
Coumarin Derivatives Inhibit ADP-Induced Platelet Activation and Aggregation

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
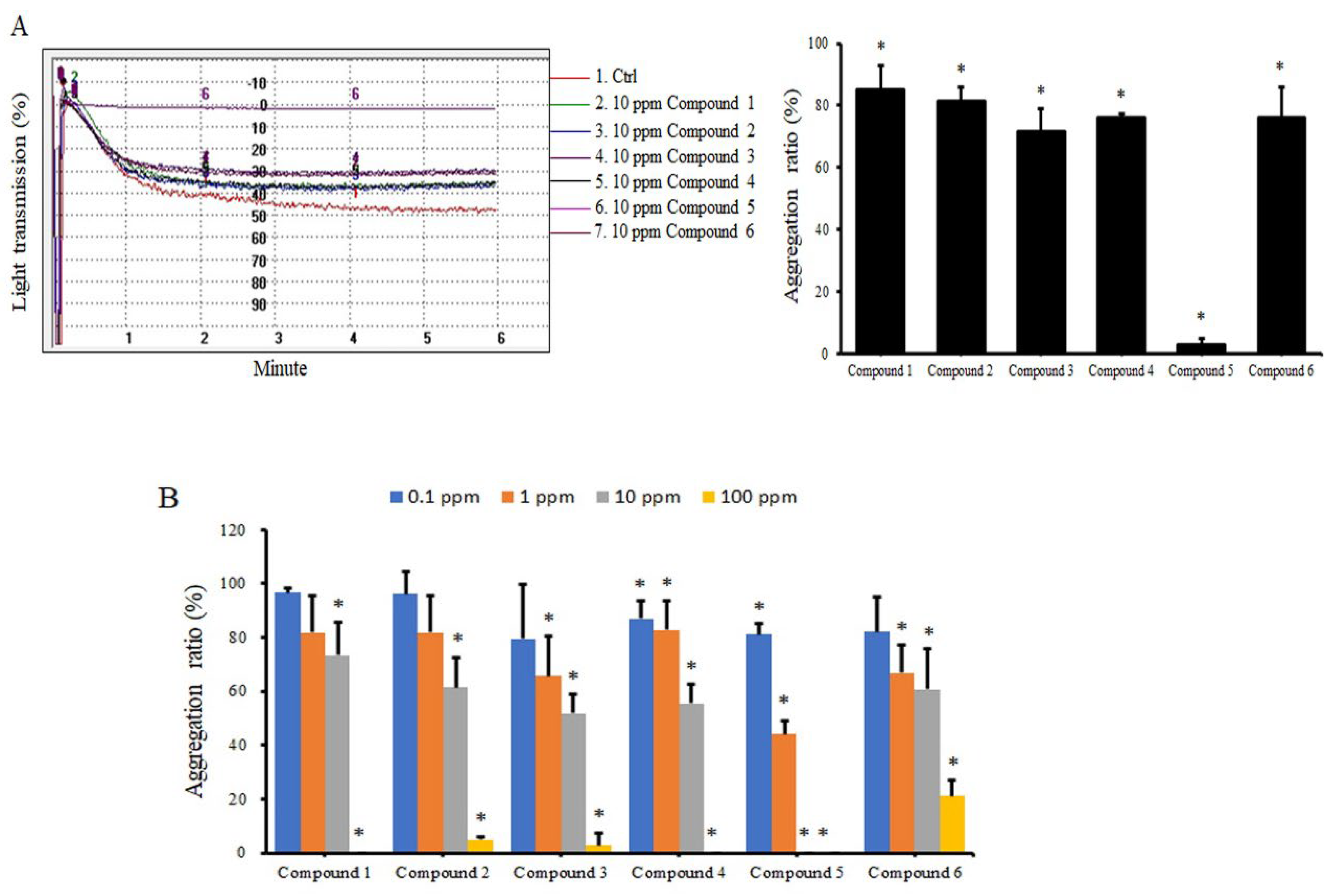
2.1. Coumarin Derivatives Inhibit ADP-Induced Platelet Aggregation
2.2. Coumarin Derivatives Inhibit Expression of Glycoproteins on Platelet Membranes
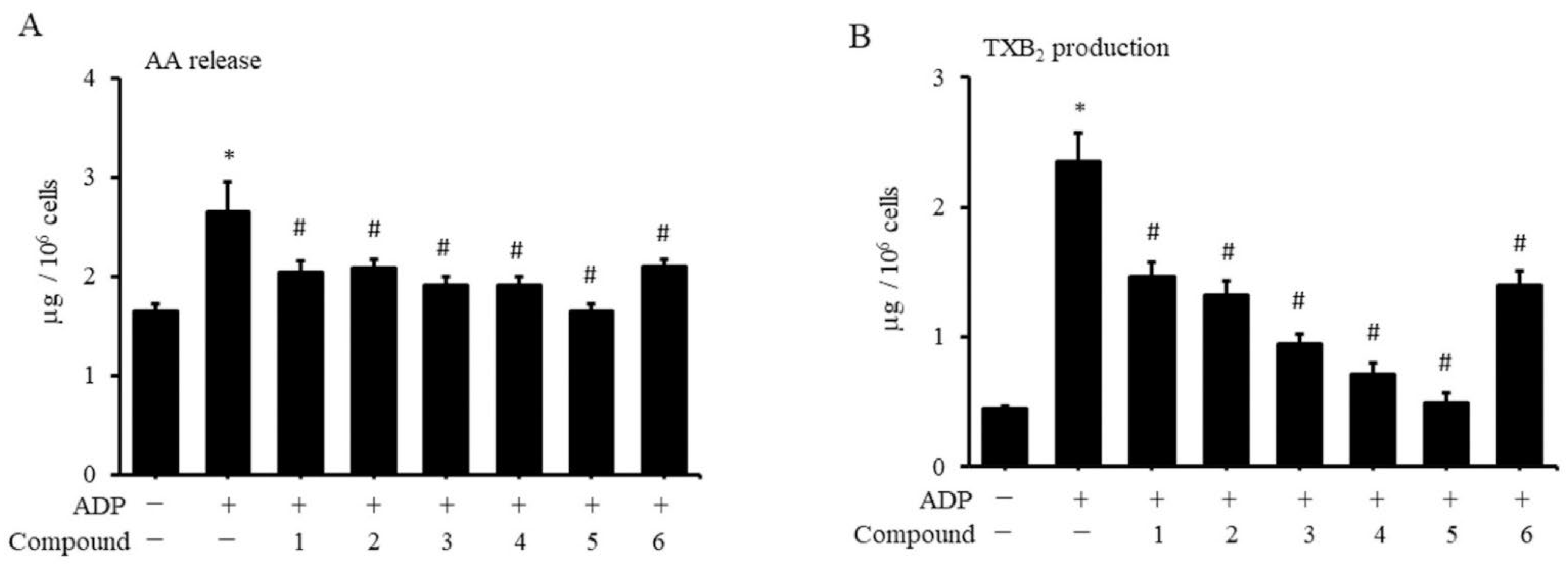
2.3. Coumarin Derivatives Inhibit Platelet Production of AA and TXA2
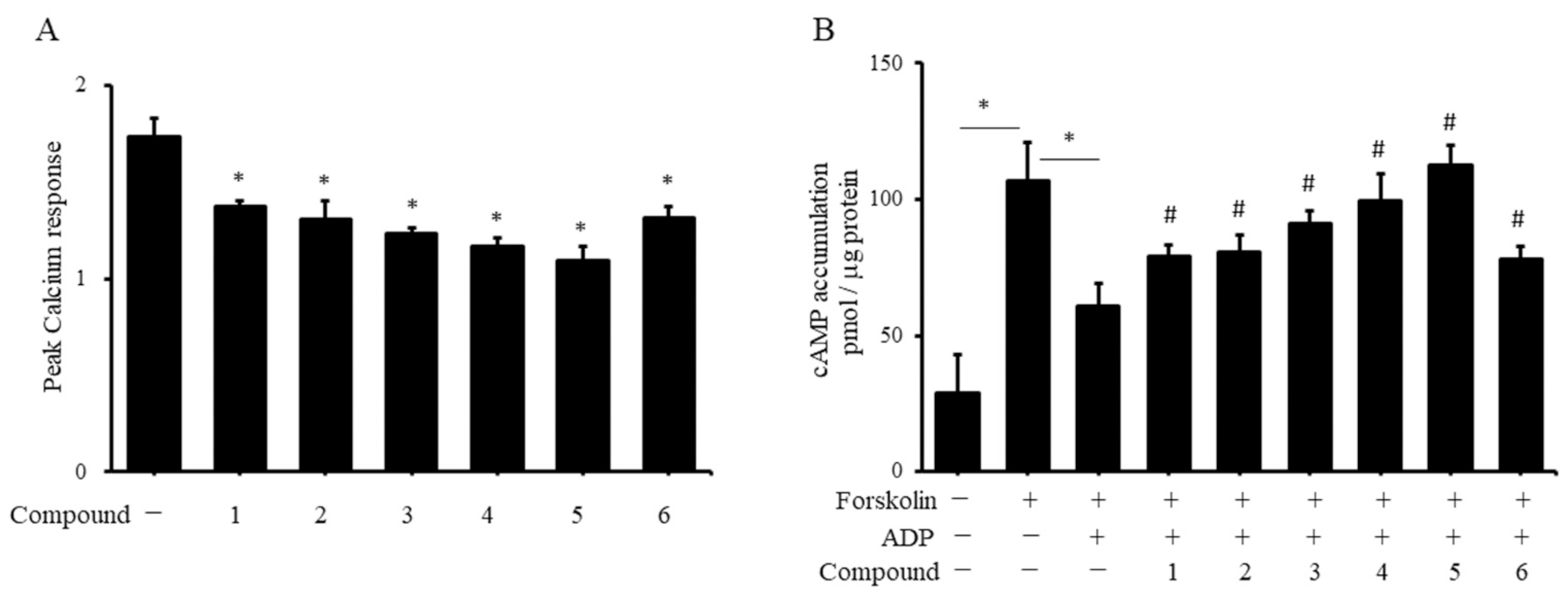
2.4. Coumarin Derivatives Inhibit Platelet ADP Receptor Signaling
3. Discussion
4. Materials and Methods
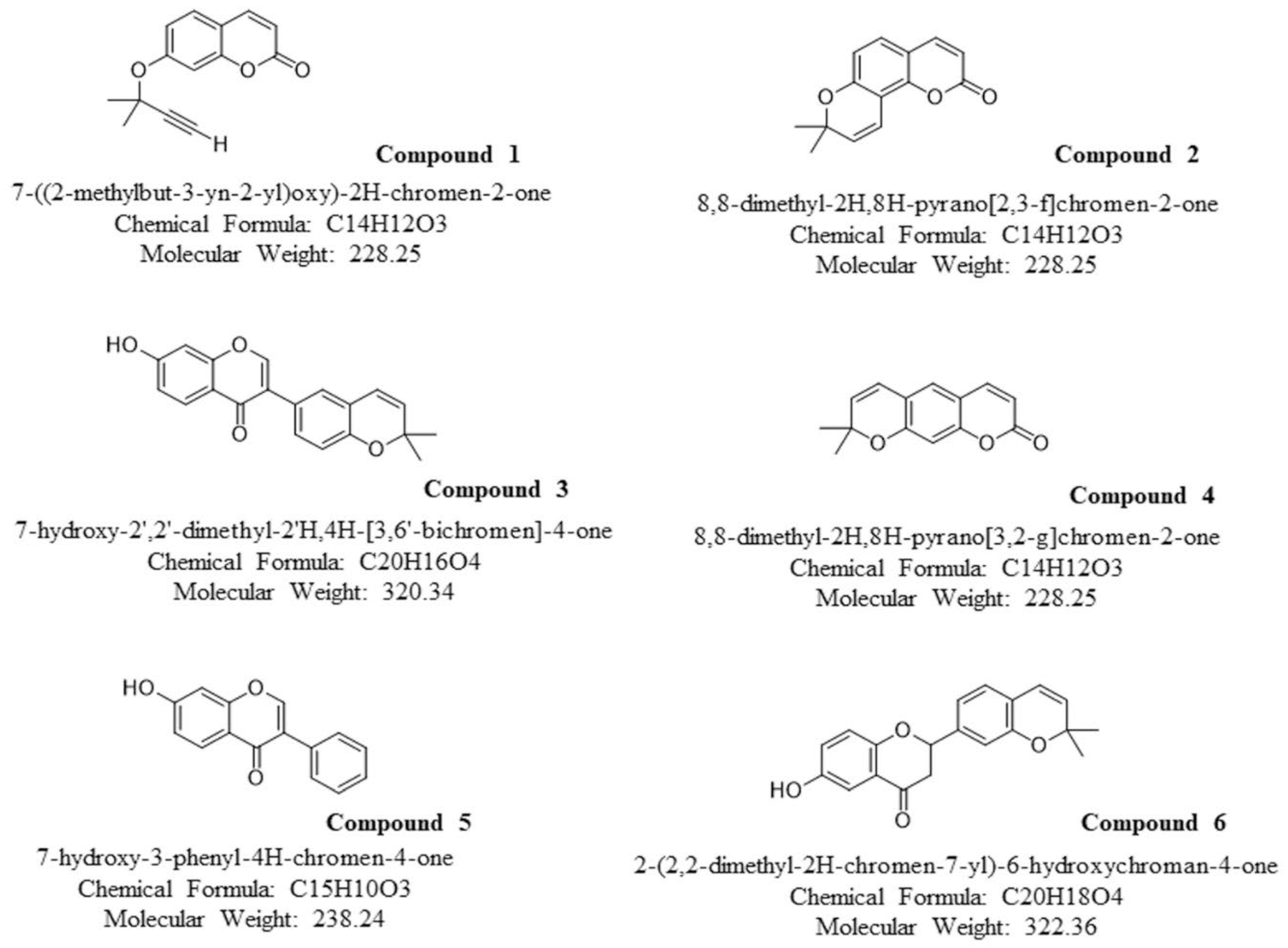
4.1. Coumarin Derivatives
4.2. Preparation of Platelet-Rich Plasma (PRP), Platelet-Poor Plasma (PPP), and Purified Platelets
4.3. Measurement of Platelet Aggregation
4.4. Measurement of Platelet Surface Glycoproteins
4.5. Measurement of Arachidonic-Acid (AA) Release from Platelets
4.6. Measurement of Thromboxane A2 (TXA2) in Platelets
4.7. Measurement of Cytosolic Free Calcium in Platelets
4.8. Measurement of cAMP Content in Platelets
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loncar, M.; Jakovljevic, M.; Subaric, D.; Pavlic, M.; Buzjak, S.V.; Cindric, I.; Molnar, M. Coumarins in Food and Methods of Their Determination. Foods 2020, 9, 645. [Google Scholar] [CrossRef] [PubMed]
- Kuruvilla, M.; Gurk-Turner, C. A review of warfarin dosing and monitoring. Proc. Bayl. Univ. Med. Cent. 2001, 14, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Hirsh, J.; Fuster, V.; Ansell, J.; Halperin, J. AHA/ACC Scientific Statement. American College of Cardiology F: American Heart Association/American College of Cardiology Foundation guide to warfarin therapy. Circulation 2003, 107, 1692–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.S.; Lai, N.M.; Nathisuwan, S.; Jahan, N.K.; Dilokthornsakul, P.; Kongpakwattana, K.; Hollingworth, W.; Chaiyakunapruk, N. Comparative efficacy and safety of warfarin care bundles and novel oral anticoagulants in patients with atrial fibrillation: A systematic review and network meta-analysis. Sci. Rep. 2020, 10, 662. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, A.M.; Pereira, J.A.; Labiris, R.; McDonald, H.; Douketis, J.D.; Crowther, M.; Wells, P.S. Systematic overview of warfarin and its drug and food interactions. Arch. Intern. Med. 2005, 165, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Palta, S.; Saroa, R.; Palta, A. Overview of the coagulation system. Indian J. Anaesth. 2014, 58, 515–523. [Google Scholar] [CrossRef]
- Periayah, M.H.; Halim, A.S.; Mat Saad, A.Z. Mechanism Action of Platelets and Crucial Blood Coagulation Pathways in Hemostasis. Int. J. Hematol. Oncol. Stem Cell Res. 2017, 11, 319–327. [Google Scholar]
- Versteeg, H.H.; Heemskerk, J.W.; Levi, M.; Reitsma, P.H. New fundamentals in hemostasis. Physiol. Rev. 2013, 93, 327–358. [Google Scholar] [CrossRef] [Green Version]
- Bonar, R.A.; Lippi, G.; Favaloro, E.J. Overview of Hemostasis and Thrombosis and Contribution of Laboratory Testing to Diagnosis and Management of Hemostasis and Thrombosis Disorders. Methods Mol. Biol. 2017, 1646, 3–27. [Google Scholar]
- Miyata, T. Guest editorial: Current understanding of thrombosis and hemostasis–from bench to bedside. Int. J. Hematol. 2012, 95, 331–332. [Google Scholar] [CrossRef] [Green Version]
- van der Meijden, P.E.J.; Heemskerk, J.W.M. Platelet biology and functions: New concepts and clinical perspectives. Nat. Rev. Cardiol. 2019, 16, 166–179. [Google Scholar] [CrossRef]
- Yun, S.H.; Sim, E.H.; Goh, R.Y.; Park, J.I.; Han, J.Y. Platelet Activation: The Mechanisms and Potential Biomarkers. Biomed. Res. Int. 2016, 2016, 9060143. [Google Scholar] [CrossRef] [Green Version]
- Nieswandt, B.; Pleines, I.; Bender, M. Platelet adhesion and activation mechanisms in arterial thrombosis and ischaemic stroke. J. Thromb. Haemost. 2011, 9 (Suppl. 1), 92–104. [Google Scholar] [CrossRef]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [Green Version]
- Swieringa, F.; Spronk, H.M.H.; Heemskerk, J.W.M.; van der Meijden, P.E.J. Integrating platelet and coagulation activation in fibrin clot formation. Res. Pract. Thromb. Haemost. 2018, 2, 450–460. [Google Scholar] [CrossRef]
- Woulfe, D.; Yang, J.; Brass, L. ADP and platelets: The end of the beginning. J. Clin. Investig. 2001, 107, 1503–1505. [Google Scholar] [CrossRef]
- Bierend, A.; Rau, T.; Maas, R.; Schwedhelm, E.; Boger, R.H. P2Y12 polymorphisms and antiplatelet effects of aspirin in patients with coronary artery disease. Br. J. Clin. Pharmacol. 2008, 65, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Koessler, J.; Schwarz, M.; Weber, K.; Etzel, J.; Koessler, A.; Boeck, M.; Kobsar, A. The role of adenosine diphosphate mediated platelet responsiveness for the stability of platelet integrity in citrated whole blood under ex vivo conditions. PLoS ONE 2017, 12, e0188193. [Google Scholar] [CrossRef] [Green Version]
- Daniel, J.L.; Dangelmaier, C.; Jin, J.; Ashby, B.; Smith, J.B.; Kunapuli, S.P. Molecular basis for ADP-induced platelet activation. I. Evidence for three distinct ADP receptors on human platelets. J. Biol. Chem. 1998, 273, 2024–2029. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Daniel, J.L.; Kunapuli, S.P. Molecular basis for ADP-induced platelet activation. II. The P2Y1 receptor mediates ADP-induced intracellular calcium mobilization and shape change in platelets. J. Biol. Chem. 1998, 273, 2030–2034. [Google Scholar] [CrossRef] [Green Version]
- Offermanns, S. Activation of platelet function through G protein-coupled receptors. Circ. Res. 2006, 99, 1293–1304. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ju, L.A.; Zhou, F.; Liao, J.; Xue, L.; Su, Q.P.; Jin, D.; Yuan, Y.; Lu, H.; Jackson, S.P.; et al. An integrin alphaIIbbeta3 intermediate affinity state mediates biomechanical platelet aggregation. Nat. Mater. 2019, 18, 760–769. [Google Scholar] [CrossRef]
- O’Reilly, R.A. Studies on the coumarin anticoagulant drugs: Interaction of human plasma albumin and warfarin sodium. J. Clin. Investig. 1967, 46, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Whitlon, D.S.; Sadowski, J.A.; Suttie, J.W. Mechanism of coumarin action: Significance of vitamin K epoxide reductase inhibition. Biochemistry 1978, 17, 1371–1377. [Google Scholar] [CrossRef]
- Akoudad, S.; Darweesh, S.K.; Leening, M.J.; Koudstaal, P.J.; Hofman, A.; van der Lugt, A.; Stricker, B.H.; Ikram, M.A.; Vernooij, M.W. Use of coumarin anticoagulants and cerebral microbleeds in the general population. Stroke 2014, 45, 3436–3439. [Google Scholar] [CrossRef] [Green Version]
- Verhoef, T.I.; Redekop, W.K.; Hasrat, F.; de Boer, A.; Maitland-van der Zee, A.H. Cost effectiveness of new oral anticoagulants for stroke prevention in patients with atrial fibrillation in two different European healthcare settings. Am. J. Cardiovasc. Drugs 2014, 14, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.L.; Adams, M.; Ravi, R.G.; Jacobson, K.A.; Harden, T.K. 2-Chloro N(6)-methyl-(N)-methanocarba-2’-deoxyadenosine-3′,5′-bisphosphate is a selective high affinity P2Y(1) receptor antagonist. Br. J. Pharmacol. 2002, 135, 2004–2010. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.A.; Reimann, S.; Schror, K. Specific inhibition of ADP-induced platelet aggregation by clopidogrel in vitro. Br. J. Pharmacol. 1999, 126, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna Kumar, D.; Gopal Shewade, D.; Parasuraman, S.; Rajan, S.; Balachander, J.; Sai Chandran, B.V.; Adithan, C. Estimation of plasma levels of warfarin and 7-hydroxy warfarin by high performance liquid chromatography in patients receiving warfarin therapy. J. Young Pharm. 2013, 5, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Faggio, C.; Sureda, A.; Morabito, S.; Sanches-Silva, A.; Mocan, A.; Nabavi, S.F.; Nabavi, S.M. Flavonoids and platelet aggregation: A brief review. Eur. J. Pharmacol. 2017, 807, 91–101. [Google Scholar] [CrossRef]
- Pignatelli, P.; Pulcinelli, F.M.; Celestini, A.; Lenti, L.; Ghiselli, A.; Gazzaniga, P.P.; Violi, F. The flavonoids quercetin and catechin synergistically inhibit platelet function by antagonizing the intracellular production of hydrogen peroxide. Am. J. Clin. Nutr. 2000, 72, 1150–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, B.; Spencer, J.P.; Lovegrove, J.A.; Gibbins, J.M. Flavonoid inhibitory pharmacodynamics on platelet function in physiological environments. Food Funct. 2013, 4, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, S.H.; Ko, W.C.; Ko, F.N.; Teng, C.M. Inhibition of platelet aggregation by some flavonoids. Thromb. Res. 1991, 64, 91–100. [Google Scholar] [CrossRef]
- Karlickova, J.; Riha, M.; Filipsky, T.; Macakova, K.; Hrdina, R.; Mladenka, P. Antiplatelet Effects of Flavonoids Mediated by Inhibition of Arachidonic Acid Based Pathway. Planta Med. 2016, 82, 76–83. [Google Scholar]
- Sangkuhl, K.; Shuldiner, A.R.; Klein, T.E.; Altman, R.B. Platelet aggregation pathway. Pharmacogenet. Genom. 2011, 21, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Ye, J.; Zhang, Y.; Xu, X.; Liu, J.; Zhang, S.H.; Kunapuli, S.P.; Ding, Z. P2Y12 protects platelets from apoptosis via PI3k-dependent Bak/Bax inactivation. J. Thromb. Haemost. 2013, 11, 149–160. [Google Scholar] [CrossRef]
- Kostrzewa-Suslow, E.; Janeczko, T. Microbial transformations of 7-hydroxyflavanone. Sci. World J. 2012, 2012, 254929. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Su, H.; Zhang, T.; Du, J.; Cui, S.; Yang, F.; Jin, Q. Inhibition of Enterovirus 71 replication by 7-hydroxyflavone and diisopropyl-flavon7-yl Phosphate. PLoS ONE 2014, 9, e92565. [Google Scholar] [CrossRef]
- Jin, Z.; Yang, Y.Z.; Chen, J.X.; Tang, Y.Z. Inhibition of pro-inflammatory mediators in RAW264.7 cells by 7-hydroxyflavone and 7,8-dihydroxyflavone. J. Pharm. Pharmacol. 2017, 69, 865–874. [Google Scholar] [CrossRef]
- Stompor, M.; Switalska, M.; Wietrzyk, J. Synthesis and biological evaluation of acyl derivatives of hydroxyflavones as potent antiproliferative agents against drug resistance cell lines. Z. Nat. C 2018, 73, 87–93. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, P.-H.; Liao, T.-H.; Chen, Y.-H.; Hsu, Y.-L.; Kuo, C.-Y.; Chan, C.-C.; Wang, L.-K.; Chern, C.-Y.; Tsai, F.-M. Coumarin Derivatives Inhibit ADP-Induced Platelet Activation and Aggregation. Molecules 2022, 27, 4054. https://doi.org/10.3390/molecules27134054
Lu P-H, Liao T-H, Chen Y-H, Hsu Y-L, Kuo C-Y, Chan C-C, Wang L-K, Chern C-Y, Tsai F-M. Coumarin Derivatives Inhibit ADP-Induced Platelet Activation and Aggregation. Molecules. 2022; 27(13):4054. https://doi.org/10.3390/molecules27134054
Chicago/Turabian StyleLu, Ping-Hsun, Tzu-Hsien Liao, Yau-Hung Chen, Yeng-Ling Hsu, Chan-Yen Kuo, Chuan-Chi Chan, Lu-Kai Wang, Ching-Yuh Chern, and Fu-Ming Tsai. 2022. "Coumarin Derivatives Inhibit ADP-Induced Platelet Activation and Aggregation" Molecules 27, no. 13: 4054. https://doi.org/10.3390/molecules27134054
APA StyleLu, P. -H., Liao, T. -H., Chen, Y. -H., Hsu, Y. -L., Kuo, C. -Y., Chan, C. -C., Wang, L. -K., Chern, C. -Y., & Tsai, F. -M. (2022). Coumarin Derivatives Inhibit ADP-Induced Platelet Activation and Aggregation. Molecules, 27(13), 4054. https://doi.org/10.3390/molecules27134054