
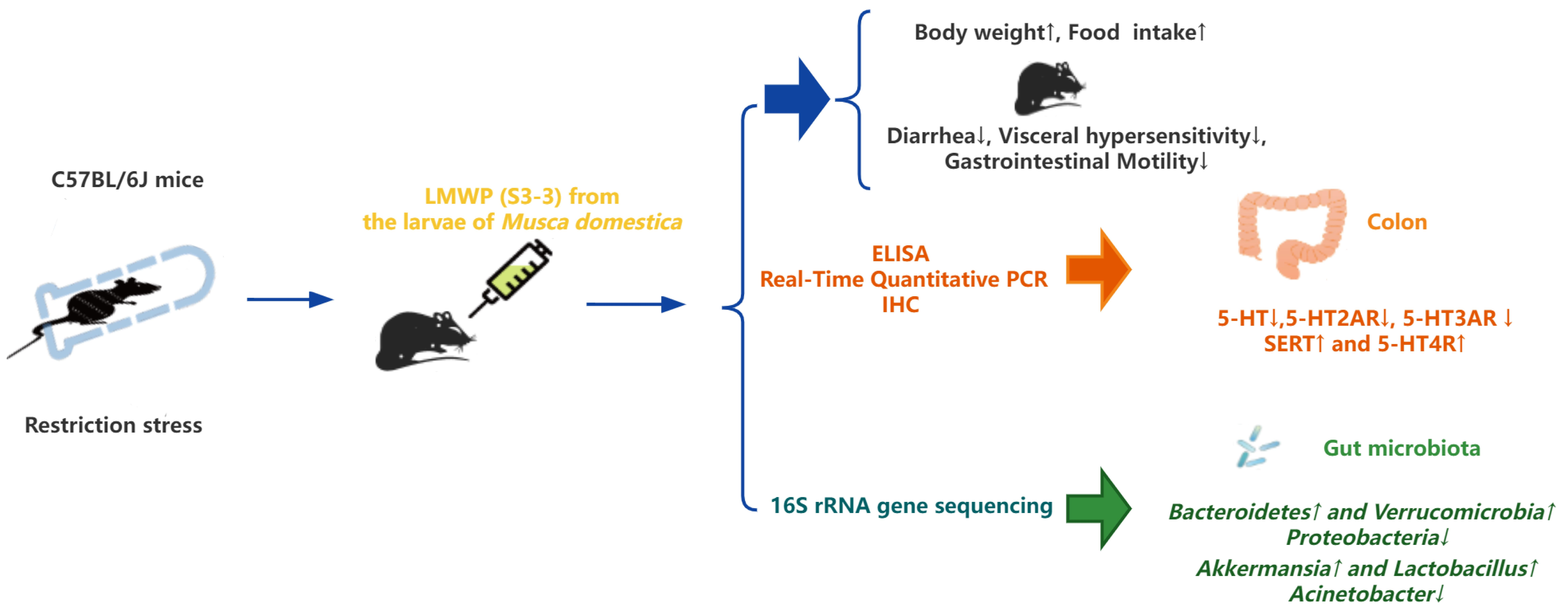
LMWP (S3-3) from the Larvae of Musca domestica Alleviate D-IBS by Adjusting the Gut Microbiota

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of LMWP (S3-3) from the Larvae of Musca domestica
2.2. Animals and Experimental Design
2.3. Introduction of D-IBS in Mice
2.4. Determination of the Rate of Loose Stool
2.5. The Abdominal Retraction Reflex (AWR) Score
2.6. Colonic Bead Expulsion Test
2.7. Upper GI Transit
2.8. Gastric Emptying
2.9. Histological Analysis
2.10. Enzyme-Linked Immunoassay (ELISA)
2.11. Real-Time Quantitative PCR Detection
2.12. Immunohistochemistry (IHC)
2.13. Gut Microbiota Analysis
2.14. Statistical Analysis
3. Results
3.1. Effects of LMWP (S3-3) from Larvae of Musca domestica on Physiological Conditions and the Frequency of Loose Stools in D-IBS Mice
3.2. Effects of LMWP (S3-3) from Larvae of Musca domestica on Gastrointestinal Motility and Visceral Sensitivity in Mice
3.3. Effect of LMWP (S3-3) from Larvae of Musca domestica on Colonic Histological Assessment
3.4. Effect of LMWP (S3-3) from Larvae of Musca domestica on the Expression of Genes and Proteins Involved in 5-HT-Related Pathways
3.5. Effects of LMWP (S3-3) from Larvae of Musca domestica on Gut Microbiota in D-IBS Mice
3.6. Potential Correlations among Phenotypes, Molecular Biology Indicators and Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lee, S.H.; Kim, K.N.; Kim, K.M.; Joo, N.S. Irritable Bowel Syndrome May Be Associated with Elevated Alanine Aminotransferase and Metabolic Syndrome. Yonsei Med. J. 2016, 57, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Lovell, R.M.; Ford, A.C. Global prevalence of and risk factors for irritable bowel syndrome: A meta-analysis. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2012, 10, 712–721.e714. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.G.; Jung, H.K. Health related quality of life in functional gastrointestinal disorders in Asia. J. Neurogastroenterol. Motil. 2011, 17, 245–251. [Google Scholar] [CrossRef]
- Gao, X.; Qin, Q.; Yu, X.; Liu, K.; Li, L.; Qiao, H.; Zhu, B. Acupuncture at heterotopic acupoints facilitates distal colonic motility via activating M3 receptors and somatic afferent C-fibers in normal, constipated, or diarrhoeic rats. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2015, 27, 1817–1830. [Google Scholar] [CrossRef] [PubMed]
- Samarajeewa, A.D.; Hammad, A.; Masson, L.; Khan, I.U.; Scroggins, R.; Beaudette, L.A. Comparative assessment of next-generation sequencing, denaturing gradient gel electrophoresis, clonal restriction fragment length polymorphism and cloning-sequencing as methods for characterizing commercial microbial consortia. J. Microbiol. Methods 2015, 108, 103–111. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Canavan, C.; West, J.; Card, T. Review article: The economic impact of the irritable bowel syndrome. Aliment. Pharmacol. Ther. 2014, 40, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Adriani, A.; Ribaldone, D.G.; Astegiano, M.; Durazzo, M.; Saracco, G.M.; Pellicano, R. Irritable bowel syndrome: The clinical approach. Panminerva Med. 2018, 60, 213–222. [Google Scholar] [CrossRef]
- Wilson, A.; Longstreth, G.F.; Knight, K.; Wong, J.; Wade, S.; Chiou, C.F.; Barghout, V.; Frech, F.; Ofman, J.J. Quality of life in managed care patients with irritable bowel syndrome. Manag. Care Interface 2004, 17, 24–28, 34. [Google Scholar]
- Ai, H.; Wang, F.; Xia, Y.; Chen, X.; Lei, C. Antioxidant, antifungal and antiviral activities of chitosan from the larvae of housefly, Musca domestica L. Food Chem. 2012, 132, 493–498. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Castro, J.; Grundy, L.; Schober, G.; Garcia-Caraballo, S.; Zhao, T.; Herzig, V.; King, G.F.; Brierley, S.M.; Lewis, R.J. A spider-venom peptide with multitarget activity on sodium and calcium channels alleviates chronic visceral pain in a model of irritable bowel syndrome. Pain 2021, 162, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Rahabi, M.; Salon, M.; Bruno-Bonnet, C.; Prat, M.; Jacquemin, G.; Benmoussa, K.; Alaeddine, M.; Parny, M.; Bernad, J.; Bertrand, B.; et al. Bioactive fish collagen peptides weaken intestinal inflammation by orienting colonic macrophages phenotype through mannose receptor activation. Eur. J. Nutr. 2022, 61, 2051–2066. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gui, S.; Liang, Z.; Liu, A.; Chen, Z.; Tang, Y.; Xiao, M.; Chu, F.; Liu, W.; Jin, X.; et al. Musca domestica Cecropin (Mdc) Alleviates Salmonella typhimurium-Induced Colonic Mucosal Barrier Impairment: Associating With Inflammatory and Oxidative Stress Response, Tight Junction as Well as Intestinal Flora. Front. Microbiol. 2019, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Chu, F.; Jin, X.; Ma, H. Anti-diarrhea effects and identification of Musca domestica larvae low molecular weight peptides (LMWP). J. Pharm. Biomed. Anal. 2019, 173, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Lin, H.T.; Wu, G.J.; Tsai, G.J. pH Effects on solubility, zeta potential, and correlation between antibacterial activity and molecular weight of chitosan. Carbohydr. Polym. 2015, 134, 74–81. [Google Scholar] [CrossRef]
- Hall, H.N.; Masey O’Neill, H.V.; Scholey, D.; Burton, E.; Dickinson, M.; Fitches, E.C. Amino acid digestibility of larval meal (Musca domestica) for broiler chickens. Poult. Sci. 2018, 97, 1290–1297. [Google Scholar] [CrossRef]
- Pei, Z.; Sun, X.; Tang, Y.; Wang, K.; Gao, Y.; Ma, H. Cloning, expression, and purification of a new antimicrobial peptide gene from Musca domestica larva. Gene 2014, 549, 41–45. [Google Scholar] [CrossRef]
- Li, L.; Cui, H.; Li, T.; Qi, J.; Chen, H.; Gao, F.; Tian, X.; Mu, Y.; He, R.; Lv, S.; et al. Synergistic Effect of Berberine-Based Chinese Medicine Assembled Nanostructures on Diarrhea-Predominant Irritable Bowel Syndrome In Vivo. Front. Pharmacol. 2020, 11, 1210. [Google Scholar] [CrossRef]
- Zhu, H.M.; Li, L.; Li, S.Y.; Yan, Q.; Li, F. Effect of water extract from Berberis heteropoda Schrenk roots on diarrhea-predominant irritable bowel syndrome by adjusting intestinal flora. J. Ethnopharmacol. 2019, 237, 182–191. [Google Scholar] [CrossRef]
- Lu, Y.; Huang, J.; Zhang, Y.; Huang, Z.; Yan, W.; Zhou, T.; Wang, Z.; Liao, L.; Cao, H.; Tan, B. Therapeutic Effects of Berberine Hydrochloride on Stress-Induced Diarrhea-Predominant Irritable Bowel Syndrome Rats by Inhibiting Neurotransmission in Colonic Smooth Muscle. Front. Pharmacol. 2021, 12, 596686. [Google Scholar] [CrossRef]
- Bülbül, M.; Sinen, O.; Bayramoğlu, O.; Akkoyunlu, G. Enteric apelin enhances the stress-induced stimulation of colonic motor functions. Stress (Amst. Neth.) 2020, 23, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Agostini, S.; Goubern, M.; Tondereau, V.; Salvador-Cartier, C.; Bezirard, V.; Lévèque, M.; Keränen, H.; Theodorou, V.; Bourdu-Naturel, S.; Goupil-Feuillerat, N.; et al. A marketed fermented dairy product containing Bifidobacterium lactis CNCM I-2494 suppresses gut hypersensitivity and colonic barrier disruption induced by acute stress in rats. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2012, 24, 376-e172. [Google Scholar] [CrossRef]
- Wu, X.; Wang, S.; Lu, J.; Jing, Y.; Li, M.; Cao, J.; Bian, B.; Hu, C. Seeing the unseen of Chinese herbal medicine processing (Paozhi): Advances in new perspectives. Chin. Med. 2018, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, R.L.; Zhang, Q.; Liu, J.; He, L.Y.; Huang, Q.W.; Peng, W.; Wu, C.J. Processing methods and mechanisms for alkaloid-rich Chinese herbal medicines: A review. J. Integr. Med. 2021, 19, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xu, J.D.; Zhou, S.S.; Shen, H.; Mao, Q.; Kong, M.; Zou, Y.T.; Xu, Y.Y.; Xu, J.; Li, S.L. Chemomics-based marker compounds mining and mimetic processing for exploring chemical mechanisms in traditional processing of herbal medicines, a continuous study on Rehmanniae Radix. J. Chromatogr. A 2017, 1530, 232–240. [Google Scholar] [CrossRef]
- Li, Z.; Li, J.; Zhang, F.; Zhu, N.; Sha, Z.; Li, D.; Tu, Y.; Hou, J. Antidiarrheal Effect of Sechang-Zhixie-San on Acute Diarrhea Mice and Network Pharmacology Deciphering Its Characteristics and Potential Mechanisms. Evid.-Based Complement. Altern. Med. Ecam 2020, 2020, 8880298. [Google Scholar] [CrossRef]
- Mo, L.; Zeng, Z.; Li, Y.; Li, D.; Yan, C.Y.; Xiao, S.; Huang, Y.H. Animal study of the anti-diarrhea effect and microbial diversity of dark tea produced by the Yao population of Guangxi. Food Funct. 2019, 10, 1999–2009. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, M.; Dou, D.; Kang, T.; Li, F. Effects of Deoxyschisandrin on Visceral Sensitivity of Mice with Inflammatory Bowel Disease. Evid.-Based Complement. Altern. Med. Ecam 2019, 2019, 2986097. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Jin, W.; Peng, Y.; He, Z.; Wei, L.; Li, S.; Wang, X.; Chang, M.; Wang, R. In vivo and vitro characterization of the effects of kisspeptin-13, endogenous ligands for GPR54, on mouse gastrointestinal motility. Eur. J. Pharmacol. 2017, 794, 216–223. [Google Scholar] [CrossRef]
- Wang, D.; Gao, N.; Zhou, T.; Zhang, Q.; Wang, J.; Li, A. Effect of Neuroligin1 and Neurexin1 on the Colonic Motility in a Mouse Model of Neuronal Intestinal Dysplasia. Gastroenterol. Res. Pract. 2020, 2020, 9818652. [Google Scholar] [CrossRef]
- Hayeeawaema, F.; Wichienchot, S.; Khuituan, P. Amelioration of gut dysbiosis and gastrointestinal motility by konjac oligo-glucomannan on loperamide-induced constipation in mice. Nutr. (Burbank Los Angeles Cty. Calif.) 2020, 73, 110715. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Mao, D.; Li, X.; Ma, Y.; Luan, Y.; Cao, Y.; Luan, Y. Changes of intestinal microflora diversity in diarrhea model of KM mice and effects of Psidium guajava L. as the treatment agent for diarrhea. J. Infect. Public Health 2020, 13, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Szymaszkiewicz, A.; Włodarczyk, J.; Mazur, M.; Olczak, J.; Fichna, J.; Zielińska, M. Cyclic derivatives of morphiceptin possess anti-transit effect in the gastrointestinal tract and alleviate abdominal pain in mice. Pharmacol. Rep. PR 2020, 72, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Barbara, G.; Stanghellini, V.; Brandi, G.; Cremon, C.; Di Nardo, G.; De Giorgio, R.; Corinaldesi, R. Interactions between commensal bacteria and gut sensorimotor function in health and disease. Am. J. Gastroenterol. 2005, 100, 2560–2568. [Google Scholar] [CrossRef] [PubMed]
- Lyte, M. Microbial endocrinology and infectious disease in the 21st century. Trends Microbiol. 2004, 12, 14–20. [Google Scholar] [CrossRef]
- Uribe, A.; Alam, M.; Johansson, O.; Midtvedt, T.; Theodorsson, E. Microflora modulates endocrine cells in the gastrointestinal mucosa of the rat. Gastroenterology 1994, 107, 1259–1269. [Google Scholar] [CrossRef]
- Spiller, R. Recent advances in understanding the role of serotonin in gastrointestinal motility in functional bowel disorders: Alterations in 5-HT signalling and metabolism in human disease. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2007, 19 (Suppl. 2), 25–31. [Google Scholar] [CrossRef]
- Gershon, M.D.; Liu, M.T. Serotonin and neuroprotection in functional bowel disorders. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2007, 19 (Suppl. 2), 19–24. [Google Scholar] [CrossRef] [Green Version]
- Gershon, M.D.; Tack, J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef]
- Stasi, C.; Bellini, M.; Bassotti, G.; Blandizzi, C.; Milani, S. Serotonin receptors and their role in the pathophysiology and therapy of irritable bowel syndrome. Tech. Coloproctol. 2014, 18, 613–621. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Arsenault, D. Gain modulation by serotonin in pyramidal neurones of the rat prefrontal cortex. J. Physiol. 2005, 566, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, M.; Lackner, J.M. Serotonin receptor modulators in the treatment of irritable bowel syndrome. Ther. Clin. Risk Manag. 2008, 4, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gershon, M.D. Review article: Roles played by 5-hydroxytryptamine in the physiology of the bowel. Aliment. Pharmacol. Ther. 1999, 13 (Suppl. 2), 15–30. [Google Scholar] [CrossRef]
- Chumpitazi, B.P.; Shulman, R.J. Underlying molecular and cellular mechanisms in childhood irritable bowel syndrome. Mol. Cell. Pediatr. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwee, K.A.; Lu, C.L.; Ghoshal, U.C. Epidemiology of irritable bowel syndrome in Asia: Something old, something new, something borrowed. J. Gastroenterol. Hepatol. 2009, 24, 1601–1607. [Google Scholar] [CrossRef]
- Camilleri, M. Physiological underpinnings of irritable bowel syndrome: Neurohormonal mechanisms. J. Physiol. 2014, 592, 2967–2980. [Google Scholar] [CrossRef]
- Furness, J.B.; Kunze, W.A.; Clerc, N. Nutrient tasting and signaling mechanisms in the gut. II. The intestine as a sensory organ: Neural, endocrine, and immune responses. Am. J. Physiol. 1999, 277, G922–G928. [Google Scholar] [CrossRef]
- Surawicz, C.M. Mechanisms of diarrhea. Curr. Gastroenterol. Rep. 2010, 12, 236–241. [Google Scholar] [CrossRef]
- Michelangeli, F.; Ruiz, M.C. I, 2. Physiology and pathophysiology of the gut in relation to viral diarrhea. Perspect. Med. Virol. 2003, 9, 23–50. [Google Scholar] [CrossRef]
- Guilarte, M.; Santos, J.; de Torres, I.; Alonso, C.; Vicario, M.; Ramos, L.; Martínez, C.; Casellas, F.; Saperas, E.; Malagelada, J.R. Diarrhoea-predominant IBS patients show mast cell activation and hyperplasia in the jejunum. Gut 2007, 56, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, S.; Tatewaki, M.; Yamada, T.; Fujimiya, M.; Mantyh, C.; Voss, M.; Eubanks, S.; Harris, M.; Pappas, T.N.; Takahashi, T. Short-chain fatty acids stimulate colonic transit via intraluminal 5-HT release in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R1269–R1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, Y.; Wei, K.; He, J.; Ding, N.; Hua, J.; Zhou, T.; Niu, F.; Zhou, G.; Shi, T.; et al. Review: Effect of Gut Microbiota and Its Metabolite SCFAs on Radiation-Induced Intestinal Injury. Front. Cell. Infect. Microbiol. 2021, 11, 577236. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cao, S.; Zhang, X. Modulation of Gut Microbiota-Brain Axis by Probiotics, Prebiotics, and Diet. J. Agric. Food Chem. 2015, 63, 7885–7895. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.J.; Chen, S.J.; Yu, Y.C.; Si, J.M.; Liu, B. A probiotic treatment containing Lactobacillus, Bifidobacterium and Enterococcus improves IBS symptoms in an open label trial. J. Zhejiang Univ. Sci. B 2006, 7, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Sinn, D.H.; Song, J.H.; Kim, H.J.; Lee, J.H.; Son, H.J.; Chang, D.K.; Kim, Y.H.; Kim, J.J.; Rhee, J.C.; Rhee, P.L. Therapeutic effect of Lactobacillus acidophilus-SDC 2012, 2013 in patients with irritable bowel syndrome. Dig. Dis. Sci. 2008, 53, 2714–2718. [Google Scholar] [CrossRef]
- Derrien, M.; Collado, M.C.; Ben-Amor, K.; Salminen, S.; de Vos, W.M. The Mucin degrader Akkermansia muciniphila is an abundant resident of the human intestinal tract. Appl. Environ. Microbiol. 2008, 74, 1646–1648. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [Green Version]
- Visca, P.; Seifert, H.; Towner, K.J. Acinetobacter infection--an emerging threat to human health. IUBMB Life 2011, 63, 1048–1054. [Google Scholar] [CrossRef]
- Smith, B.J.; Miller, R.A.; Ericsson, A.C.; Harrison, D.C.; Strong, R.; Schmidt, T.M. Changes in the gut microbiome and fermentation products concurrent with enhanced longevity in acarbose-treated mice. BMC Microbiol. 2019, 19, 130. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, J.G. Clinical practice. Antibiotic-associated diarrhea. N. Engl. J. Med. 2002, 346, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Méndez-García, C.; Rojo, D.; Barbas, C.; Moya, A. Antibiotic use and microbiome function. Biochem. Pharmacol. 2017, 134, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.A.; Konnikova, L.; Gerber, J.S. Impact of Antibiotics on Necrotizing Enterocolitis and Antibiotic-Associated Diarrhea. Gastroenterol. Clin. North Am. 2017, 46, 61–76. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M.; Sellin, J.H.; Barrett, K.E. Pathophysiology, Evaluation, and Management of Chronic Watery Diarrhea. Gastroenterology 2017, 152, 515–532.e512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (from 5’ End to 3’ End) | Product Size (bp) |
|---|---|---|
| 5-HT2AR-F | ACCGCTTTGGCAGTTTT | 140 |
| 5-HT2AR-R | GCGTTGAGGTGGCTTATT | |
| 5-HT3AR-F | GCAACCCCAGTCTCTTTGT | 143 |
| 5-HT3AR-R | GCTTGACGCCCTGATAAGT | |
| 5-HT4R-F | CATGCCCAGCAGATACAG | 147 |
| 5-HT4R-R | GAAACAGAAGCAGCCCAT | |
| SERT-F | CTCCTCCCCTCTAAGCCA | 185 |
| SERT-R | CCTCCTTCCTCTCCTCACA | |
| GAPDH-F | GATGGACACATTGGGGTT | 148 |
| GAPDH-R | AAAGCTGTGGCGTGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, S.; Ling, X.; Rui, W.; Jin, X.; Chu, F. LMWP (S3-3) from the Larvae of Musca domestica Alleviate D-IBS by Adjusting the Gut Microbiota. Molecules 2022, 27, 4517. https://doi.org/10.3390/molecules27144517
Peng S, Ling X, Rui W, Jin X, Chu F. LMWP (S3-3) from the Larvae of Musca domestica Alleviate D-IBS by Adjusting the Gut Microbiota. Molecules. 2022; 27(14):4517. https://doi.org/10.3390/molecules27144517
Chicago/Turabian StylePeng, Siyuan, Xiwen Ling, Wenjing Rui, Xiaobao Jin, and Fujiang Chu. 2022. "LMWP (S3-3) from the Larvae of Musca domestica Alleviate D-IBS by Adjusting the Gut Microbiota" Molecules 27, no. 14: 4517. https://doi.org/10.3390/molecules27144517
APA StylePeng, S., Ling, X., Rui, W., Jin, X., & Chu, F. (2022). LMWP (S3-3) from the Larvae of Musca domestica Alleviate D-IBS by Adjusting the Gut Microbiota. Molecules, 27(14), 4517. https://doi.org/10.3390/molecules27144517