
Unveiling Chemical Cues of Insect-Tree and Insect-Insect Interactions for the Eucalyptus Weevil and Its Egg Parasitoid by Multidimensional Gas Chromatographic Methods



Abstract
:
1. Introduction
2. Results
2.1. GC/MS-EAD: Detection and Identification of Bioactive Compounds
2.2. Circadian Variation of G. platensis Emitted Compounds
2.3. Behavioral Bioassays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Spent in Arm (s) | Virgin Females | Mated Females | Virgin Males | Mated Males | ||||
|---|---|---|---|---|---|---|---|---|
| + | − | + | − | + | − | + | − | |
| Compound Blend | ||||||||
| cis + trans-Verbenol | 601 ± 249 | 482 ± 267 | 448 ± 323 | 657 ± 315 | 566 ± 301 | 452 ± 303 | 478 ± 394 | 572 ± 419 |
| cis + trans-Verbenol + verbenene | 631 ± 371 | 455 ± 356 | 489 ± 347 | 573 ± 343 | 635 ± 358 | 442 ± 363 | ||
| cis + trans-Verbenol + verbenene + myrtenol | 595 ± 341 | 488 ± 336 | 500 ± 255 | 619 ± 254 | 603 ± 390 | 457 ± 376 | 485 ± 331 | 543 ± 362 |
| cis + trans-Verbenol + verbenene + myrtenol + trans-pinocarveol | 672 ± 328 * | 435 ± 310 | 550 ± 398 | 511 ± 390 | 653 ± 371 * | 446 ± 358 | 520 ± 281 | 558 ± 288 |
3. Discussion
3.1. The Co-Elution Problem
3.2. GC×GC-FID
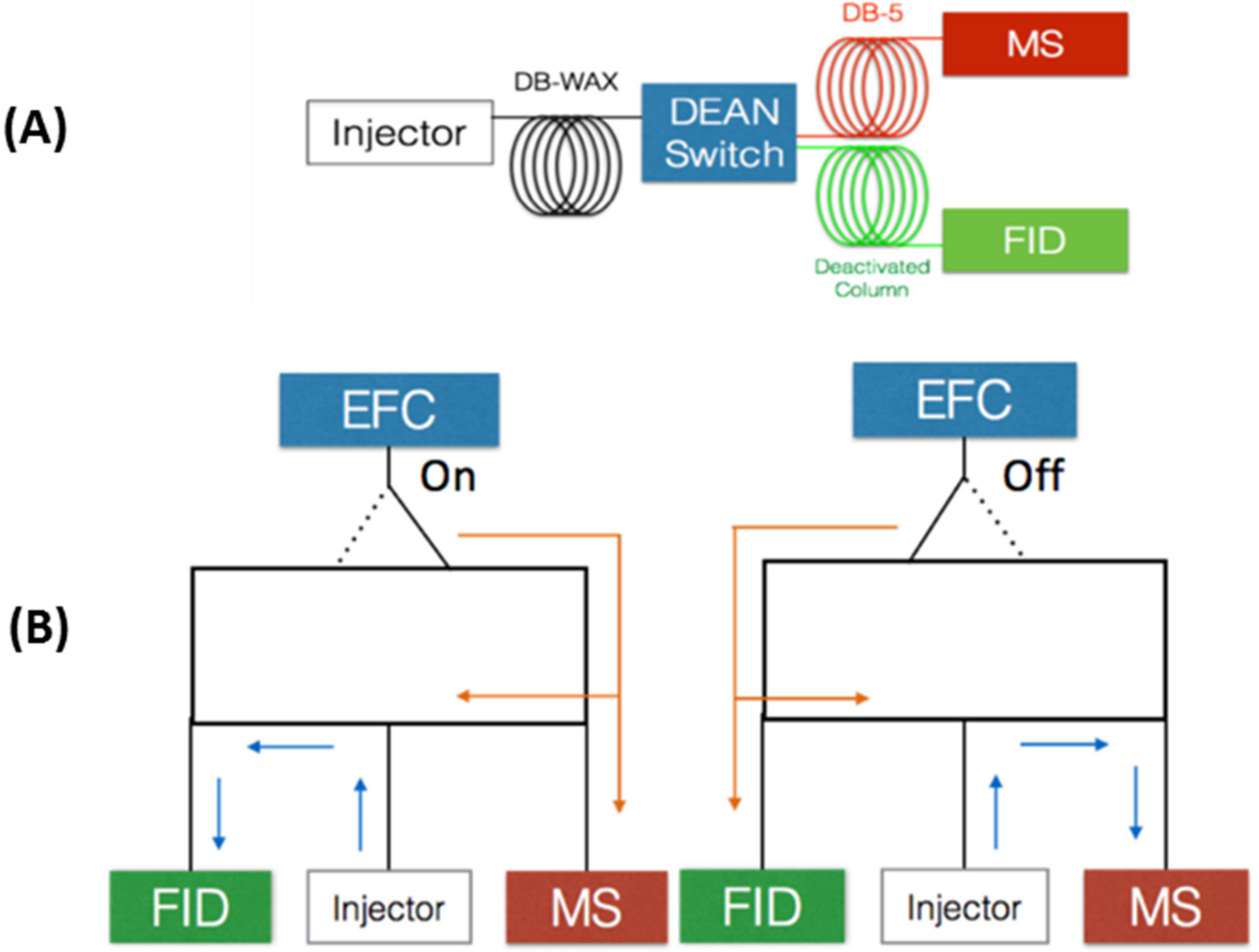
3.3. MD-GC with Dean Switch—Heart-Cutting
4. Materials and Methods
4.1. Reagents and Materials
4.2. Insect Rearing and Plant Sampling
4.3. Collection of VOCs
4.3.1. Headspace Monolithic Material Sorption Extraction (HS-MMSE)-MonoTrap™ Disks
4.3.2. Simultaneous Distillation–Extraction (SDE)
4.3.3. Extraction with Dichloromethane
4.3.4. Headspace Solid-Phase Microextraction (HS-SPME)
4.4. Gas Chromatography/Mass Spectrometry (GC/MS)
4.5. MD-GC/MS Analysis
4.6. GC×GC-FID Analysis
4.7. Gas Chromatography–Mass Spectrometry/Electroantennographic Detection (GC-MS/EAD) Analysis
4.8. Behavioral Bioassays
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
List of Abbreviations and Acronyms
| 1D | One-dimensional |
| 2D | Two-dimensional |
| 1D | First dimension column |
| 2D | Second dimension column |
| df | Film thickness |
| EAD | Electroantennographic detection |
| EAG | Electroantennography |
| EFC | Electronic flow control |
| EI | Electron ionization |
| CF | Constant flow |
| CP | Constant pressure |
| DCM | Dichloromethane |
| DS | Dean switch |
| FID | Flame ionization detector |
| GC | Gas chromatography |
| GC×GC | Comprehensive two-dimensional gas chromatography |
| GC×GC-FID | Comprehensive two-dimensional gas chromatography -flame ionization detection |
| GC/MS | Gas chromatography/mass spectrometry |
| GC/MS-EAD | Gas chromatography/mass spectrometry-electroantennographic detection |
| H/C | Heart-cut |
| H/C-MD-GC | Heart-cut multidimensional gas chromatography |
| HS | Headspace |
| HS-MMSE | Headspace monolithic material sorption extraction |
| HS-SPME | Headspace solid phase microextraction |
| i.d. | Internal diameter |
| IPM | Integrated pest management |
| LRI | Linear retention indices |
| MD-GC | Multidimensional gas chromatography |
| MD-GC/MS | Multidimensional gas chromatography/mass spectrometry |
| MS | Mass spectrometry |
| MSD | Mass selective detector |
| nc | Peak capacity |
| PM | Modulation period |
| SDE | Simultaneous distillation-extraction |
| VOCs | Volatile organic compounds |
References
- Mapondera, T.S.; Burgess, T.; Matsuki, M.; Oberprieler, R.G. Identification and molecular phylogenetics of the cryptic species of the Gonipterus scutellatus complex (Coleoptera: Curculionidae: Gonipterini). Aust. J. Entomol. 2012, 51, 175–188. [Google Scholar] [CrossRef]
- Reis, A.R.; Ferreira, L.; Tomé, M.; Araujo, C.; Branco, M. Efficiency of biological control of Gonipterus platensis (Coleoptera: Curculionidae) by Anaphes nitens (Hymenoptera: Mymaridae) in cold areas of the Iberian Peninsula: Implications for defoliation and wood production. For. Ecol. Manag. 2012, 270, 216–222. [Google Scholar] [CrossRef]
- Gumovsky, A.V.; De Little, D.; Rothmann, S.; Jaques, L.; Ide, S.E. Re-description and first host and biology records of Entedon magnificus (Girault & Dodd) (Hymenoptera, Eulophidae), a natural enemy of Gonipterus weevils (Coleoptera, Curculionidae), a pest of Eucalyptus trees. Zootaxa 2015, 3957, 577–584. [Google Scholar] [PubMed]
- Schröder, M.L.; Nahrung, H.F.; de Souza, N.M.; Lawson, S.A.; Slippers, B.; Wingfield, M.J.; Hurley, B.P. Distribution of Gonipterus species and their egg parasitoids in Australia: Implications for biological control. Forests 2021, 12, 969. [Google Scholar] [CrossRef]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- El-Shafie, H.A.F.; Faleiro, J.R. Semiochemicals and Their Potential Use in Pest Management. In Biological Control of Pest and Vector Insects; IntechOpenScience: London, UK, 2017. [Google Scholar]
- Branco, S.; Mateus, E.; Schütz, S.; Paiva, M.R. The eucalyptus weevil Gonipterus platensis (Coleoptera, Curculionidae): New control perspectives based on semiochemicals. IOBC-WPRS Bull. 2019, 146, 32–34. [Google Scholar]
- Rizvi, S.A.H.; George, J.; Reddy, G.V.; Zeng, X.; Guerrero, A. Latest developments in insect sex pheromone research and its application in agricultural pest management. Insects 2021, 12, 484. [Google Scholar] [CrossRef]
- Branco, S.; Mateus, E.P.; Gomes da Silva, M.D.R.; Mendes, D.; Pereira, M.M.A.; Schütz, S.; Paiva, M.R. Identification of pheromone candidates for the eucalyptus weevil, Gonipterus platensis (Coleoptera, Curculionidae). J. Appl. Entomol. 2020, 144, 41–53. [Google Scholar] [CrossRef]
- Branco, S.; Mateus, E.P.; Gomes da Silva, M.D.; Mendes, D.; Rocha, S.; Mendel, Z.; Schütz, S.; Paiva, M.R. Electrophysiological and behavioural responses of the Eucalyptus weevil, Gonipterus platensis, to host plant volatiles. J. Pest. Sci. 2019, 92, 221–235. [Google Scholar] [CrossRef]
- Branco, S.; Mateus, E.P.; Gomes da Silva, M.D.R.; Mendes, D.; Pereira, M.M.A.; Schütz, S.; Paiva, M.R. Olfactory responses of Anaphes nitens (Hymenoptera, Mymaridae) to host and habitat cues. J. Appl. Entomol. 2021, 145, 675–687. [Google Scholar] [CrossRef]
- Mateus, E.; Farral, M.H.; Paiva, M.R. Characterization of the physiological condition of Eucalyptus globulus Labill by headspace HRGC analysis of the bouquet of odours. J. Microcol. Sep. 1995, 7, 641–645. [Google Scholar] [CrossRef]
- Silvestre, A.J.D.; Cavaleiro, J.A.S.; Delmond, B.; Filliarte, C.; Bourgeois, G. The essential oil of Eucalyptus globulus Labill. from Portugal. Flav. Frag. J. 1994, 9, 51–53. [Google Scholar] [CrossRef]
- Moore, B.D.; Wallis, I.R.; Wood, J.T.; Foley, W.J. Foliar nutrition, site quality, and temperature influence foliar chemistry of tallowwood (Eucalyptus microcorys). Ecol. Mon. 2004, 74, 553–568. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Visser, J.H. Host odor perception in phytophagous insects. Annu. Rev. Entomol. 1986, 31, 121–144. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; Von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “talking trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B. Twenty-five years of chemical ecology. Nat. Prod. Rep. 2001, 18, 361. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Barreca, S.; La Bella, S.; Maggio, A.; Licata, M.; Buscemi, S.; Leto, C.; Pace, A.; Tuttolomondo, T. Flavouring extra-virgin olive oil with aromatic and medicinal plants essential oils stabilizes oleic acid composition during photo-oxidative stress. Agriculture 2021, 11, 266. [Google Scholar] [CrossRef]
- Gomes da Silva, M.D.R.; Mateus, E.P.; Munhá, J.; Drazyk, A.; Farrall, M.H.; Paiva, M.R.; Chaves das Neves, H.J.; Mosandl, A. Differentiation of ten pine species from central Portugal by monoterpene enantiomer-selective composition analysis using multidimensional gas chromatography. Chromatographia 2001, 53, S412. [Google Scholar] [CrossRef]
- Wise, A.; Borg-Karlson, A.-K.; Persson, M.; Norin, T.; Mustaparta, H. Enantiomeric composition of monoterpene hydrocarbons in some conifers and receptor neuron discrimination of α-pinene and limonene enantiomers in the pine weevil. Hylobius Abietis. J. Chem. Ecol. 1998, 24, 273–287. [Google Scholar]
- Borg-Karlson, A.-K.; Unelius, C.R.; Valterová, I.; Nilsson, L.A. Floral fragrance chemistry in the early flowering shrub Daphne mezereum. Phytochemistry 1996, 41, 1477–1483. [Google Scholar] [CrossRef]
- Davis, J.M.; Giddings, J.C. Statistical theory of component overlap in multicomponent chromatograms. Anal. Chem. 1983, 55, 418–424. [Google Scholar] [CrossRef]
- Amigo, J.M.; Skov, T.; Coello, J.; Maspoch, S.; Bro, R. Solving GC-MS problems with PARAFAC2. Trends Anal. Chem. 2008, 27, 714–725. [Google Scholar] [CrossRef]
- Jalali-Heravi, M.; Moazeni-Pourasil, R.S.; Sereshti, H. Elimination of chromatographic and mass spectrometric problems in GC-MS analysis of Lavender essential oil by multivariate curve resolution techniques: Improving the peak purity assessment by variable size moving window-evolving factor analysis. J. Chromatogr. B 2015, 983–984, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Xu, Z.; Zhang, H.; Liu, D.; Yang, Q.; Tao, Q.; Wen, M.; Kang, X.; Zhang, Z.; Lu, H. Fully automatic resolution of untargeted GC-MS data with deep learning assistance. Talanta 2022, 244, 123415. [Google Scholar] [CrossRef] [PubMed]
- Lough, W.J.; Ferguson, P. Meeting Review: Advances in Microcolumn and Related Separation Technologies. Column 2016, 18, 17–24. Available online: https://www.chromatographyonline.com/view/meeting-review-advances-microcolumn-and-related-separation-technologies-1 (accessed on 22 May 2022).
- Kulsing, C.; Nolvachai, Y.; Marriott, P.J. Concepts, selectivity options and experimental design approaches in multidimensional and comprehensive two-dimensional gas chromatography. Trends Anal. Chem. 2020, 130, 115995. [Google Scholar] [CrossRef]
- Liu, Z.; Phillips, J.B. Comprehensive two-dimensional gas chromatography using an on-column thermal modulator interface. J. Chromatogr. Sci. 1991, 29, 227–231. [Google Scholar] [CrossRef]
- Marriott, P.J.; Chin, S.T.; Nolvachai, Y. Techniques and application in comprehensive multidimensional gas chromatography—Mass spectrometry. J. Chromatogr. A 2021, 1636, 461788. [Google Scholar] [CrossRef]
- Johnsen, L.G.; Skou, P.B.; Khakimov, B.; Bro, R. Gas chromatography—Mass spectrometry data processing made easy. J. Chromatogr. A 2021, 1503, 57–64. [Google Scholar] [CrossRef]
- Risum, A.B.; Bro, R. Using deep learning to evaluate peaks in chromatographic data. Talanta 2019, 204, 255–260. [Google Scholar] [CrossRef]
- Lebanov, L.; Tedone, L.; Ghiasvand, A.; Paull, B. Random Forests machine learning applied to gas chromatography—Mass spectrometry derived average mass spectrum data sets for classification and characterisation of essential oils. Talanta 2020, 208, 120471. [Google Scholar] [CrossRef]
- Schneider, D. Elektrophysiologische Untersuchungen von Chemound Mechanoreceptoren de Antenne des Seidenspinners Bombyx mori L. Z. Vergl. Physiol. 1957, 40, 8–41. [Google Scholar] [CrossRef]
- Ryan, M.F. Insect Chemoreception, Fundamental and Applied; Kluwer Academic Publishers: Alphen aan den Rijn, The Netherlands, 2002. [Google Scholar]
- Roelofs, W.L. Electroantennogram assays: Rapid and convenient screening procedures for pheromones. In Techniques in Pheromone Research; Springer: New York, NY, USA, 1984; pp. 131–159. [Google Scholar]
- Bartelt, R. Pheromones of Non-Lepidopteran Insects Associated with Agricultural Plants; CABI Publishing: Oxford, UK, 1999; pp. 91–112. [Google Scholar]
- Weissbecker, B.; Holighaus, G.; Schütz, S. Gas chromatography with mass spectrometric and electroantennographic detection: Analysis of wood odorants by direct coupling of insect olfaction and mass spectrometry. J. Chromatogr. A 2004, 1056, 209–216. [Google Scholar] [CrossRef]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects–finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Barth, H. Chromatography fundamentals, part VIII: The meaning and significance of chromatographic resolution. LCGC N. Am. 2019, 37, 824–882. [Google Scholar]
- Marriott, P.J.; Chin, S.T.; Maikhunthod, B.; Schmarr, H.G.; Bieri, S. Multidimensional Gas Chromatography. Trends Anal. Chem. 2012, 34, 1–21. [Google Scholar] [CrossRef]
- Nolvachai, Y.; Kulsing, C.; Marriott, P.J. Multidimensional gas chromatography in food analysis. Trends Anal. Chem. 2017, 96, 124–137. [Google Scholar] [CrossRef]
- Nolvachai, Y.; McGregor, L.; Spadafora, N.D.; Bukowski, N.P.; Marriott, P.J. Comprehensive two-dimensional gas chromatography with mass spectrometry: Toward a super-resolved separation technique. Anal. Chem. 2020, 92, 12572–12578. [Google Scholar] [CrossRef]
- Deans, D.R. A new Technique for Heart Cutting in Gas Chromatography. Chromatographia 1968, 1, 18–22. [Google Scholar] [CrossRef]
- Tranchida, P.Q.; Sciarrone, D.; Dugo, P.; Mondello, L. Heart-cutting multidimensional gas chromatography: A review of recent evolution, applications, and future prospects. Anal. Chim. Acta 2012, 716, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Tranchida, P.Q.; Aloisi, I.; Mondello, L. Heart-cutting and comprehensive multidimensional gas chromatography: Basic principles. Compr. Anal. Chem. 2022, 96, 69–92. [Google Scholar]
- Mendes, D. Desenvolvimento de um Método de Cromatografia Gasosa Multidimensional para a Determinação de Atributos Olfactivos Negativos em Discos de Cortiça (Development of a Multidimensional Gas Chromatography Method for the Determination of Negative Olfactory Attributes in Cork Discs). Bachelor’s Thesis, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, Caparica, Portugal, 2016. [Google Scholar]
- Pavlova, A.; Mikhalchenko, O.; Rogachev, A.; Il’ina, I.; Korchagina, D.; Gatilov, Y.; Tolstikova, T.; Volcho, K.; Salakhutdinov, N. Synthesis and analgesic activity of stereoisomers of 2-(3(4)-hydroxy-4(3)-methoxyphenyl)-4,7-dimethyl-3,4,4a,5,8,8a-hexahydro-2H-chromene-4,8-diols. Med. Chem. Res. 2015, 24, 3821–3830. [Google Scholar] [CrossRef]
- Gemal, A.L.; Luche, J.-L. Lanthanoids in organic synthesis. 6. Reduction of. alpha-enones by sodium borohydride in the presence of lanthanoid chlorides: Synthetic and mechanistic aspects. J. Am. Chem. Soc. 1981, 103, 5454–5459. [Google Scholar] [CrossRef]
- Uzarewicz, A.; Dresler, R. The reaction of cyclic allylic alcohols with aliphatic alcohols in the presence of cerium (III) chloride. Pol. J. Chem. 1997, 71, 181–195. [Google Scholar] [CrossRef]
- Schaffarczyk, M.; Balaban, T.S.; Rychlik, M.; Buettner, A. Syntheses of chiral 1,8-Cineole metabolites and determination of their enantiomeric composition in human urine after ingestion of 1,8-Cineole-containing capsules. ChemPlusChem 2013, 78, 77–85. [Google Scholar] [CrossRef]
- Horst, K.; Rychlik, M. Quantification of 1,8-cineole and of its metabolites in humans using stable isotope dilution assays. Mol. Nutr. Food Res. 2010, 54, 1515–1529. [Google Scholar] [CrossRef]
- Joulain, D.; Konig, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; EB Verlag: Hamburg, Germany, 1998. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2001. [Google Scholar]
- Kirsch, F.; Horst, K.; Röhrig, W.; Rychlik, M.; Buettner, A. Tracing metabolite profiles in human milk: Studies on the odorant 1,8-cineole transferred into breast milk after oral intake. Metabolomics 2013, 9, 483–496. [Google Scholar] [CrossRef]
- El-Sayed, A.M. The Pherobase: Database of Pheromones and Semiochemicals. 2022. Available online: http://www.pherobase.com (accessed on 21 May 2022).
- Mateus, E.; Barata, R.C.; Zrostlíková, J.; da Silva, M.G.; Paiva, M.R. Characterization of the volatile fraction emitted by Pinus spp by one and two dimensional chromatographic techniques with mass spectrometric detection. J. Chromtogr. A 2010, 1217, 1845–1855. [Google Scholar] [CrossRef]
- Sarracino, F.; Brinco, J.; Gomes da Silva, M.; Ferreira, R.B.; Chagas, R.; Ferreira, L.M. DCMC as a new white wine stabilizing agent. Impact on protein stability and wine aromatic fraction against bentonite. Molecules 2021, 26, 6188. [Google Scholar] [CrossRef]
- Van den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas—Liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Sauer, A.E.; Karg, G.; Koch, U.T.; De Kramer, J.J.; Milli, R. A portable EAG system for the measurement of pheromone concentrations in the field. Chem. Senses 1992, 17, 543–553. [Google Scholar] [CrossRef]



| Compound No. | Putative ID | RT Innowax | E. globulus | E. globulus + ♂ | ♂ | Standard |
|---|---|---|---|---|---|---|
| 1 | α-Pinene | 6.11 | + | + | + | + |
| 2 | Camphene | 6.33 | + | + | + | + |
| 3 | Methyl 2-methylpentanoate | 6.59 | + | - | - | - |
| 4 | β-Pinene | 6.84 | + | + | + | + |
| 5 | Verbenene | 7.33 | - | + | + | + |
| 6 | β-Myrcene | 7.78 | + * | + | + | + |
| 7 | α-Phellandrene | 7.88 | + * | + | + | + |
| 8 | α-Terpinene | 8.23 | + * | + | + | - |
| 9 | Limonene | 8.63 | + * | + | + | + |
| 10 | 1,8-Cineole | 8.95 | + * | + | + | + |
| 11 | γ-Terpinene | 9.49 | + | + | + | + |
| 12 | p-Cymene | 9.91 | + | + | + | + |
| 13 | α-Terpinolene | 10.11 | + | + | + | + |
| 14 | neo-allo-Ocimene | 10.45 | + | + | - | + |
| 15 | (E)-4,8-Dimethyl-1,3,7-nonatriene | 10.44 | - | + | - | - |
| 16 | (Z)-3-Hexen-1-yl acetate | 10.61 | + | |||
| 17 | 6-Methyl-5-hepten-2-one | 11.01 | + | + | + | - |
| 18 | (Z)-3-Hexen-1-ol | 11.69 | + | + | + | + |
| 19 | Nonanal | 11.98 | + | + | + | - |
| 20 | 1,3,8-p-Menthatriene | 12.68 | - | |||
| 21 | cis-Linalool oxide | 12.81 | + | + | + | + |
| 22 | trans-Sabinene hydrate | 13.08 | + | + | + | + |
| 23 | Bicycloelemene | 13.55 | + | + | + | - |
| 24 | α-Copaene | 13.78 | + | + | + | + |
| 25 | α-Gurjunene | 14.42 | + | + | + | + |
| 26 | Fenchol | 15.09 | + | - | - | + |
| 27 | Calarene | 15.49 | + | + | + | + |
| 28 | β-Caryophyllene | 15.57 | - | + | + | + |
| 29 | Aromadendrene | 15.79 | + | + * | + | + |
| 30 | allo-Aromadendrene | 16.33 | + * | + * | + * | + |
| 31 | trans-Pinocarveol | 16.34 | + * | + * | + * | + |
| 32 | cis-Verbenol | 16.35 | - | + * | + * | + |
| 33 | δ-Terpineol | 16.52 | + | + | - | - |
| 34 | trans-Verbenol | 16.68 | - | + | + | + |
| 35 | α-Terpineol | 16.94 | + * | + * | + * | + |
| 36 | α-Terpinyl acetate | 17.02 | + * | + * | + * | + |
| 37 | Ledene | 17.11 | + * | + * | + * | + |
| 38 | 2-Oxo-1,8-cineole | 17.20 | - | + | + | + |
| 39 | Verbenone | 17.31 | - | + | + | + |
| 40 | exo-2-Hydroxycineole acetate | 17.42 | + | - | - | - |
| 41 | 2-β-Hydroxy-8-cineole | 17.44 | - | + | + | - |
| 42 | Bicyclogermacrene | 17.66 | + | + | + | - |
| 43 | δ-Cadinene | 17.93 | + | + | - | - |
| 44 | Ethyl 2-phenylethanoate | 18.30 | + | - | - | - |
| 45 | 7-Hydroxy-1,8-cineole | 18.36 | - | + * | + * | - |
| 46 | Myrtenol | 18.38 | + | + * | + * | + |
| 47 | 2-Phenethyl acetate | 18.74 | + | + | + | - |
| 48 | trans-Carveol | 18.90 | + | + | - | + |
| 49 | Geraniol | 19.00 | + | + | - | + |
| 50 | 2-α-Hydroxy-1,8-cineole | 19.31 | - | + * | + * | + |
| 51 | 9-Hydroxy-1,8-cineole | 19.40 | - | + * | + * | + |
| 52 | Benzyl alcohol | 19.56 | + | - | - | - |
| 53 | 3-α-Hydroxy-1,8-cineole | 19.96 | - | + | + | - |
| 54 | 2-Phenylethanol | 20.05 | + | + | - | + |
| 55 | Phenylethyl isovalerate | 21.13 | + | + | + | + |
| 56 | epi-Globulol | 21.48 | + | + | + | + |
| 57 | Globulol | 22.32 | + | + | - | + |
| 58 | Rosifoliol | 22.65 | + | + | - | - |
| 59 | Spathulenol | 22.89 | + | + | + | - |
| 60 | tau-Cadinol | 23.42 | + | + | - | - |
| 61 | iso-Spathulenol | 24.01 | + | + | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, D.; Branco, S.; Paiva, M.R.; Schütz, S.; Mateus, E.P.; Gomes da Silva, M. Unveiling Chemical Cues of Insect-Tree and Insect-Insect Interactions for the Eucalyptus Weevil and Its Egg Parasitoid by Multidimensional Gas Chromatographic Methods. Molecules 2022, 27, 4042. https://doi.org/10.3390/molecules27134042
Mendes D, Branco S, Paiva MR, Schütz S, Mateus EP, Gomes da Silva M. Unveiling Chemical Cues of Insect-Tree and Insect-Insect Interactions for the Eucalyptus Weevil and Its Egg Parasitoid by Multidimensional Gas Chromatographic Methods. Molecules. 2022; 27(13):4042. https://doi.org/10.3390/molecules27134042
Chicago/Turabian StyleMendes, Davide, Sofia Branco, Maria Rosa Paiva, Stefan Schütz, Eduardo P. Mateus, and Marco Gomes da Silva. 2022. "Unveiling Chemical Cues of Insect-Tree and Insect-Insect Interactions for the Eucalyptus Weevil and Its Egg Parasitoid by Multidimensional Gas Chromatographic Methods" Molecules 27, no. 13: 4042. https://doi.org/10.3390/molecules27134042
APA StyleMendes, D., Branco, S., Paiva, M. R., Schütz, S., Mateus, E. P., & Gomes da Silva, M. (2022). Unveiling Chemical Cues of Insect-Tree and Insect-Insect Interactions for the Eucalyptus Weevil and Its Egg Parasitoid by Multidimensional Gas Chromatographic Methods. Molecules, 27(13), 4042. https://doi.org/10.3390/molecules27134042