1. Introduction
Coix lacryma-jobi L. var.
ma-yuen Gramineae, widely planted in Taiwan, China, and Japan, is commonly called coicis (or coix), adlay, adlay millet, Job’s tears (USA, UK), Huanren, or Pearl barley (China). For many decades, it has served as a healthy food supplement in many Asian countries. Among all cultivars,
Coix lacryma-jobi var.
ma-yuen Stapf is mostly used, due to its soft shell [
1]. Coicis seeds (
Figure S1 in Supplementary Materials) contain abundant lipids, polysaccharides, lignans, polyphenols, and adenosines [
2], and are widely utilized in traditional Chinese medicine for the treatment of various ailments, particularly cancer [
3]. To date, a number of its biological activities have attracted a great deal of attention, with activities including antioxidant/free radical scavenging [
4,
5,
6], anti-inflammatory [
7,
8], anti-mutagenic [
9], anti-tumor, anticancer [
10,
11,
12], hypolipidemics, hypocholesterolemics [
6,
13,
14,
15], hypoglycemics [
13,
16,
17], and anti-obesity [
18], etc. Coicis seeds consist of (g/100g) moisture 10.83, protein 13.05 (approximately double that of rice), fat 5.45, carbohydrate 68.60, fiber 0.36 and ash 1.3, and worth noting, coicis flour also contains 2.25% amylase [
19,
20].
Coicis seeds have also been reported to contain thiamine, riboflavin, niacin and ascorbic acid [
21], as well as a diversity of active components, including coniferyl alcohol, syringic acid, ferulic acid, syringaresinol, 4-ketopinoresinol, mayuenolide,
p-hydroxybenzaldehyde, vanillin, syringaldehyde, sinapaldehyde, coixol [
4,
22], trans-coniferylaldehyde and sinapaldehyde; coixenolide, caffeic acid, chlorogenic acid, coixspirolactams A-D, methyl dioxindole-3-acetate, ceramides, naringenin, gallic acid, and caffeic acid [
4,
23]. GC/MS analysis identified coix lactams and coixol to be present in the hull of coix seeds, and after polishing, the signals disappeared [
22].
Recently, ‘Pearl and Job’s Tears Powder’ has been commercialized as a health food with the indications ‘skin whitening and moisturizing’ and ‘spleen strengthening and wet-repelling’. Some products are applied as facial masks, such as ‘Pearl Barley & Milk’ and ‘Clear Turn Princess Veil, Rich Moist Mask’, and so on.
The safest and most effective way to treat cutaneous hyperpigmentation is to reduce melanin production by inhibiting tyrosinase activity [
24]. However, most tyrosinase inhibitors described in the literature lack clinical efficacy when incorporated into topical products [
24]. Hydroquinone (IC
50 > 4000 μM) and its derivative arbutin (IC
50 > 4000 μM) only weakly inhibited hTyr (human tyrosinase), and kojic acid showed a weak efficacy (IC
50 > 500 μM) [
24]. Thus, it is apparent that the facial mask market requires a potent and reliable anti hTyr preparation. To promote the market values of coicis, we explored the molecular action mechanism of the coicis active components regarding its anti-tyrosinase bioactivity. On the other hand, considering the cost efficiency and overall bioactivity beneficial to the skin physiology, we accessed the common extraction technology to reclaim the active constituents from coicis seeds and screened the best cultivar for processing the facial masks.
3. Materials and Methods
3.1. Source of Coix Seeds
Coicis seeds (Taichung No. 1~Taichung No. 5) (
Figure S4 in Supplementary Materials) were gifted by the local gross sales at Nan-Tou County, Taiwan. The seeds were harvested from the year 2019 to 2020 in batch wise collections. Each batch weighed 1 kg. A total of 8 batches were collected. The seeds were kept at 4–15 °C during transportation and rinsed quickly in laboratory with double distilled water twice for 2 min, immediately followed by tissue-wiping and blow drying. The desiccated fresh samples were stored at −20 °C until use.
3.2. Chemicals and Reagents
2,2-diphenyl-1-picrylhydrazyl (DPPH *) (0.5 mM in methanol), 2,2-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS
+), and ethylenediaminetetraacetic acid (EDTA) were provided by E. Merck (Dresden, Germany). Reference standard coixol (6-methoxy-2-benzoxazolinone; 6-MBOA) (purity 98.21%) was supplied by Med Chem Express (MCE, South Brunswick Township, NJ, USA). Coixenolide (CAS No. 29066-43-1) [(2S,3R)-3-[(Z)-hexadec-9-enoyl]oxybutan-2-yl] (
E)-octadec-11-enoate was purchased from Neostar United Industrial Co., Ltd. (Binjiang Economic Development Zone, Changzhou, Jiangsu 213033, China) (
www.neostarunited.com, accessed on 11 June 2022). The human tyrosinase fragment 369–377 was provided by Sigma Aldrich.
3.3. Sources of Cell Line
The B16F10 cells (ATCC CRL-6475, BCRC60031) were obtained from the Bioresource Collection and Research Center (BCRC, Hsinchu, Taiwan). The cells were maintained in DMEM (Hyclone, Logan, UT, USA) supplemented with 10% fetal bovine serum and 1% antibiotics at 37 °C, 5% CO2 in a humidified incubator.
3.4. Solvent Extractability
To 10 g of pulverized coicis seed powder, 200 mL of deionized water (or ethanol 95%, or methanol, or ethyl acetate) was added, and the mixture was maintained at 60 °C for 3 h with constant stirring. The extract was decanted to recover the solvent extract. The extractions were repeated for three times, the respective extracts were combined and subjected to rotary evaporation under reduced pressure until 100 mL to obtain the extracts of water or ethanol 95%, or methanol, or ethyl acetate, respectively.
3.5. Determination of Total Phenolic Acids and Flavonoids
The determination of the total phenolics and flavonoids were carried out by following the method as previously reported in our laboratory [
49].
3.6. DPPH Free Radical Scavenging Capability
The measurement of the DPPH radical scavenging activity was performed according to Brand-Williams et al. (1995) [
50]. Briefly, to 0.5 mL sample extracts, 0.3 mL DPPH radical reagent and 3 mL absolute ethanol were added. The optical density (changes in color from deep violet to light yellow) were monitored with a UV/VIS spectrophotometer (DU 800; Beckman Coulter, Fullerton, CA, USA) at 517 nm for 100 min against a blank containing a mixture of ethanol (3.5 mL) and DPPH radical reagent (0.3 mL). The scavenging activity percentage (
SA%) was calculated from Equation (1).
3.7. ABTS+ Antioxidant Capability
ABTS+ Antioxidant Assay Kit (Cat# AOX-1) (ZenBios, West Bengal, Indian) was used to evaluate the antioxidant capability of different extracts of coicis seeds by following the instructions given by the manufacturer.
3.8. Determination of the Phytosterol Content
The phytosterols were determined according to the method as previously reported [
49].
3.9. Extraction of Water-Soluble Polysaccharides
Chen et al. was followed with slight modification [
51]. In brief, the desiccated sample coix seeds were pulverized and defatted in a preparative Soxhlet apparatus (RBS company, Taoyuan, Taiwan). To the defatted residue, a 20-fold volume of deionized water was added, sonicated with 10 KW, refluxed at 95 °C for 4 h, and filtered. The extraction was repeated three times. The following step was carried out as directed. The purified polysaccharides were combined and lyophilized for use.
3.10. Isolation and Purification of Coixol
The protocol described by Li and Liu (2011) [
52] was followed to prepare high-purity coixol. The coixol obtained was then further purified in our laboratory. In brief, desiccated coix seeds were pulverized and sieved through #80 mesh. Then, 1000 g coix powder was percolated with 20 L sulfuric acid solution (1%). The following procedure was performed as cited. The coixol obtained was then further recrystallized in acetone–petroleum ether. Physicochemically, coixol is colorless and appears as needle like crystals (acetone–petroleum ether), m.p. 159–160 °C.
3.11. Isolation and Purification of Coixenolide
Ukita and Tanimura (1961) [
53] was followed with slight modification to carry out the isolation procedures. In brief, desiccated coix seeds were pulverized and sieved through #80 mesh. The powder (1000 g) was accurately weighed and subjected to preparative Soxhlet extraction with acetone (2.5 L) for 3 h. The syrup was dissolved in 250 mL petroleum ether, filtered, and the filtrate was concentrated in the rotary evaporator to yield reddish brown syrup. The syrup was dissolved in 50 mL petroleum ether, applied to a silica gel column (60 × 450 mm) and eluted with 800 mL petroleum ether. The eluate was evaporated in a rotary evaporator to give residue 1 (R1). R1 was redissolved in 50 mL petroleum ether, shaken twice with 40 mL 0.2 N KOH solution. The aqueous KOH layer was discarded. The alkali insoluble neutral substance (NS) was evaporated in a rotary evaporator to give residue RN1. The following procedures were carried out as directed using the alumina and silica gel (ASG) column for further separation. The chloroform eluate containing the NS was evaporated to give residue RN2. To advance the purity, 250 mg of the RN2 was subjected to silica gel column chromatographic separation (stationary phase: silica gel, column 10 mm × 350 mm), and sequentially eluted with petroleum ether containing ether (4%, 10%, and 50% ether in petroleum ether). The fractions eluted by 50:50 ether in petroleum were collected, combined, filtered, and evaporated under reduced pressure to obtain pure coixenolide (
1.4702–1.4705,
0
o). The purified coixenolide was used as the reference standard in the following assay for colixenolide content in coix seeds.
3.12. Determination of Coixol
The content of coixol in the extracts of coicis seeds was determined according to Ali et al. (2017) [
54]. In brief, the HPLC analyses of the standard coixol and coicis extracts were performed on Agilent 1200 SL Rapid Resolution HPLC-UV system (Agilent Technologies, Singapore). An amount of 2.0 μL of sample solution was injected. The mobile phase consisted of two solutions. Solution A (0.1% TFA in water,
v/v) and solution B (methanol) were operated in a linear gradient manner: 15% B from 0–1 min, 15–50% B from 1–2 min, 50–95% B from 2–6 min, maintained at 95% 6–7 min, and 95–15% B from 7–8 min. The column temperature was maintained at 30 °C [
54].
3.13. Assay of Coixenolide
The method of Yang et al. (2004) [
55] was employed with modification. Briefly, the desiccated pulverized (#80) coix seed powder was extracted in a preparative Soxhlet extractor with petroleum ether. Crude oil (1 g) obtained was dissolved in 20 mL of 7% (
w/w) methanolic HCl solution, subjected to acid-catalyzed transesterification by refluxing in a water bath at 100 °C for 4 h cooled and neutralized with 30% methanolic sodium methoxide (Sigma). The next steps were carried out as cited. The coixenolide content in the coix seeds was calculated from the calibration curve against 2,3-butanediol content obtained from different amount of coixenolide standards obtained in the above.
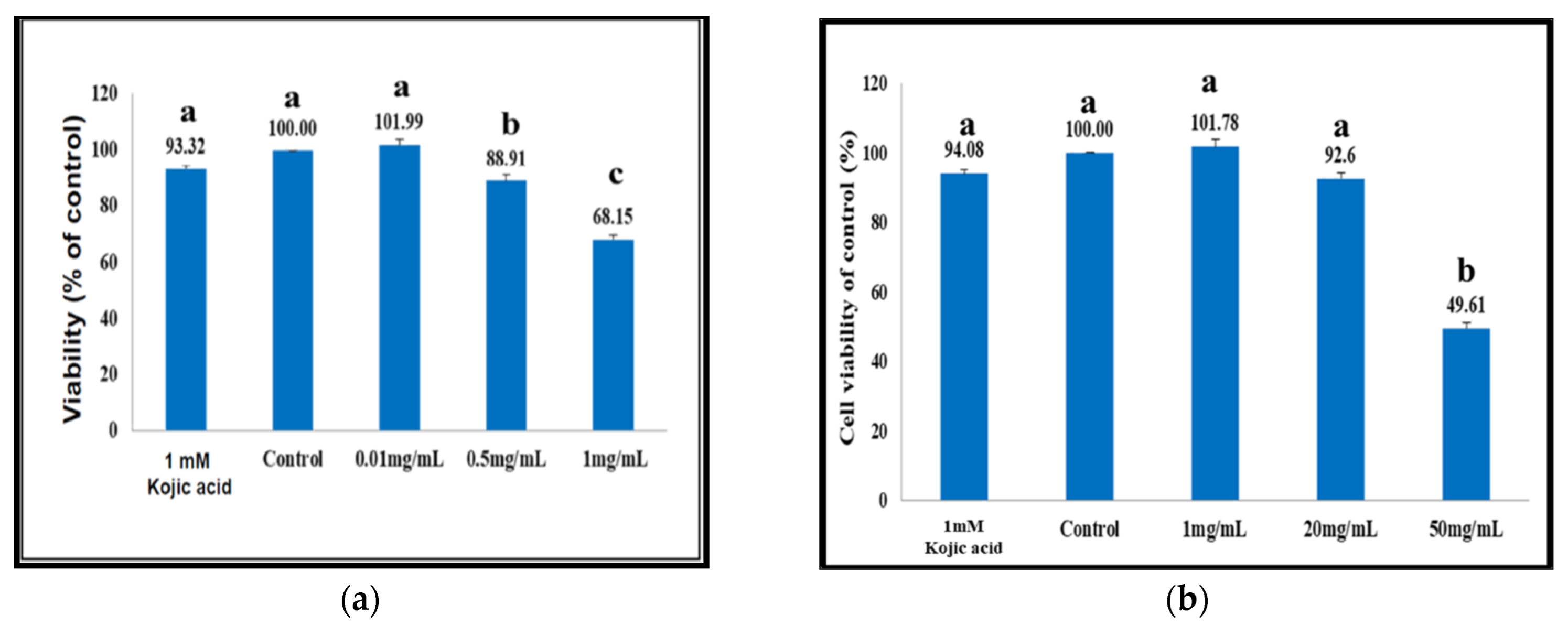
3.14. Cell Viability Affected by Different Coicis Seed Extracts
The B16F10 cells (1 × 105 cells/mL) were evenly pipetted into 10 culture tubes, 5 mL in each. In tubes 1–5, the aqueous extract was tested at 0 to 1.0 mg/mL; the ethanolic extract was tested in tubes 6–10 at 0 to 50 mg/mL. Kojic acid (1 mM) was used as the positive control. The tubes were further incubated in a humidified incubator at 37 °C under an atmosphere of 5% CO2 for 24 h. The absorbance of the broth was read at 600 nm using a microplate reader Gen5™ (BIO-TEK Instrument, Vermont, VT, USA). The percent viability was calculated taking the absorbance of the control as 100%.
3.15. Assay for the Intracellular Tyrosinase Activity
According to Yang et al. (2006) [
56], the cells obtained from the above were disrupted by ultrasonication, centrifuged at 3000×
g to discard the cell debris. The supernatant cell extract (100 μL) was mixed with freshly prepared L-DOPA solution (0.1% in phosphate-buffered saline) and incubated at 37 °C. The absorbance of this solution at 490 nm was measured with a microplate reader Gen5™ (BIO-TEK Instrument, Vermont, VT, USA) to estimate the production of dopachrome. Corrections were made for auto-oxidation of L-DOPA.
3.16. Assay for the Anti-Human Tyrosinase Fragment 369–377 Activity
The isolated coixol 100 mg was dissolved in 100 mL 95% ethanol to serve the stock solution (1 mg/mL). The inhibition assay for human tyrosinase fragment 369–377 (hTyr 369–377) was conducted as follows. In brief, different amount of coix seed extracts (aqueous extract 0.01, 0.50, and 1.00 mg/mL; ethanolic extracts 1.0, 20.0, 50.0 mg/mL), coixol (5, 10, 15 μg/mL), and 5 mM DOPA (in 50 mM sodium phosphate buffer, pH 6.8) were transferred into 96-well microtiter plate. To each well, hTyr 369–377 (200 units in 10 μL) was added. The reaction mixture was incubated at 37 °C for 30 min. The amount of dopachrome produced in the reaction mixture was determined spectrophotometrically at 490 nm (OD
490) using the ELISA microplate reader. Experiments were performed triplicate, and the inhibition percentage was evaluated using Equation (2):
where
B is the OD
490 values of the blank control, and
A is the OD
490 values for the coix seed extract treated group. Corrections were made for auto-oxidation of L-DOPA.
3.17. Preparation of the Facial Masks
The recipes for preparation of the coicis seed essence consisted of hyaluronic acid 3%, corn polyol 3%, deep moisturer 3%, deionized water 70.3%, antioxidant 0.1%, hydrophilic antibacterial agent 0.6%, and coicis seed extract 20%. In brief, the deionized water was heated to 50 °C, to which the antioxidant was added in portions. The mixture was cooled to ambient temperature and all other ingredients were added, mixed well to obtain the coicis seed essence. For preparation of the masks, the mask films, made of either bi-axially oriented polypropylene (BOPP) or cast polypropylene (CPP), were dipped into the coicis essence for 20 min to fully soak up the essence. The face masks were then removed and sealed in the packaging vinyl bags.
3.18. Test for Moisture Holding Capability
Skin moisture analyzer (Amazon, Co., Marston Gate, UK) was used to measure the percent skin moisture (IRB No: HK-HSSI-005 approved by HungKuang University, Taichung, Tiwan).
3.19. Statistical Analysis
The experiment was performed in triplicate for each substance. The results were expressed in percentage with respect to control values and compared by one-way ANOVA and Tukey’s test. A difference was considered statistically significant when p ≤ 0.05.
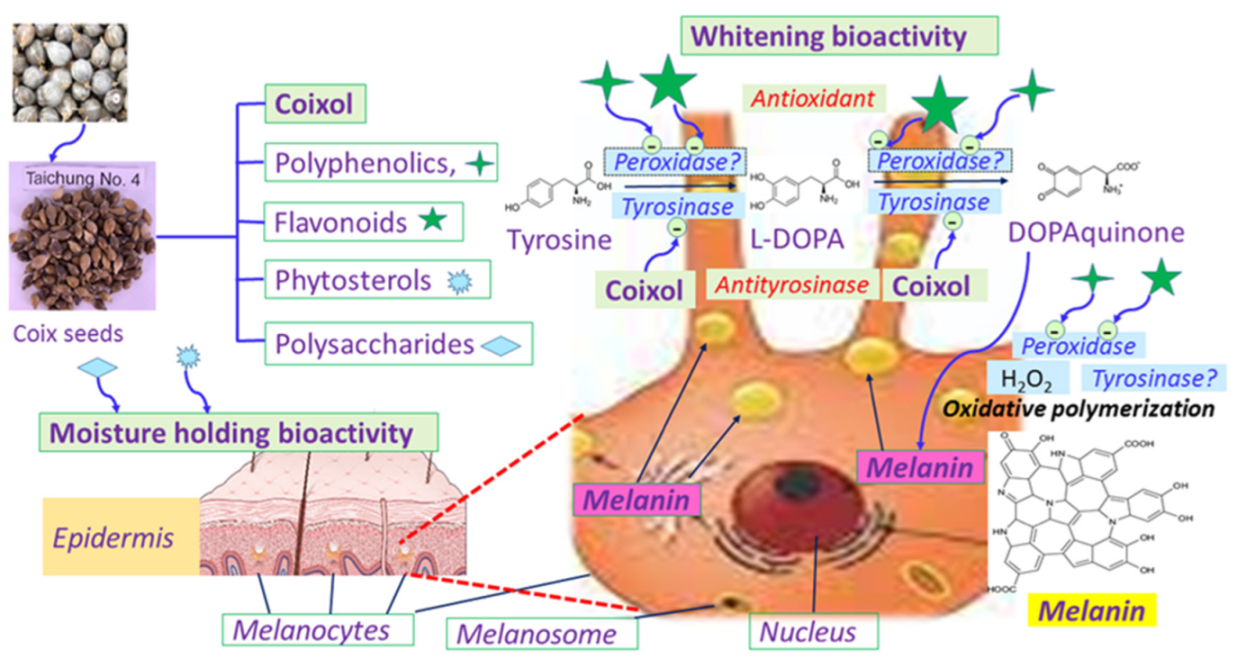
4. Conclusions
Facial skin is very delicate and vulnerable to environmental damage, including from chemical, physical, mechanical, and biological agents. Chemical damage may originate from UV screeners, whiting agents, moisturizers, emulsifies and face cleansing agents. Physical damage may involve radiation, thermal injuries, and improper sonication. Mechanical damage can be caused by underqualified massagers. In addition, biological damage could be induced by microbial infections coming from unsanitary or expired cosmetics. The development of facial masks from natural sources could avoid most of the above-mentioned sources of damage. Coicis seeds bear several nutraceutical and biomedical advantages, involving the promising anti-hTyr components coixol and others, as well as high contents of antioxidant and antimicrobial polyphenolics, flavonoids, and terpenes, high moisture holding and anti-inflammatory polysaccharides, phytosterol and ceramics. By carefully integrating the formulation and processing, coicis seeds are extremely feasible materials for facial mask fabrication.





{kind=link}
{kind=link}