
Quercetin-Rich Ethanolic Extract of Polygonum odoratum var Pakphai Leaves Decreased Gene Expression and Secretion of Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Murine RAW264.7 Macrophages


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Extraction Yield, Total Phenolic and Flavonoid Contents
2.2. UHPLC-ESI-QTOF-MS/MS Profiling of the Extract
2.3. HPLC-DAD Analysis of Phenolic Compounds in the Extract
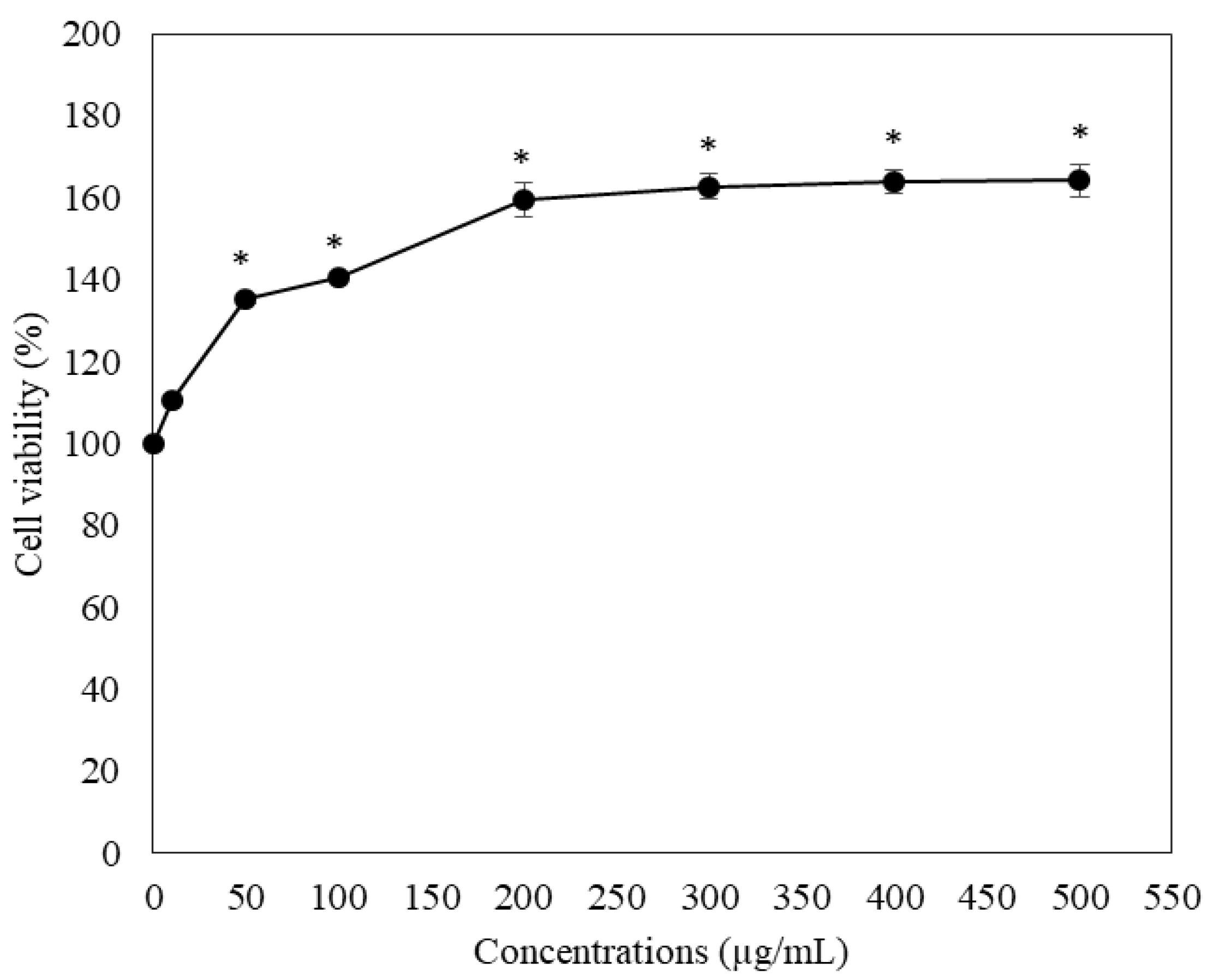
2.4. Toxic Effect on RAW264.7 Macrophage Cells
2.5. Effect on Nitric Oxide Production and Inducible Nitric Oxide Synthase Gene Expression
2.6. Inhibitory Effects on Cyclooxygenase 2 Gene Expression and PGE2 Production
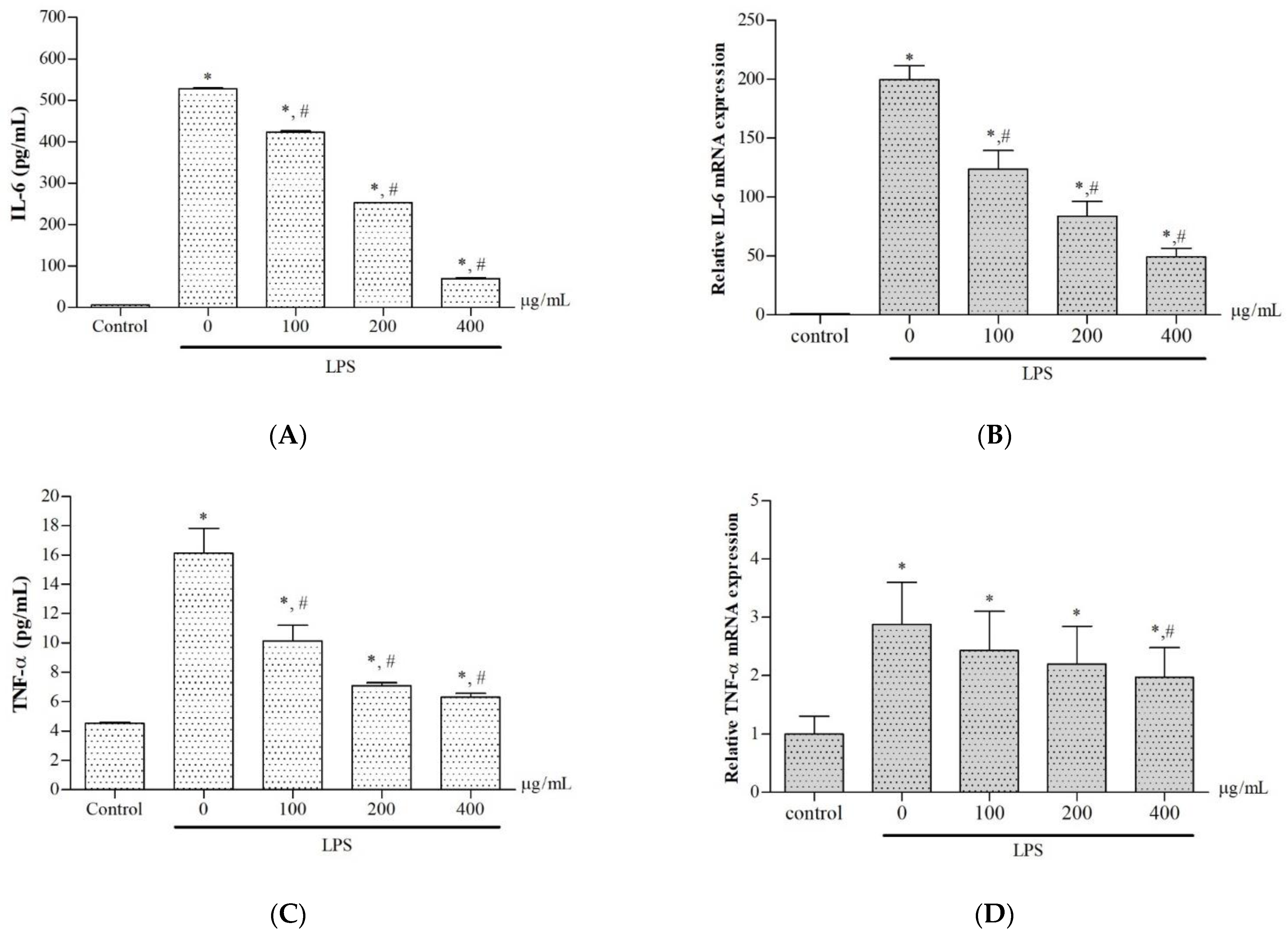
2.7. Effects on Production and Gene Expression of Pro-Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Preparation of P. odoratum Ethanolic Extract
4.4. Determination of TPC
4.5. Determination of TFC
4.6. UHPLC-ESI-QTOF-MS/MS Analysis
4.7. HPLC-DAD Analysis
4.8. Cell Viability Assay
4.9. Assessment of Pro-Inflammatory Mediators
4.10. Determination of Nitric Oxide Concentrations
4.11. Determination of PGE2, IL-6 and TNFα Concentrations
4.12. Quantitative Real-Time Polymerase Chain Reaction
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Neagu, M.; Constantin, C.; Caruntu, C.; Dumitru, C.; Surcel, M.; Zurac, S. Inflammation: A key process in skin tumorigenesis. Oncol. Lett. 2019, 17, 4068–4084. [Google Scholar] [CrossRef] [Green Version]
- Ohkawara, S.; Goto, F.; Yoshinaga, M. Cytokines as inflammatory mediator. Tanpakushitsu Kakusan Koso 1991, 36, 935–943. Available online: https://www.ncbi.nlm.nih.gov/pubmed/1866451 (accessed on 5 May 2021).
- Chae, H.S.; Kang, O.H.; Lee, Y.S.; Choi, J.G.; Oh, Y.C.; Jang, H.J.; Kim, M.S.; Kim, J.H.; Jeong, S.I.; Kwon, D.Y. Inhibition of LPS-induced iNOS, COX-2 and inflammatory mediator expression by paeonol through the MAPKs inactivation in RAW 264.7 cells. Am. J. Chin. Med. 2009, 37, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Gardener, S.L.; Rainey-Smith, S.R.; Martins, R.N. Diet and inflammation in Alzheimer’s disease and related chronic diseases: A Review. J. Alzheimers Dis. 2016, 50, 301–334. [Google Scholar] [CrossRef]
- Howcroft, T.K.; Campisi, J.; Louis, G.B.; Smith, M.T.; Wise, B.; Wyss-Coray, T.; Augustine, A.D.; McElhaney, J.E.; Kohanski, R.; Sierra, F. The role of inflammation in age-related disease. Aging 2013, 5, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corlett, J.L.; Clegg, M.S.; Keen, C.L.; Grivetti, L.E. Mineral content of culinary and medicinal plants cultivated by Hmong refugees living in Sacramento, California. Int. J. Food Sci. Nutr. 2002, 53, 117–128. [Google Scholar] [CrossRef]
- Chansiw, N.; Chotinantakul, K.; Srichairatanakool, S. Anti-inflammatory and antioxidant activities of the extracts from leaves and stems of Polygonum odoratum Lour. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2019, 18, 45–54. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Arraouadi, S.; Mighri, H.; Ghannay, S.; Aouadi, K.; Adnan, M.; Elasbali, A.M.; Noumi, E.; Snoussi, M.; Kadri, A. HPLC-MS profiling, antioxidant, antimicrobial, antidiabetic, and cytotoxicity activities of Arthrocnemum indicum (Willd.) Moq. extracts. Plants 2022, 11, 232. [Google Scholar] [CrossRef]
- Abu-Gharbieh, E.; Shehab, N.G. Therapeutic potentials of Crataegus azarolus var. eu-azarolus Maire leaves and its isolated compounds. BMC Complement. Altern. Med. 2017, 17, 218. [Google Scholar] [CrossRef] [PubMed]
- Baptista, A.B.; Sarandy, M.M.; Goncalves, R.V.; Novaes, R.D.; Goncalves da Costa, C.; Leite, J.P.V.; Peluzio, M. Antioxidant and anti-inflammatory effects of Anacardium occidentale L. and Anacardium microcarpum D. extracts on the liver of IL-10 knockout mice. Evid. Based Complement. Altern. Med. 2020, 2020, 3054521. [Google Scholar] [CrossRef]
- Caprioli, G.; Iannarelli, R.; Innocenti, M.; Bellumori, M.; Fiorini, D.; Sagratini, G.; Vittori, S.; Buccioni, M.; Santinelli, C.; Bramucci, M.; et al. Blue honeysuckle fruit (Lonicera caerulea L.) from eastern Russia: Phenolic composition, nutritional value and biological activities of its polar extracts. Food Funct. 2016, 7, 1892–1903. [Google Scholar] [CrossRef]
- El-Hawary, S.A.; Sokkar, N.M.; Ali, Z.Y.; Yehia, M.M. A profile of bioactive compounds of Rumex vesicarius L. J. Food Sci. 2011, 76, C1195–C1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ye, Y.H.; Wang, H.H.; Liu, J.; Liu, Y.J.; Jiang, B.W. HPLC-QTOF-MS/MS profiling, antioxidant, and alpha-glucosidase inhibitory activities of Pyracantha fortuneana fruit extracts. J. Food Biochem. 2019, 43, e12821. [Google Scholar] [CrossRef]
- Liu, X.; Yan, X.; Bi, J.; Liu, J.; Zhou, M.; Wu, X.; Chen, Q. Determination of phenolic compounds and antioxidant activities from peel, flesh, seed of guava (Psidium guajava L.). Electrophoresis 2018, 39, 1654–1662. [Google Scholar] [CrossRef] [PubMed]
- Okonogi, S.; Kheawfu, K.; Holzer, W.; Unger, F.M.; Viernstein, H.; Mueller, M. Anti-inflammatory effects of ocmpounds from Polygonum odoratum. Nat. Prod. Commun. 2016, 11, 1651–1654. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30475499 (accessed on 5 May 2021).
- Murray, A.F.; Satooka, H.; Shimizu, K.; Chavasiri, W.; Kubo, I. Polygonum odoratum essential oil inhibits the activity of mushroom derived tyrosinase. Heliyon 2019, 5, e02817. [Google Scholar] [CrossRef] [PubMed]
- Ngamsurach, P.; Praipipat, P. Modified alginate beads with ethanol extraction of Cratoxylum formosum and Polygonum odoratum for antibacterial activities. ACS Omega 2021, 6, 32215–32230. [Google Scholar] [CrossRef]
- Starkenmann, C.; Luca, L.; Niclass, Y.; Praz, E.; Roguet, D. Comparison of volatile constituents of Persicaria odorata(Lour.) Sojak (Polygonum odoratum Lour.) and Persicaria hydropiper L. Spach (Polygonum hydropiper L.). J. Agric. Food Chem. 2006, 54, 3067–3071. [Google Scholar] [CrossRef]
- Fujita, K.; Chavasiri, W.; Kubo, I. Anti-Salmonella activity of volatile compounds of Vietnam coriander. Phytother. Res. 2015, 29, 1081–1087. [Google Scholar] [CrossRef]
- Yamazaki, S.; Miyoshi, N.; Kawabata, K.; Yasuda, M.; Shimoi, K. Quercetin-3-O-glucuronide inhibits noradrenaline-promoted invasion of MDA-MB-231 human breast cancer cells by blocking beta(2)-adrenergic signaling. Arch. Biochem. Biophys. 2014, 557, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.H.; Hwangbo, K.; Zheng, M.S.; Cho, J.H.; Son, J.K.; Kim, H.Y.; Baek, S.H.; Choi, H.C.; Park, S.Y.; Kim, J.R. Quercetin-3-O-beta-D-glucuronide isolated from Polygonum aviculare inhibits cellular senescence in human primary cells. Arch. Pharm. Res. 2014, 37, 1219–1233. [Google Scholar] [CrossRef] [PubMed]
- Shirai, M.; Moon, J.H.; Tsushida, T.; Terao, J. Inhibitory effect of a quercetin metabolite, quercetin 3-O-beta-D-glucuronide, on lipid peroxidation in liposomal membranes. J. Agric. Food Chem. 2001, 49, 5602–5608. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.E.; Kim, O.S.; Yoo, S.R.; Seo, C.S.; Kim, Y.; Shin, H.K.; Jeong, S.J. Anti-inflammatory effect and action mechanisms of traditional herbal formula Gamisoyo-san in RAW 264.7 macrophages. BMC Complement. Altern. Med. 2016, 16, 219. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Sun, X.Y.; Li, X.W.; Yang, T.; Qi, L.W. Enrichment and separation of quercetin-3-O-beta-D-glucuronide from lotus leaves (Nelumbo nucifera gaertn.) and evaluation of its anti-inflammatory effect. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1040, 186–191. [Google Scholar] [CrossRef]
- Fan, D.; Zhou, X.; Zhao, C.; Chen, H.; Zhao, Y.; Gong, X. Anti-inflammatory, antiviral and quantitative study of quercetin-3-O-beta-D-glucuronide in Polygonum perfoliatum L. Fitoterapia 2011, 82, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.B.; Xiao, T.; Gong, G.Y.; Bian, Y.Q.; Li, Y.F. A new isoflavone with anti-inflammatory effect from the seeds of Millettia pachycarpa. Nat. Prod. Res. 2020, 34, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Chung, C.P.; Chiang, W.; Lin, Y.L. Anti-inflammatory effects and chemical study of a flavonoid-enriched fraction from adlay bran. Food Chem. 2011, 126, 1741–1748. [Google Scholar] [CrossRef]
- Sungkamanee, S.; Wattanathorn, J.; Muchimapura, S.; Thukham-mee, W. Antiosteoporotic effect of combined extract of Morus alba and Polygonum odoratum. Oxidative Med. Cell. Longev. 2014, 2014, 579305. [Google Scholar] [CrossRef] [Green Version]
- Wattanathorn, J.; Somboonporn, W.; Sungkamanee, S.; Thukummee, W.; Muchimapura, S. A double-blind placebo-controlled randomized trial evaluating the effect of polyphenol-rich herbal congee on bone turnover markers of the perimenopausal and menopausal women. Oxidative Med. Cell. Longev. 2018, 2018, 2091872. [Google Scholar] [CrossRef] [Green Version]
- Itharat, A.; Hiransai, P. Dioscoreanone suppresses LPS-induced nitric oxide production and inflammatory cytokine expression in RAW 264.7 macrophages by NF-kappaB and ERK1/2 signaling transduction. J. Cell. Biochem. 2012, 113, 3427–3435. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Jeong, D.; Yi, Y.S.; Lee, B.H.; Kim, T.W.; Htwe, K.M.; Kim, Y.D.; Yoon, K.D.; Hong, S.; Lee, W.S.; et al. Myrsine seguinii ethanolic extract and its active component quercetin inhibit macrophage activation and peritonitis induced by LPS by targeting to Syk/Src/IRAK-1. J. Ethnopharmacol. 2014, 151, 1165–1174. [Google Scholar] [CrossRef]
- Cho, B.O.; Nchang Che, D.; Yin, H.H.; Jang, S.I. Enhanced biological activities of gamma-irradiated persimmon leaf extract. J. Radiat. Res. 2017, 58, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.S.; Weng, H.Z.; Huang, J.L.; Tang, G.H.; Yin, S. Anti-inflammatory ingenane diterpenoids from the roots of Euphorbia kansui. Planta. Med. 2018, 84, 1334–1339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, X.; Li, J.; Hu, Y.; Liu, J.; Wang, F.; Zhang, W.; Chang, F. Mechanism of anti-inflammatory and antibacterial effects of QingXiaoWuWei decoction based on network pharmacology, molecular docking and in vitro experiments. Front. Pharmacol. 2021, 12, 678685. [Google Scholar] [CrossRef]
- Kawvised, S.; Prabsattroo, T.; Munkong, W.; Pattum, P.; Iamsaard, S.; Boonsirichai, K.; Uttayarat, P.; Maikaeo, L.; Sudchai, W.; Kirisattayakul, W. Polygonum odoratum leaf extract attenuates oxidative stress and cell death of Raw 264.7 cells exposed to low dose ionizing radiation. J. Food Biochem. 2022, 46, e13909. [Google Scholar] [CrossRef]
- Duangjai, A.; Parseatsook, K.; Sajjapong, W.; Saokaew, S. Assessment of Polygonum odoratum Lour. leaf extract on rat’s ileum contraction and the mechanisms involved. J. Med. Food 2020, 23, 1169–1175. [Google Scholar] [CrossRef]
- Park, J.Y.; Lim, M.S.; Kim, S.I.; Lee, H.J.; Kim, S.S.; Kwon, Y.S.; Chun, W. Quercetin-3-O-beta-D-glucuronide suppresses lipopolysaccharide-induced JNK and ERK phosphorylation in LPS-challenged RAW264.7 cells. Biomol. Ther. 2016, 24, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieroni, L.G.; de Rezende, F.M.; Ximenes, V.F.; Dokkedal, A.L. Antioxidant activity and total phenols from the methanolic extract of Miconia albicans (Sw.) Triana leaves. Molecules 2011, 16, 9439–9450. [Google Scholar] [CrossRef]
- Curcic, M.G.; Stankovic, M.S.; Radojevic, I.D.; Stefanovic, O.D.; Comic, L.R.; Topuzovic, M.D.; Djacic, D.S.; Markovic, S.D. Biological effects, total phenolic content and flavonoid concentrations of fragrant yellow onion (Allium flavum L.). Med. Chem. 2012, 8, 46–51. [Google Scholar] [CrossRef]
- Pintha, K.; Yodkeeree, S.; Pitchakarn, P.; Limtrakul, P. Anti-invasive activity against cancer cells of phytochemicals in red jasmine rice (Oryza sativa L.). Asian Pac. J. Cancer Prev. 2014, 15, 4601–4607. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, M.; Fornasiero, M.C.; Isetta, A.M. MTT colorimetric assay for testing macrophage cytotoxic activity in vitro. J. Immunol. Methods 1990, 131, 165–172. [Google Scholar] [CrossRef]
- Lee, J.W.; Bae, C.J.; Choi, Y.J.; Kim, S.I.; Kim, N.H.; Lee, H.J.; Kim, S.S.; Kwon, Y.S.; Chun, W. 3,4,5-Trihydroxycinnamic acid inhibits LPS-induced iNOS expression by suppressing NF-kappaB activation in BV2 microglial cells. Korean J. Physiol. Pharmacol. 2012, 16, 107–112. [Google Scholar] [CrossRef] [Green Version]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A&B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Kienast, K.; Knorst, M.; Muller-Quernheim, J.; Ferlinz, R. Modulation of IL-1 beta, IL-6, IL-8, TNF-alpha, and TGF-beta secretions by alveolar macrophages under NO2 exposure. Lung 1996, 174, 57–67. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | TR (min) | Mode of Ionization | Estimated Mass (m/z) | Exact Mass [M] (g/mole) | Molecular Formula | Error (ppm) | Major MS/MS Fragments | Compounds | Chemical Structure |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.356 | - | - | - | - | - | - | unidentified | - |
| 2 | 0.677 | - | - | - | - | - | - | unidentified | - |
| 3 | 0.913 | - | - | - | - | - | - | unidentified | - |
| 4 | 2.112 | - | - | - | - | - | - | unidentified | - |
| 5 | 9.455 | [M—H]− | 478.07 | 478.36 | C21H18O13 | 0.41 | 301.5, 162.1 | Quercetin 3′-O-glucuronide |  |
| 6 | 11.016 | [M—H]− | 462.08 | 462.40 | C21H18O12 | 0.21 | 317.0, 147.0 | 3-Methylellagic acid 8-rhamnoside |  |
| 7 | 14.787 | [M—H]− | 302.04 | 302.23 | C15H10O7 | 0.66 | 285.0, 193.0, 109.0 | 5,7,8,3′,4′-Pentahydroxyisoflavone or Q |  |
| 8 | 17.112 | - | - | - | - | - | - | - | - |
| 9 | 17.312 | [M—H]+ | 291.21 | 290.26 | C15H14O6 | 0.02 | 287.0, 269.9 | EC |  |
| 10 | 18.242 | [M—H]− | 305.24 | 306.27 | C15H14O7 | 0.34 | 125.0, 137.0 | GC |  |
| 11 | 20.468 | - | 518.13 | - | - | - | - | unidentified | - |
| 12 | 21.531 | [M—H]− | 609.16 | 610.19 | C28H34O15 | 0.65 | 304.2, 157.1 | Hesperidin |  |
| 13 | 22.146 | - | - | - | - | - | - | - | - |
| 14 | 22.761 | [M—H]− | 121.09 | 121.18 | C8H11N | 3.30 | 109.0 | 2,6-Dimethylaniline |  |
| Peak No. | TR (min) | Compounds | MW (g/mol) | Amount | |
|---|---|---|---|---|---|
| (μg/mL Extract) | (μg/mg Dry Extract) | ||||
| 2 | 7.02 | GA | 170.12 | 3.85 ± 1.17 | 1.93 ± 0.04 |
| 3 | 12.27 | C | 290.26 | 1.70 ± 0.83 | 0.85 ± 0.04 |
| 4 | 14.00 | EGCG | 458.37 | ND | ND |
| 5 | 15.40 | EC | 290.26 | ND | ND |
| 6 | 17.28 | ECG | 442.37 | 10.85 ± 5.15 | 5.43 ± 0.01 |
| 7 | 21.27 | CO | 146.14 | 7.91 ± 0.67 | 3.95 ± 0.08 |
| 8 | 26.48 | Q | 302.24 | 13.47 ± 6.11 | 6.73 ± 0.01 |
| Gene | Forward | Reverse |
|---|---|---|
| COX-2 | TTCCTCTACATAAGCCAGTGA | TCCACATTACATGCTCCTATC |
| iNOS | AAGGTCTACGTTCAGGACATC | AGAAATAGTCTTCCACCTGCT |
| IL-6 | GAGGATACCACTCCCAACAGACC | AAGTGCATCATCGTTGTTCATACA |
| TNFα | CATCTTCTCAAAATTCGAGTGACAA | TGGGAGTAGACAAGGTACAACCC |
| GAPDH | TGTGTCCGTCGTGGATCTGA | CCTGCTTCACCACCTTCTTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chansiw, N.; Champakam, S.; Chusri, P.; Pangjit, K.; Srichairatanakool, S. Quercetin-Rich Ethanolic Extract of Polygonum odoratum var Pakphai Leaves Decreased Gene Expression and Secretion of Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Murine RAW264.7 Macrophages. Molecules 2022, 27, 3657. https://doi.org/10.3390/molecules27123657
Chansiw N, Champakam S, Chusri P, Pangjit K, Srichairatanakool S. Quercetin-Rich Ethanolic Extract of Polygonum odoratum var Pakphai Leaves Decreased Gene Expression and Secretion of Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Murine RAW264.7 Macrophages. Molecules. 2022; 27(12):3657. https://doi.org/10.3390/molecules27123657
Chicago/Turabian StyleChansiw, Nittaya, Sorraya Champakam, Pattranuch Chusri, Kanjana Pangjit, and Somdet Srichairatanakool. 2022. "Quercetin-Rich Ethanolic Extract of Polygonum odoratum var Pakphai Leaves Decreased Gene Expression and Secretion of Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Murine RAW264.7 Macrophages" Molecules 27, no. 12: 3657. https://doi.org/10.3390/molecules27123657
APA StyleChansiw, N., Champakam, S., Chusri, P., Pangjit, K., & Srichairatanakool, S. (2022). Quercetin-Rich Ethanolic Extract of Polygonum odoratum var Pakphai Leaves Decreased Gene Expression and Secretion of Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Murine RAW264.7 Macrophages. Molecules, 27(12), 3657. https://doi.org/10.3390/molecules27123657