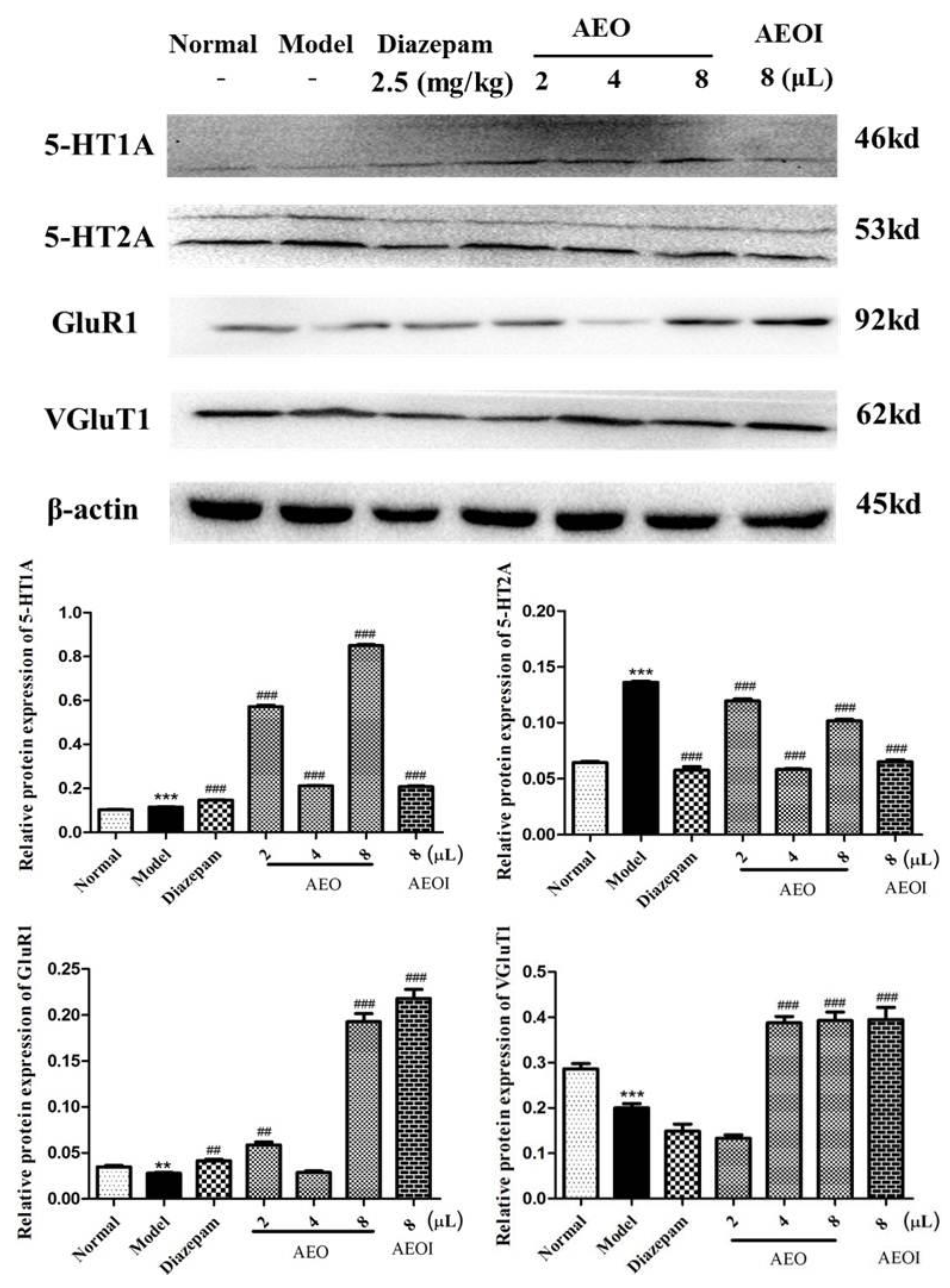
2.4. AEO Affects Protein Expression
2.5. Headspace Solid-Phase Microextraction Optimization
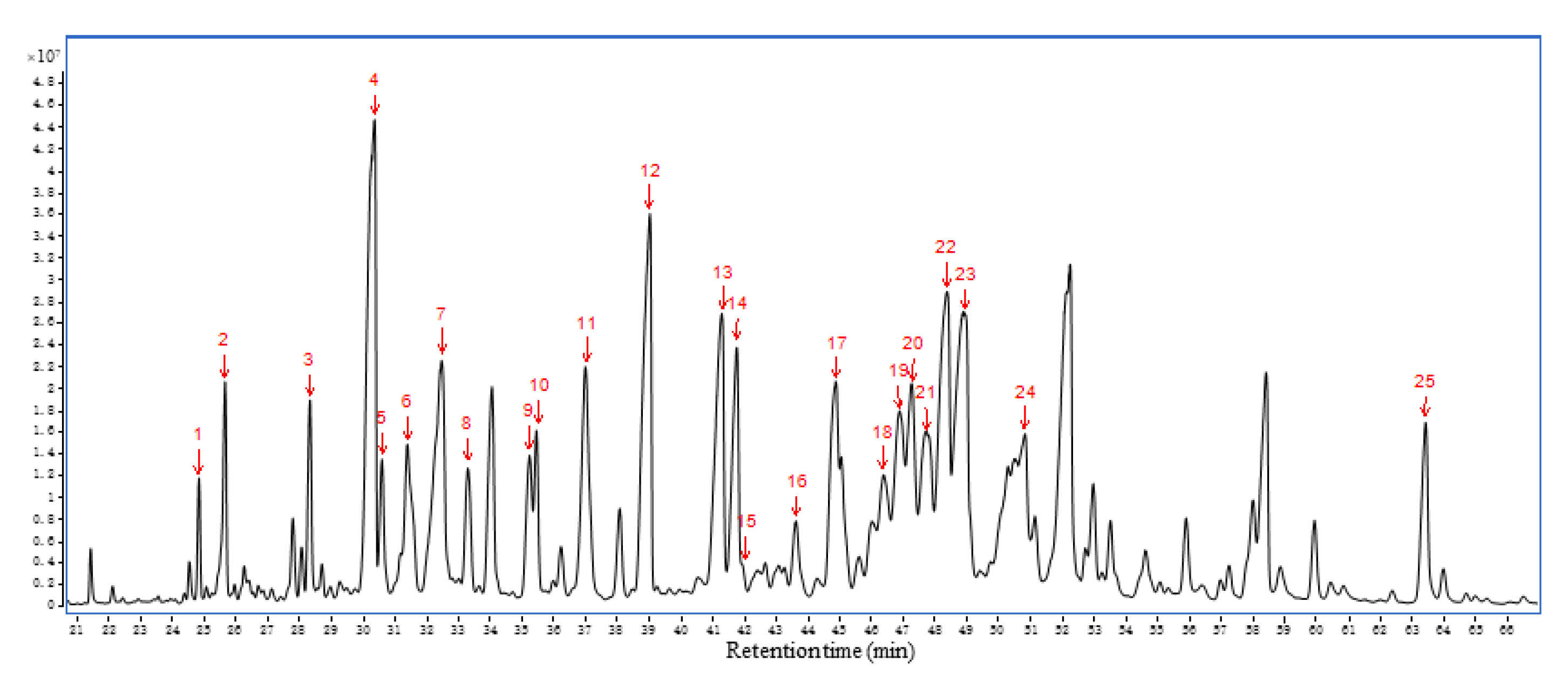
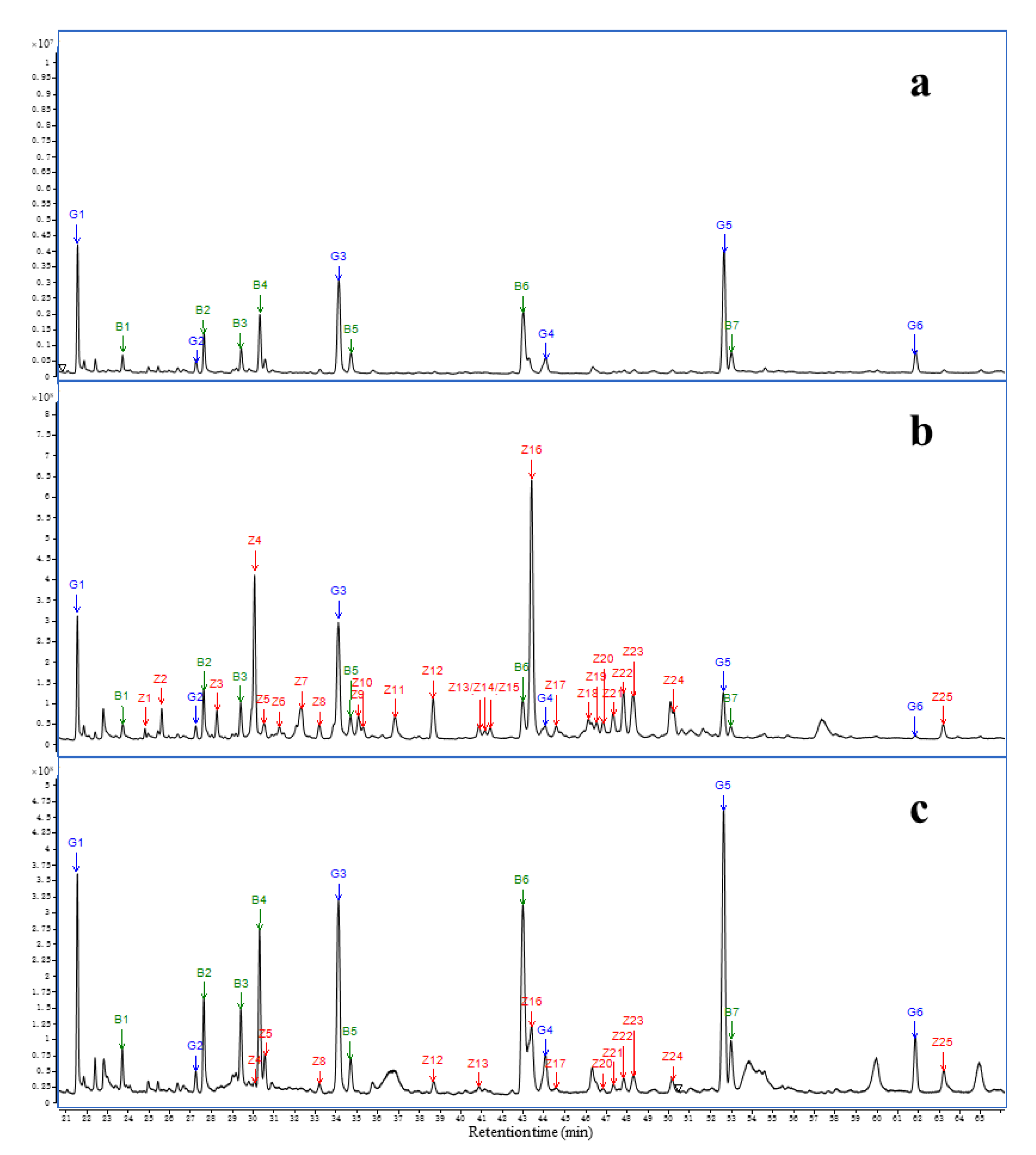
2.6. Chemical Profile of AEO and Serum
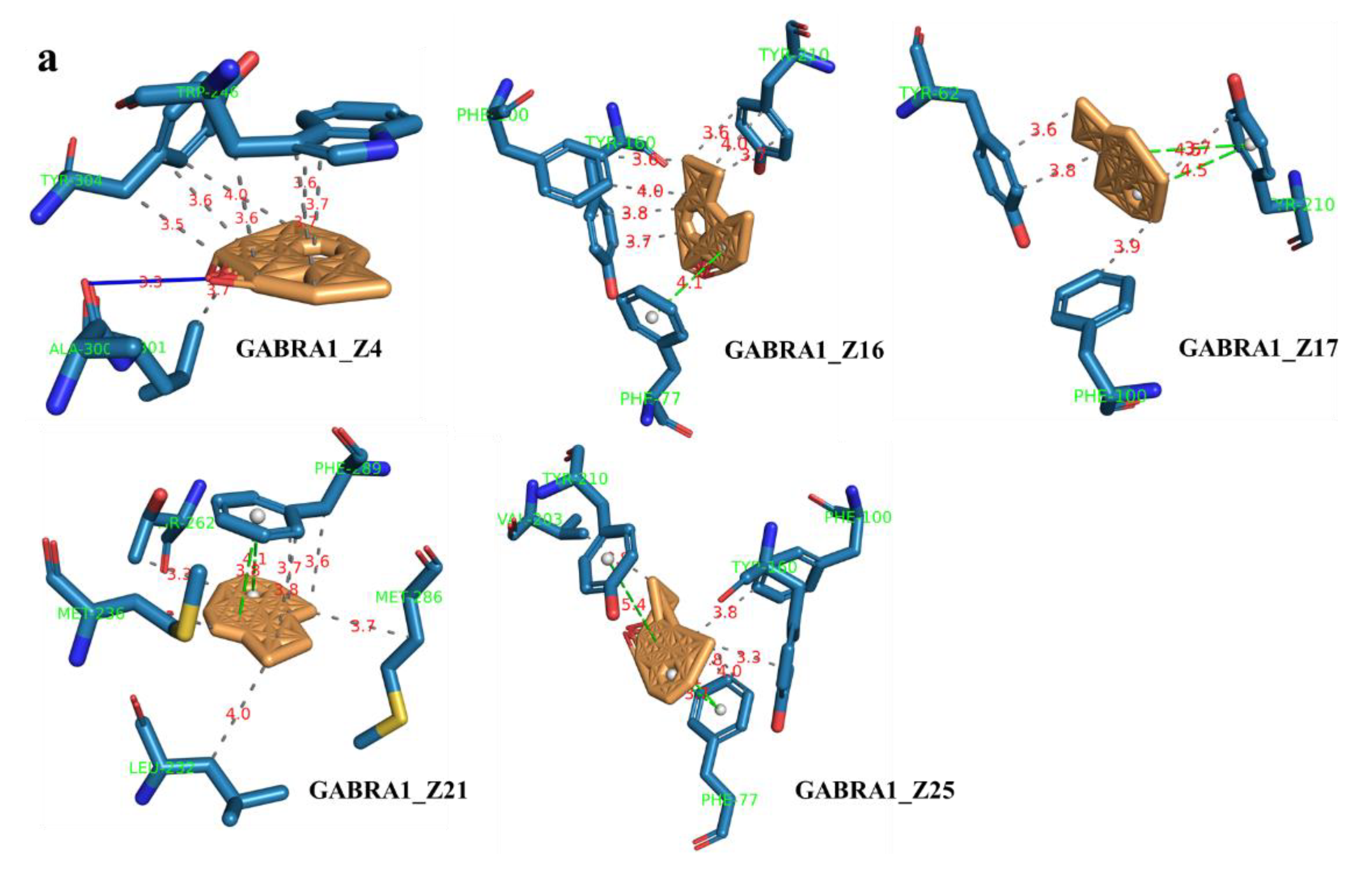
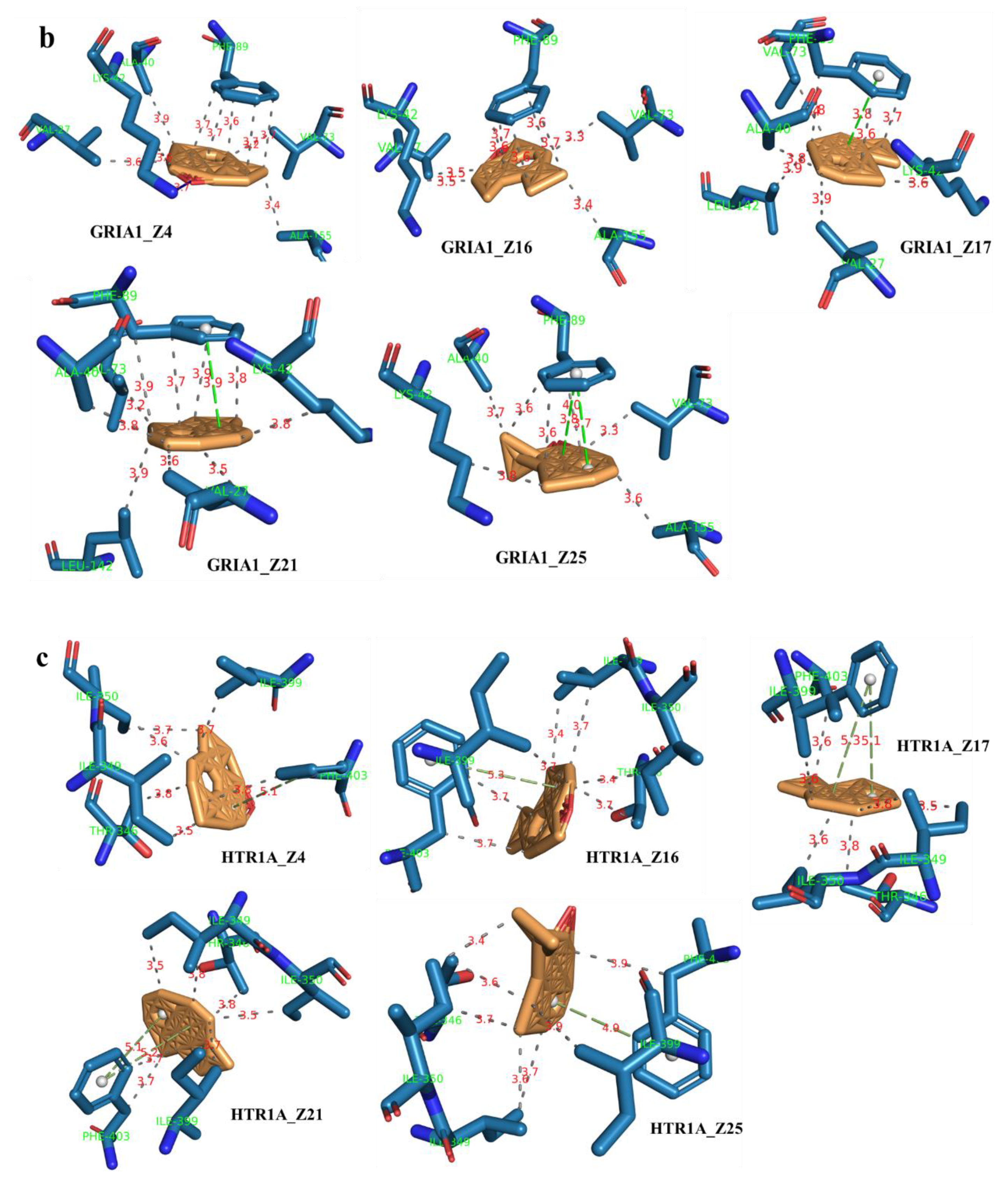
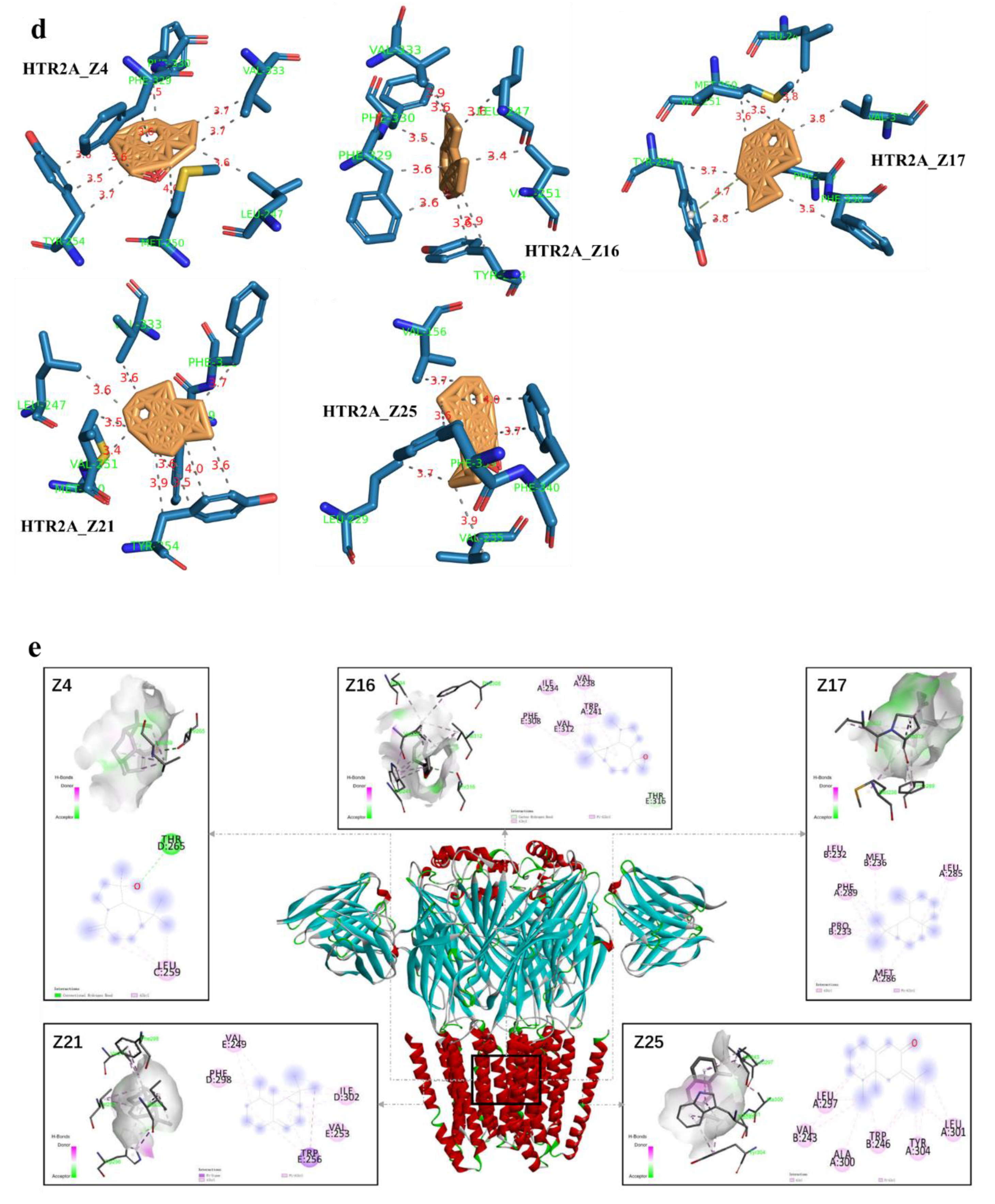
2.7. Component–Target Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Reagents
4.3. Instruments
4.4. Animals
4.5. Animal Experiment and Administration
4.6. Sleep Promotion Test
4.7. Autonomous Activity and Open-Field Detection
4.8. Neurotransmitters Detected by Enzyme-Linked Immunoassay in Brain Tissues
4.9. Protein Expression Detected in Brain Tissue by Western Blotting
4.10. Headspace Solid-Phase Microextraction
4.11. Gas Chromatography–Mass Spectrometry Analysis
4.12. Component–Target Molecular Docking
4.13. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Riemann, D. Sleep hygiene, insomnia and mental health. J. Sleep Res. 2018, 27, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glidewell, R.N.; Mcpherson, B.E.; Orr, W.C. Insomnia and anxiety: Diagnostic and management implications of complex interactions. Sleep Med. Clin. 2015, 10, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Zambotti, M.D.; Goldstone, A.; Colrain, I.M.; Baker, F.C. Insomnia disorder in adolescence: Diagnosis, impact, and treatment. Sleep Med. Rev. 2017, 39, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, G.; Contaldi, P.; Fogliame, G. Evaluation of effectiveness and safety of a herbal 379 compound in primary insomnia symptoms and sleep disturbances not related to medical or psychiatric 380 causes. Nat. Sci. Sleep 2017, 9, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarris, J.; McIntyre, E. Herbal anxiolytics with sedative actions. In Evidence-Based Herbal 384 and Nutritional Treatments for Anxiety in Psychiatric Disorders; Springer: Cham, Switzerland, 2017; pp. 11–31. [Google Scholar]
- Liu, Y.Y.; Wei, J.H.; Gao, Z.H.; Zhang, Z.; Lyu, J.C. A review of quality assessment and grading for agarwood. Chin. Herb. Med. 2017, 9, 22–30. [Google Scholar] [CrossRef]
- Rothan, H.A.; Paydar, M.; Mohamad, J.; Bahrani, H. Isolation and characterisation of acetylcholinesterase inhibitors from aquilaria subintegra for the treatment of Alzheimer’s disease (AD). Curr. Alzheimer Res. 2014, 11, 206–214. [Google Scholar]
- Takemoto, H.; Ito, M.; Shiraki, T.; Yagura, T.; Honda, G. Sedative effects of vapor inhalation of agarwood oil and spikenard extract and identification of their active components. J. Nat. Med. 2007, 62, 41–46. [Google Scholar] [CrossRef]
- Wang, S.; Wang, C.; Peng, D.; Liu, X.; Wu, C.; Guo, P.; Wei, J.H. Agarwood essential oil displays sedative-hypnotic effects through the GABAergic system. Molecules 2017, 22, 2190. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Wang, W.; Zhang, J. Effects of novel anxiolytic 4-butyl-alpha-agarofuran on levels of monoamine neurotransmitters in rats. Eur. J. Pharm. 2004, 504, 39–44. [Google Scholar]
- Huo, H.X.; Zhu, Z.X.; Pang, D.R.; Li, Y.T.; Huang, Z.; Shi, S.P.; Zheng Jiao Zhang, Q.; Zhao, Y.F.; Tu, P.F. Anti-neuroinflammatory sesquiterpenes from Chinese eaglewood. Fitoterapia 2015, 106, 115–121. [Google Scholar] [CrossRef]
- Lin, Y.; Guo, S.X. Two new sesquiterpene derivatives from Chinese eaglewood. J. Asian Nat. Prod. Res. 2012, 14, 1054–1058. [Google Scholar]
- López-Sampson, A.; Page, T. History of use and trade of agarwood. Econ. Bot. 2018, 72, 107–129. [Google Scholar] [CrossRef]
- Liang, Y.; Kong, D.W.; Zhou, Q.M. Effect of Agarwood Gas on the Sleep of Mice by Rgulating Neurotransmitters. Pharm. Clin. TMC 2019, 35, 71–77. [Google Scholar]
- Tanaka, J.; Uchimura, N.; Hashizume, Y.; Shirakawa, S.; Maeda, H. Effects of aroma on sleep and biological rhythms. Psychiatry Clin. Neurosci. 2010, 56, 299–300. [Google Scholar] [CrossRef]
- Lei, L.; Zhang, T.; Gao, D.; Huang, Q.L. Study on clinical effect of agarwood fumigation therapy on patients with insomnia disorder. J. Apoplexy Nerv. Dis. 2019, 36, 609–612. [Google Scholar]
- Lee, R.S.; Steffensen, S.C.; Henriksen, S.J. Discharge profiles of ventral tegmental area gaba neurons during movement, anesthesia, and the sleep-wake cycle. J. Neurosci. 2001, 21, 1757–1766. [Google Scholar] [CrossRef]
- Rao, M. Effects of the contents of gaba and glu in brain of rats of wuweiziningshen oral liquid. Proc. Clin. Med. 2008, 1, 739–740. [Google Scholar]
- Ronald, S.; Janice, D.; Dennis, M.G. Exposure to heat restores sleep in cats with preoptic/anterior hypothalamic cell loss. Brain Res. 1991, 541, 134–138. [Google Scholar]
- Teng, J.; Zhou, W.; Zeng, Z.; Zhao, W.F.; Huang, Y.H.; Zhang, X. Quality components and antidepressant-like effects of GABA green tea. Food Funct. 2017, 8, 3311–3318. [Google Scholar] [CrossRef]
- Yuan, C.S.; Attele, A.S.; Wu, J.A.; Liu, D. Modulation of American ginseng on brainstem GABAergic effects in rats. J. Ethnopharmacol. 1998, 62, 215–222. [Google Scholar] [CrossRef]
- Azuma, S.; Kodama, T.; Honda, K.; Inoué, S. State-dependent changes of extracellular glutamate in the medial preoptic area in freely behaving rats. Neurosci. Lett. 1996, 214, 179–182. [Google Scholar] [CrossRef]
- Ryoo, N.; Rahman, M.A.H.; Wang, H.; Ko, S.K.; Nah, S.Y.; Kim, H.C.; Rhim, H. Ginsenoside Rk1 is a novel inhibitor of NMDA receptors in cultured rat hippocampal neurons. J. Ginseng Res. 2020, 44, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Sattler, R.; Tymianski, M. Molecular mechanisms of glutamate receptor-mediated excitotoxic neuronal cell death. Mol. Neurobiol. 2001, 24, 107–129. [Google Scholar] [CrossRef]
- Li, N.; Liu, J.; Wang, M.; Yu, Z.; Zhu, K.; Gao, J.; Wang, C.; Sun, J.; Chen, J.; Li, H. Sedative and hypnotic effects of Schisandrin B through increasing GABA/Glu ratio and upregulating the expression of GABAA in mice and rats. Biomed. Pharmacother. 2018, 103, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Matito, S.; Cubero, J.; Paredes, S.; Franco, L.; Rivero, M.; Rodríguez, A.; Barriga, C. Tryptophan-enriched cereal intake improves nocturnal sleep, melatonin, serotonin, and total antioxidant capacity levels and mood in elderly humans. Age 2013, 35, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Bjorvatn, B.; Ursin, R. Changes in sleep and wakefulness following 5-HT1A ligands given systemically and locally in different brain regions. Rev. Neurosci. 1998, 9, 265–274. [Google Scholar] [CrossRef]
- Azzi-Achkouty, S.; Estephan, N.; Ouaini, N.; Rutledge, D.N. Headspace solid-phase microextraction for wine volatile analysis. Crit. Rev. Food Sci. Nutr. 2017, 57, 2009–2020. [Google Scholar]
- Tufariello, M.; Pati, S.; D’Amico, L.; Bleve, G.; Losito, I.; Grieco, F. Quantitative issues related to the headspace-SPME-GC/MS analysis of volatile compounds in wines: The case of Maresco sparkling wine. LWT 2019, 108, 268–276. [Google Scholar] [CrossRef]
- Saitoa, K.; Satomi, K.; Yuka, F.; Yumi, A.; Rie, I.; Ken-ichi, S.; Mamoru, A.; Youichi, Y. Confirmation of synthetic cannabinoids in herb and blood by hs-spme-gc/ms. Forensic Chem. 2019, 13, 100156. [Google Scholar] [CrossRef]
- Kataoka, H.; Lord, H.L.; Pawliszyn, J. Applications of solid-phase microextraction in food analysis. J. Chromatogr. A 2000, 880, 35–62. [Google Scholar] [CrossRef]
- Li, W.; Chen, H.Q.; Wang, H.; Mei, W.L.; Dai, H.F. Natural products in agarwood and Aquilaria plants: Chemistry, biological activities and biosynthesis. Nat. Prod. Rep. 2021, 38, 528–565. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Han Xm Sun, Y.; Chen Hj Yang, Y.; Liu, Y.; Meng, H.; Gao, Z.; Xu, Y.; Zhang, Z.; Han, J. Overview of sesquiterpenes and chromones of agarwood originating from four main species of the genus Aquilaria. RSC Adv. 2019, 9, 4113–4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garlet, Q.I.; Pires, L.; Milanesi, L.H.; Marafiga, J.R.; Baldisserotto, B.; Mello CFHeinzmann, B.M. (+)-dehydrofukinone modulates membrane potential and delays seizure onset by gabaa receptor-mediated mechanism in mice. Toxicol. Appl. Pharmacol. 2017, 332, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.Q.; Yang, Y.; Zhang, Z.; Wei, J.H.; Meng, H. Whole-tree agarwood-inducing technique: An efficient novel technique for producing high-quality agarwood in cultivated Aquilaria sinensis trees. Molecules 2013, 18, 3086–3106. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Kim, M.; Brown, C.E.; Wang, Z.; Yang, Z.; Hollebone, B.P.; Lambert, P. Oil fingerprinting analysis using gas chromatography-quadrupole time-of-flight (GC–QTOF). In Standard Handbook Oil Spill Environmental Forensics; Academic Press: Amsterdam, The Netherlands, 2016; pp. 449–480. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunghwan, K.; Thiessen, P.A.; EEBolton Jie, C.; Fu, G.; Asta GHan, L.; He, J.; SHe, S.; Shoemaker, B.A. Pubchem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar]
- Karuppasamy, M.P.; Venkateswaran, S.; Subbiah, P. PDB-2-PBv3.0: An updated protein block database. J. Bioinform. Comput. Biol. 2020, 18, 2050009. [Google Scholar] [CrossRef]
- Sebastian, S.; Sven, S.; Joachim, H.V.; Adasme, M.F.; Michael, S. Plip: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | RT/Min | Sesquiterpene Type | Compound | Peak Area of Intraperitoneal Injection Group/% | Peak Area of Inhalation Group/% | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 min | 30 min | 1 h | 2 h | 10 min | 30 min | 1 h | 2 h | ||||
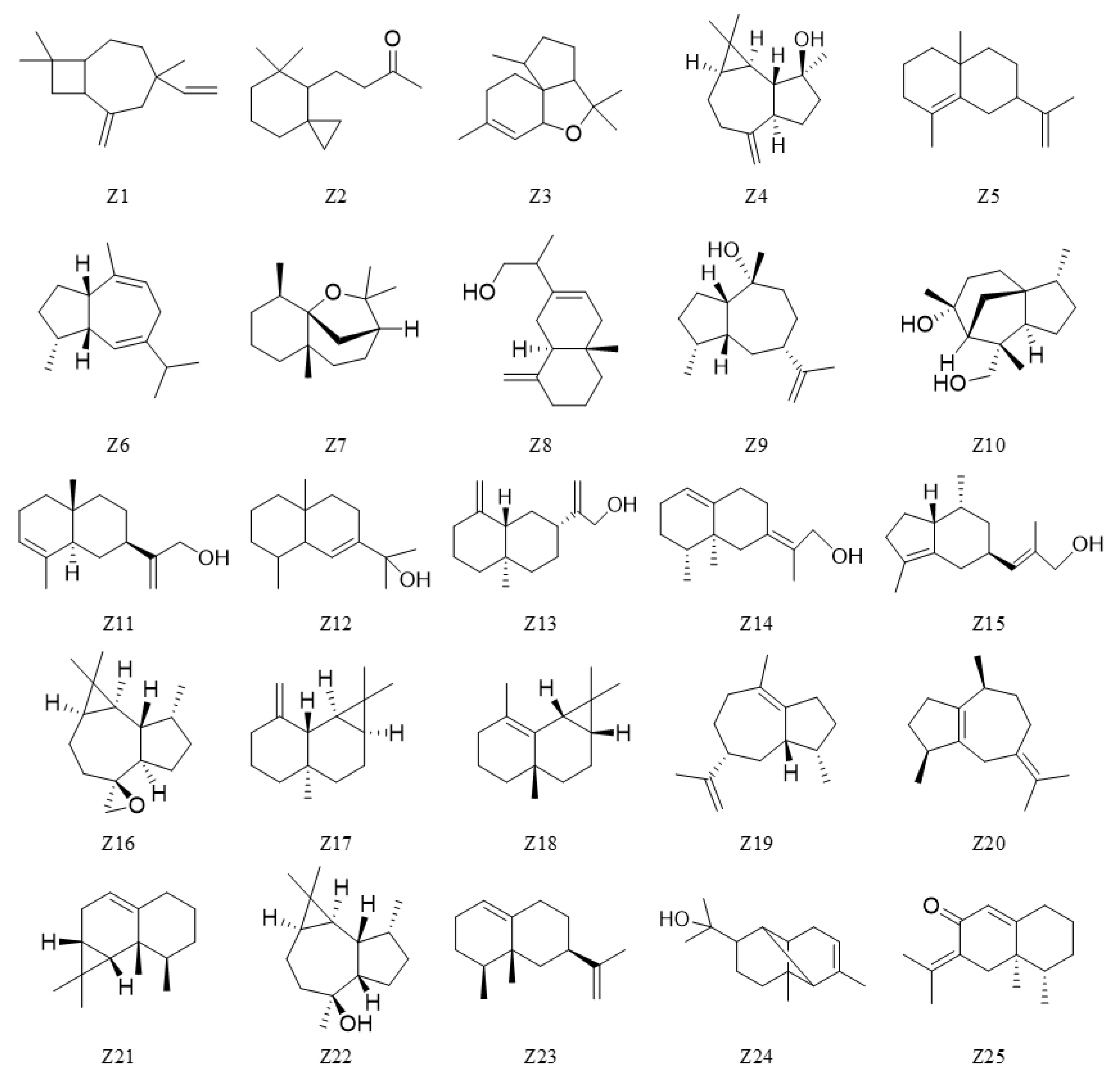
| Z1 | 24.8 | - | Bicyclo [5.2.0]nonane,2-methylene-4,8,8-trimethyl-4-vinyl- | 0.31 | 0.34 | - | - | - | - | - | - |
| Z2 | 25.6 | - | Spiro [2.5]octane,5,5-dimethyl-4-(3-oxobutyl)- | 1.30 | 0.61 | 0.10 | - | - | - | - | - |
| Z3 | 28.3 | Silphiperfolane | (1R,3aR,5aR,9aS)-1,4,4,7-Tetramethyl-1,2,3,3a,4,5a18,9-octahydrocyclopenta[c]benzofuran | 1.24 | 0.86 | 0.26 | - | - | - | - | - |
| Z4 | 30.1 | Aromadendrane | Spathulenol | 8.99 | 5.69 | 0.34 | 0.83 | 0.12 | 0.17 | - | - |
| Z5 | 30.5 | Eremophilane | 4a,8-Dimethyl-2-(prop-1-en-2-yl)-1,2,3,4,4a,5,6,7-octahydronaphthalene | 0.78 | 0.41 | 0.54 | 0.62 | 1.01 | 0.61 | 0.31 | 0.72 |
| Z6 | 31.3 | Guaiane | (1R,3aS,8aS)-7-Isopropyl-1,4-dimethyl-1,2,3,3a,6,8a-hexahydroazulene | 0.38 | 0.47 | - | - | - | - | - | - |
| Z7 | 32.3 | - | Dihydro-beta-agarofuran | 2.84 | 1.16 | - | - | - | - | - | - |
| Z8 | 33.2 | Eremophilane | (R)-2-((4aS,8aR)-4a-Methylene-1,4,4a,5,6,7,8,8a-octahydronaphthalen-2-yl) propan-1-ol | 0.90 | 0.67 | 0.40 | 0.49 | 0.47 | 0.21 | 0.36 | 0.45 |
| Z9 | 35.1 | Guaiane | Pogostol | 0.98 | 0.52 | - | - | - | - | - | - |
| Z10 | 35.3 | Cedrane | α-Costol | 0.16 | 0.11 | - | - | - | - | - | - |
| Z11 | 36.9 | Eremophilane | 2-((2R,4aR,8aR)-4a,8-Dimethyl-1,2,3,4,4a,5,6,8a-octahydronaphthalen-2-yl)prop-2-en-1-ol | 2.45 | 1.14 | - | - | - | 0.26 | - | - |
| Z12 | 38.7 | Eremophilane | 2-((4aS,8R,8aR)-4a,8-Dimethyl)-3,4,4a,5,6,7,8,8a-octahydronaphthalen-2-yl)propan-2-ol | 3.02 | 3.82 | 0.53 | 0.32 | 0.69 | 1.01 | 0.29 | 0.28 |
| Z13 | 40.9 | Eremophilane | (+)-β-Costol | 0.61 | 0.64 | - | - | 0.20 | 0.25 | - | - |
| Z14 | 41.1 | Cadinane | (E)-2-((8R,8aS)-8,8a-Dimethyl-3,4,6,7,8,8a-hexahydronaphthalen-2(1H)-ylidene)propan-1-ol | 0.11 | 0.14 | - | - | - | - | - | - |
| Z15 | 41.5 | Brasilane | Aristol-1(10)-en-9-ol | 0.49 | 0.35 | - | - | - | - | - | - |
| Z16 | 43.5 | Aromadendrane | Aromadendrene oxide-(2) | 19.01 | 16.21 | 14.37 | 5.70 | 2.70 | 2.07 | 2.58 | 0.15 |
| Z17 | 44.6 | Maaliane | γ-Maaliene | 0.82 | 0.49 | - | - | 0.19 | 0.30 | - | - |
| Z18 | 46.2 | Maaliane | β-Maaliene | 0.31 | 0.42 | - | - | - | - | - | - |
| Z19 | 46.6 | Guaiane | Δ-Guaiene | 0.44 | 0.45 | - | - | - | - | - | - |
| Z20 | 46.8 | Guaiane | β-Guaiene | 0.63 | 0.48 | - | - | 0.14 | 0.15 | - | - |
| Z21 | 47.3 | Aristolane | (-)-Aristolene | 1.25 | 1.28 | - | - | 0.31 | 0.47 | - | - |
| Z22 | 47.8 | Aromadendrane | Viridiflorol | 2.85 | 3.27 | 0.42 | - | 0.61 | 0.75 | - | - |
| Z23 | 48.3 | Cadinane | Eremophilene | 3.45 | 4.29 | 0.58 | - | 1.02 | 1.42 | - | - |
| Z24 | 50.4 | - | α-Copaen-11-ol | 0.36 | 3.90 | 0.63 | - | 0.86 | 0.78 | - | - |
| Z25 | 63.2 | Cadinane | Dehydrofukinone | 1.05 | 2.20 | 0.93 | - | 1.17 | 0.89 | - | - |
| Relative percentage of total peak area/% | 55.45 | 49.92 | 19.10 | 7.96 | 9.76 | 6.53 | 3.54 | 1.60 | |||
| Number of total peaks | 25 | 25 | 11 | 5 | 13 | 12 | 4 | 4 | |||
| Ligands | Compound Names | PubChem_CID | Receptors | Affinity (kcal/mol) |
|---|---|---|---|---|
| Z16 | Aromadendrene oxide 2 | 16211192 | GABRA1 | −11.3 |
| Z17 | gamma-Maaliene | 21775138 | GABRA1 | −11.5 |
| Z21 | Aristoler | 530421 | GABRA1 | −10 |
| Z25 | Dehydrofukinone | 177072 | GABRA1 | −10.8 |
| Z4 | Spathulenol | 92231 | GABRA1 | −9.6 |
| Z16 | Aromadendrene oxide 2 | 16211192 | GRIA1 | −10.7 |
| Z17 | gamma-Maaliene | 21775138 | GRIA1 | −10.6 |
| Z21 | Aristoler | 530421 | GRIA1 | −10.7 |
| Z25 | Dehydrofukinone | 177072 | GRIA1 | −10 |
| Z4 | Spathulenol | 92231 | GAIA1 | −10.2 |
| Z16 | Aromadendrene oxide 2 | 16211192 | HTR1A | −9.5 |
| Z17 | gamma-Maaliene | 21775138 | HTR1A | −9.5 |
| Z21 | Aristoler | 530421 | HTR1A | −9.5 |
| Z25 | Dehydrofukinone | 177072 | HTR1A | −9.2 |
| Z4 | Spathulenol | 92231 | HTR1A | −9.2 |
| Z16 | Aromadendrene oxide 2 | 16211192 | HTR2A | −10.6 |
| Z17 | gamma-Maaliene | 21775138 | HTR2A | −10.3 |
| Z21 | Aristolene | 530421 | HTR2A | −10.3 |
| Z25 | Dehydrofukinone | 177072 | HTR2A | −9.6 |
| Z4 | Spathulenol | 92231 | HTR2A | −10.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Wang, Y.; Gong, B.; Wu, Y.; Chen, X.; Liu, Y.; Wei, J. Effective Components and Molecular Mechanism of Agarwood Essential Oil Inhalation and the Sedative and Hypnotic Effects Based on GC-MS-Qtof and Molecular Docking. Molecules 2022, 27, 3483. https://doi.org/10.3390/molecules27113483
Wang C, Wang Y, Gong B, Wu Y, Chen X, Liu Y, Wei J. Effective Components and Molecular Mechanism of Agarwood Essential Oil Inhalation and the Sedative and Hypnotic Effects Based on GC-MS-Qtof and Molecular Docking. Molecules. 2022; 27(11):3483. https://doi.org/10.3390/molecules27113483
Chicago/Turabian StyleWang, Canhong, Yunyun Wang, Bao Gong, Yulan Wu, Xiqin Chen, Yangyang Liu, and Jianhe Wei. 2022. "Effective Components and Molecular Mechanism of Agarwood Essential Oil Inhalation and the Sedative and Hypnotic Effects Based on GC-MS-Qtof and Molecular Docking" Molecules 27, no. 11: 3483. https://doi.org/10.3390/molecules27113483
APA StyleWang, C., Wang, Y., Gong, B., Wu, Y., Chen, X., Liu, Y., & Wei, J. (2022). Effective Components and Molecular Mechanism of Agarwood Essential Oil Inhalation and the Sedative and Hypnotic Effects Based on GC-MS-Qtof and Molecular Docking. Molecules, 27(11), 3483. https://doi.org/10.3390/molecules27113483