
Pectinases Secretion by Saccharomyces cerevisiae: Optimization in Solid-State Fermentation and Identification by a Shotgun Proteomics Approach

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Microorganism
2.2. Sequential Strategy of Experimental Designs for the Production of Pectinases by SSF
2.2.1. Culture Medium and Inoculum
2.2.2. Experimental Designs
2.2.3. Crude Enzymatic Extract
2.2.4. Enzyme Assays
2.2.5. Effect of pH and Temperature on the Crude Extract Activity and Stability
2.3. Proteomic Analysis
2.4. Protein Digestion
2.5. Mass Spectrometry: Nano LC-MS/MS
2.6. Data Analysis
3. Results
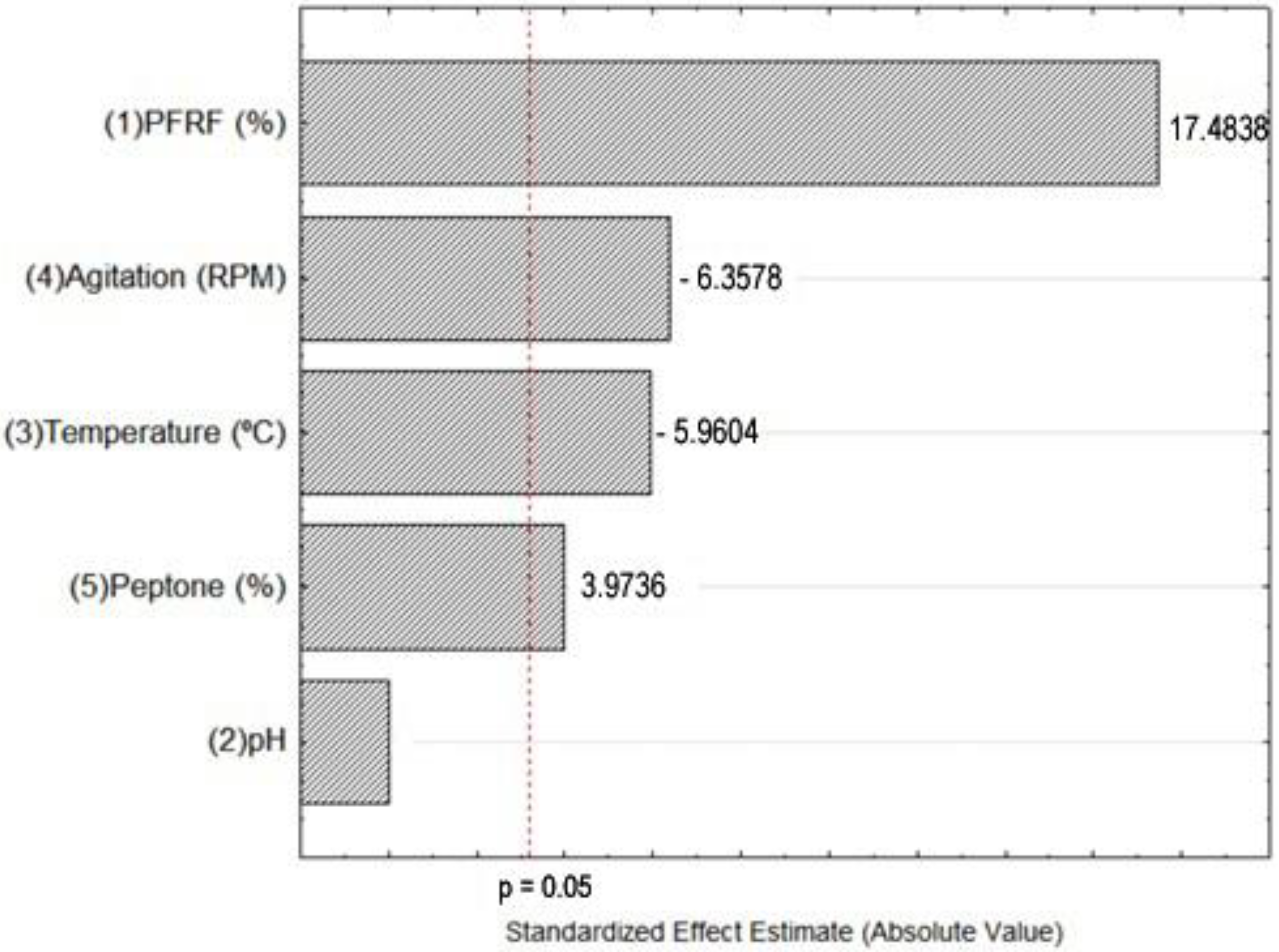
3.1. Fractional Factorial Experimental Design
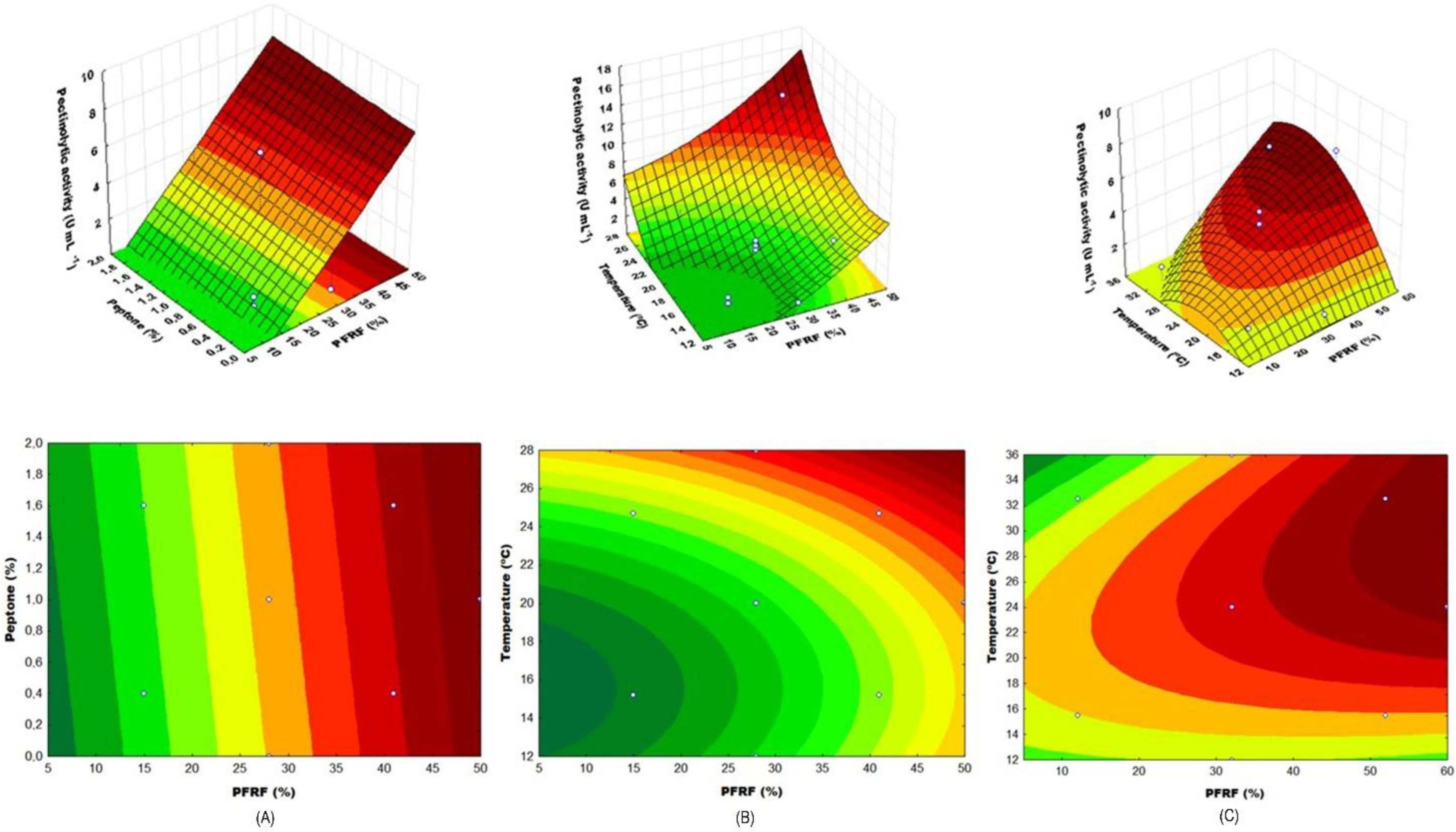
3.2. Central Composite Rotational Design 1 (CCRD-1)
3.3. Central Composite Rotational Design 2 (CCRD-2)
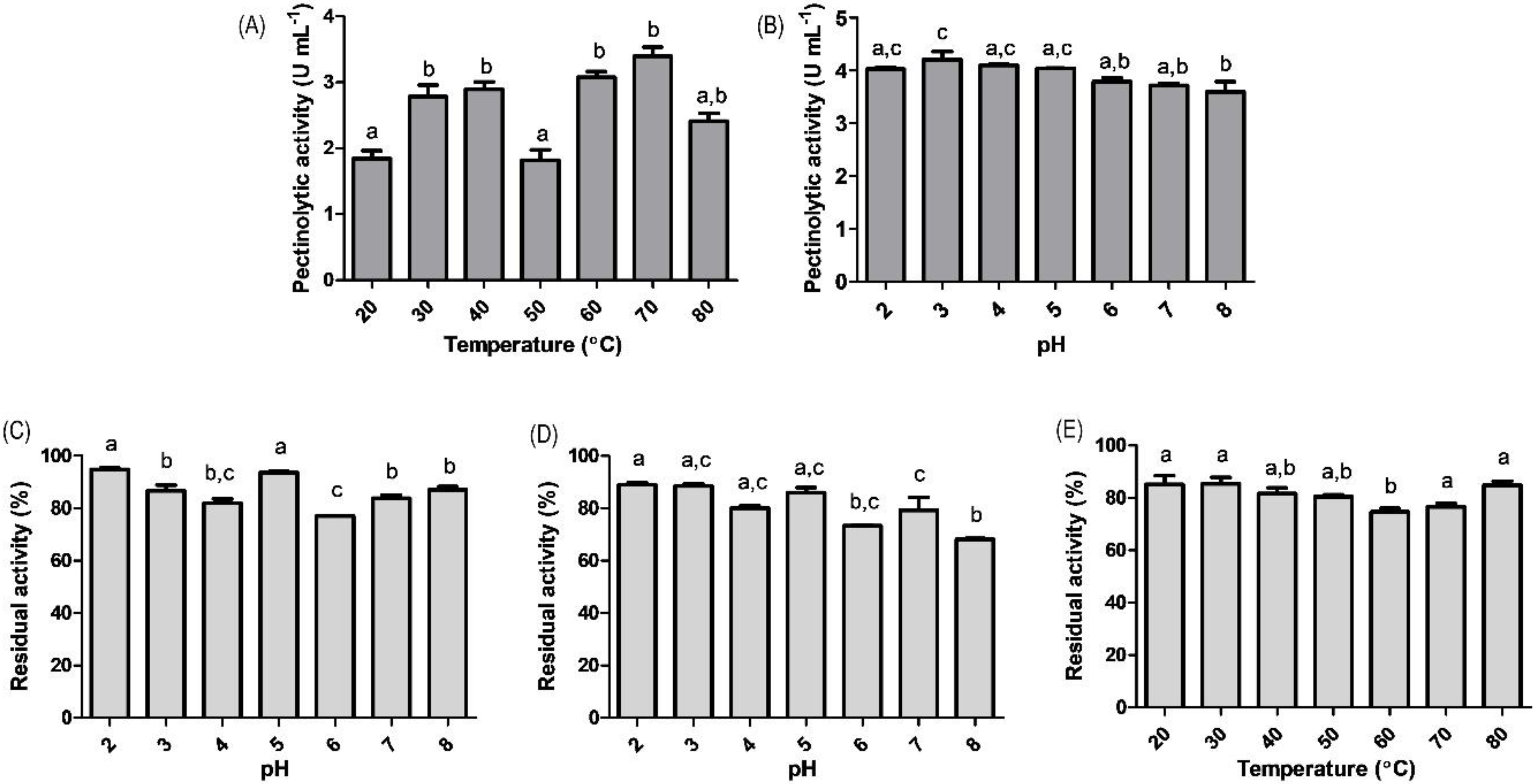
3.4. Effect of pH and Temperature on the Enzymatic Crude Extract Activity and Stability
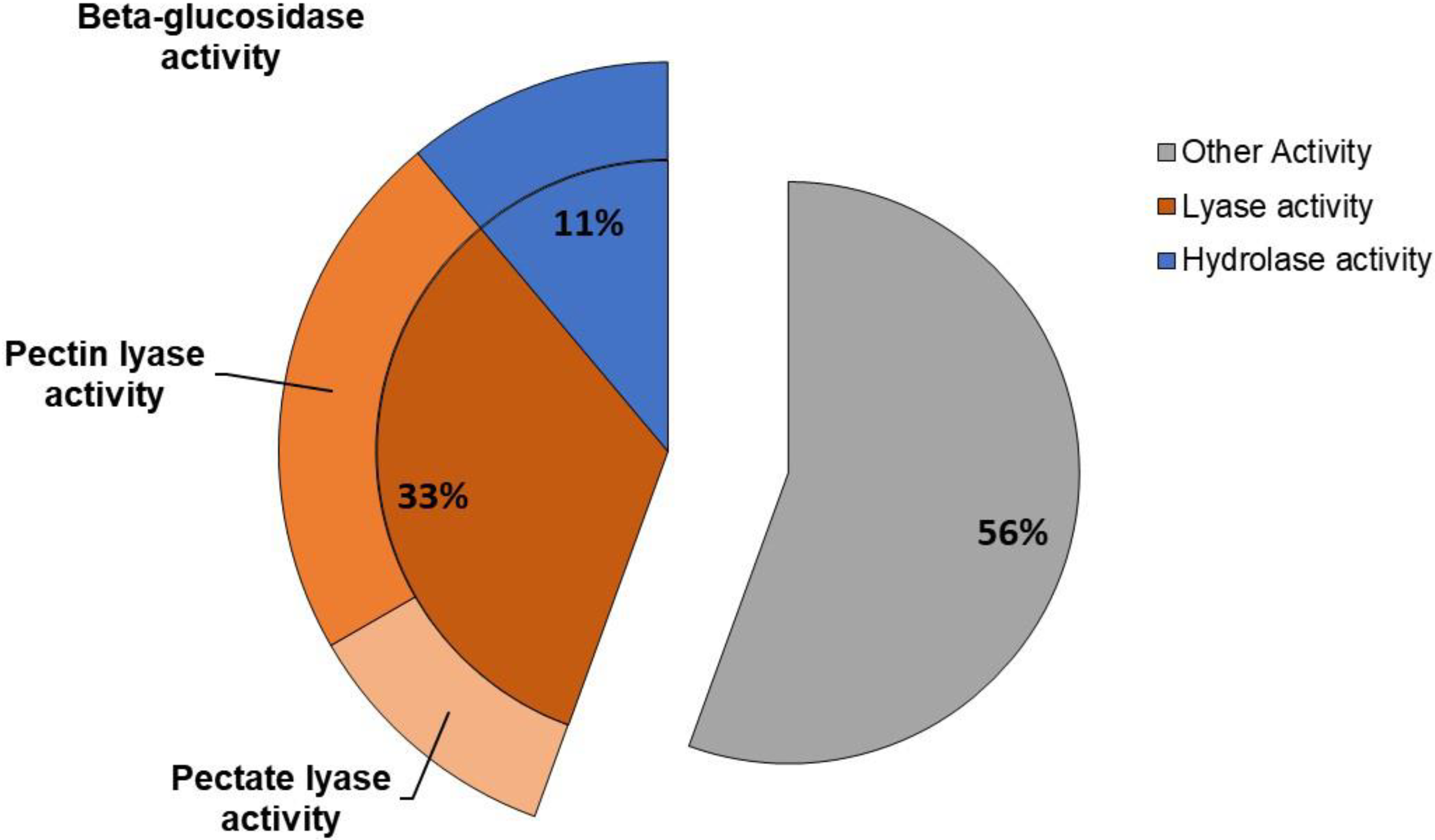
3.5. Pectinolytic Enzymes Activity
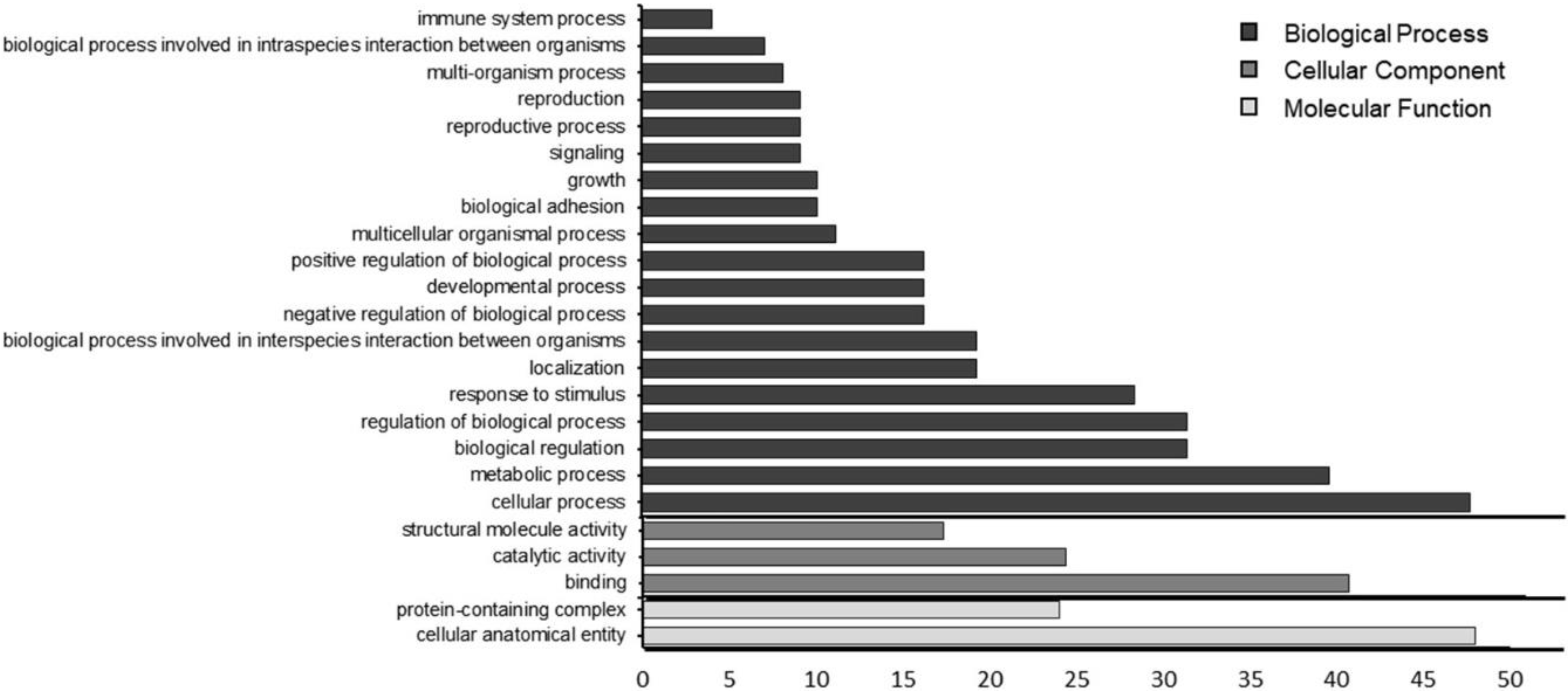
3.6. Proteomic Characterization
4. Discussion
4.1. Sequential Strategy of Experimental Designs for the Production of Pectinases by SSF
4.2. Fractional Factorial Experimental Design
4.3. Central Composite Rotational Design 1 (CCRD-1)
4.4. Central Composite Rotational Design 2 (CCRD-2)
4.5. Effect of pH and Temperature on the Enzymatic Crude Extract Activity and Stability
4.6. Pectinolytic Enzymes Activity
4.7. Proteomic Characterization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Nomenclature
References
- Amin, F.; Bhatti, H.N.; Bilal, M. Recent advances in the production strategies of microbial pectinases—A review. Int. J. Biol. Macromol. 2019, 122, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Oumer, O.J. Pectinase: Substrate, Production and their Biotechnological Applications. Int. J. Environ. Agric. Biotechnol. 2017, 2, 1007–1014. [Google Scholar] [CrossRef]
- Rebello, S.; Anju, M.; Aneesh, E.M.; Sindhu, R.; Binod, P.; Pandey, A. Recent advancements in the production and application of microbial pectinases: An overview. Rev. Environ. Sci. Biotechnol. 2017, 16, 381–394. [Google Scholar] [CrossRef]
- Samanta, S. Microbial Pectinases: A Review on Molecular and Biotechnological Perspectives. J. Microbiol. Biotechnol. Food Sci. 2019, 9, 248–266. [Google Scholar] [CrossRef]
- John, J.; Kaimal, K.K.S.; Smith, M.L.; Rahman, P.K.S.M.; Chellam, P.V. Advances in upstream and downstream strategies of pectinase bioprocessing: A review. Int. J. Biol. Macromol. 2020, 162, 1086–1099. [Google Scholar] [CrossRef] [PubMed]
- Maidana, S.A.; Butiuk, A.P.; Zubreski, E.R.; Hours, R.A.; Brumovsky, L.A.; Martos, M.A. Production of an endopolygalacturonase from Wickerhanomyces anomalus with disintegration activity on plant tissues. Biocatal. Agric. Biotechnol. 2019, 18, 101042. [Google Scholar] [CrossRef]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef]
- Naumova, E.S.; Borovkova, A.N.; Shalamitskiy, M.Y.; Naumov, G.I. Natural Polymorphism of Pectinase PGU Genes in the Saccharomyces Yeasts. Microbiology 2021, 90, 349–360. [Google Scholar] [CrossRef]
- Lian, J.; Mishra, S.; Zhao, H. Recent advances in metabolic engineering of Saccharomyces cerevisiae: New tools and their applications. Metab. Eng. 2018, 50, 85–108. [Google Scholar] [CrossRef]
- Melnichuk, N.; Braia, M.J.; Anselmi, P.A.; Meini, M.-R.; Romanini, D. Valorization of two agroindustrial wastes to produce alpha-amylase enzyme from Aspergillus oryzae by solid-state fermentation. Waste Manag. 2020, 106, 155–161. [Google Scholar] [CrossRef]
- Panesar, P.S.; Kaur, R.; Singla, G.; Sangwan, R.S. Bio-processing of agro-industrial wastes for production of food-grade enzymes: Progress and prospects. Appl. Food Biotechnol. 2016, 3, 208–227. [Google Scholar] [CrossRef]
- FAO. Food Outlook: Biannual Report on Global Food Markets 169; FAO: Rome, Italy, 2018. [Google Scholar]
- Duarte, Y.; Chaux, A.; Lopez, N.; Largo, E.; Ramírez, C.; Nuñez, H.; Simpson, R.; Vega, O. Effects of Blanching and Hot Air Drying Conditions on the Physicochemical and Technological Properties of Yellow Passion Fruit (Passiflora edulis Var. Flavicarpa) by-Products. J. Food Process Eng. 2017, 40, e12425. [Google Scholar] [CrossRef]
- Silva, N.C.; Santana, R.C.; Duarte, C.R.; Barrozo, M.A.S. Impact of freeze-drying on bioactive compounds of yellow passion fruit residues. J. Food Process Eng. 2017, 40, e12514. [Google Scholar] [CrossRef]
- Silva, N.C.; Duarte, C.R.; Barrozo, M.A.S. Effects of dehydration methods on quality characteristics of yellow passion fruit co-products. J. Sci. Food Agric. 2017, 97, 4750–4759. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Molecular relationships among hyphal ascomycetous yeasts and yeastlike taxa. Can. J. Bot. 1995, 73, 824–830. [Google Scholar] [CrossRef]
- De Carvalho, M.C.; Fai, A.E.C.; Koblitz, M.G.B.; Uetanabaro, A.P.T.; da Costa, A.M. Seleção de linhagens de Saccharomyces cerevisiae com maior potencial secretor de pectinases. In Anais do Simpósio Latino-Americano de Ciências de Alimentos, 2015, Anais eletrônicos. Campinas, Galoá. 2015. Available online: https://proceedings.science/slaca/slaca-2015/papers/selecao-de-linhagens-de-saccharomyces-cerevisiae-com-maior-potencial-secretor-de-pectinases (accessed on 12 October 2021).
- Barbosa, C.D.; Da Costa, E.C.; Costa, I.M.; Lacerda, I.C.A.; Lopes, E.D.S.O. Obtenção e caracterização de vinagre de manga pelo método de acetificação de Orleans. Res. Soc. Dev. 2020, 9, e127985593. [Google Scholar] [CrossRef]
- Bhattacharya, S. Chapter Central Composite Design for Response Surface Methodology and Its Application in Pharmacy. In Response Surface Methodology in Engineering Science; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar] [CrossRef]
- Uszkoreit, J.; Winkelhardt, D.; Barkovits, K.; Wulf, M.; Roocke, S.; Marcus, K.; Eisenacher, M. MaCPepDB: A Database to Quickly Access All Tryptic Peptides of the UniProtKB. J Proteome Res. 2021, 20, 2145–2150. [Google Scholar] [CrossRef]
- De Carvalho Silva, J.; de França, P.R.L.; de Melo, A.H.F.; Neves-Petersen, M.T.; Converti, A.; Souza Porto, T. Optimized production of Aspergillus aculeatus URM4953 polygalacturonases for pectin hydrolysis in hog plum (Spondias mombin L.) juice. Process Biochem. 2019, 79, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Uenojo, M.; Pastore, G.M. Isolamento e seleção de microrganismos pectinolíticos a partir de resíduos provenientes de agroindústrias para produção de aromas frutais. Food Sci. Technol. 2006, 26, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Poondla, V.; Bandikari, R.; Subramanyam, R.; Reddy Obulam, V.S. Low temperature active pectinases production by Saccharomyces cerevisiae isolate and their characterization. Biocatal. Agric. Biotechnol. 2015, 4, 70–76. [Google Scholar] [CrossRef]
- Heerd, D.; Yegin, S.; Tari, C.; Fernandez-Lahore, M. Pectinase enzyme-complex production by Aspergillus spp. in solid-state fermentation: A comparative study. Food Bioprod. Process. 2012, 90, 102–110. [Google Scholar] [CrossRef]
- Santos, R.M.; Nogueira, F.C.; Brasil, A.A.; Carvalho, P.C.; Leprevost, F.V.; Domont, G.B.; Eleutherio, E.C. Quantitative proteomic analysis of the Saccharomyces cerevisiae industrial strains CAT-1 and PE-2. J. Proteom. 2017, 151, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Gogus, N.; Evcan, E.; Tarı, C.; Cavalitto, S.F. Evaluation of agro-industrial wastes, their state, and mixing ratio for maximum polygalacturonase and biomass production in submerged fermentation. Environ. Technol. 2015, 36, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Paz, T.A.; Dos Santos, V.A.F.F.M.; Inácio, M.C.; Dias, N.B.; Palma, M.S.; Pereira, A.M.S.; Furlan, M. Proteome profiling reveals insights into secondary metabolism in Maytenus ilicifolia (Celastraceae) cell cultures producing quinonemethide triterpenes. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 130, 405–416. [Google Scholar] [CrossRef]
- Poondla, V.; Yannam, S.K.; Gummadi, S.N.; Subramanyam, R.; Reddy Obulam, V.S. Enhanced production of pectinase by Saccharomyces cerevisiae isolate using fruit and agro-industrial wastes: Its application in fruit and fiber processing. Biocatal. Agric. Biotechnol. 2016, 6, 40–50. [Google Scholar] [CrossRef]
- Paulo, J.A.; O’Connell, J.D.; Gaun, A.; Gygi, S.P. Proteome-wide quantitative multiplexed profiling of protein expression: Carbon-source dependency inSaccharomyces cerevisiae. Mol. Biol. Cell 2015, 26, 4063–4074. [Google Scholar] [CrossRef] [PubMed]
- McCotter, S.W.; Horianopoulos, L.C.; Kronstad, J.W. Regulation of the fungal secretome. Curr. Genet. 2016, 62, 533–545. [Google Scholar] [CrossRef]
- Biz, A.; Finkler, A.T.J.; Pitol, L.O.; Medina, B.S.; Krieger, N.; Mitchell, D.A. Production of pectinases by solid-state fermentation of a mixture of citrus waste and sugarcane bagasse in a pilot-scale packed-bed bioreactor. Biochem. Eng. J. 2016, 111, 54–62. [Google Scholar] [CrossRef]
- Martos, M.A.; Zubreski, E.R.; Garro, O.A.; Hours, R.A. Production of Pectinolytic Enzymes by the Yeast Wickerhanomyces anomalus Isolated from Citrus Fruits Peels. Biotechnol. Res. Int. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jayani, R.S.; Saxena, S.; Gupta, R. Microbial pectinolytic enzymes: A review. Process Biochem. 2005, 40, 2931–2944. [Google Scholar] [CrossRef]
- Zakhartsev, M.; Yang, X.; Reuss, M.; Pörtner, H.O. Metabolic efficiency in yeast Saccharomyces cerevisiae in relation to temperature dependent growth and biomass yield. J. Therm. Biol. 2015, 52, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Arévalo-Villena, M.; Fernández, M.; López, J.; Briones, A. Pectinases yeast production using grape skin as carbon source. Adv. Biosci. Biotechnol. 2011, 2, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Singh, A.; Dua, A.; Mahajan, R. Cost-effective and concurrent production of industrially valuable xylano-pectinolytic enzymes by a bacterial isolate Bacillus pumilus AJK. Prep. Biochem. Biotechnol. 2016, 47, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Rollero, S.; Roberts, S.; Bauer, F.F.; Divol, B. Agitation impacts fermentation performance as well as carbon and nitrogen metabolism in Saccharomyces cerevisiae under winemaking conditions. Aust. J. Grape Wine Res. 2018, 24, 360–367. [Google Scholar] [CrossRef]
- Jaramillo, P.M.D.; Andreaus, J.; Neto, G.P.D.S.; Castro, C.F.D.S.; Filho, E.X.F. The characterization of a pectin-degrading enzyme fromAspergillus oryzaegrown on passion fruit peel as the carbon source and the evaluation of its potential for industrial applications. Biocatal. Biotransformation 2015, 33, 310–322. [Google Scholar] [CrossRef]
- Mohandas, A.; Raveendran, S.; Parameswaran, B.; Abraham, A.; Athira, R.S.; Mathew, A.K.; Pandey, A. Production of Pectinase from Bacillus sonorensis MPTD1. Food Technol. Biotechnol. 2018, 56, 110–116. [Google Scholar] [CrossRef]
- Müller-Maatsch, J.; Bencivenni, M.; Caligiani, A.; Tedeschi, T.; Bruggeman, G.; Bosch, M.; Petrusan, J.; Van Droogenbroeck, B.; Elst, K.; Sforza, S. Pectin content and composition from different food waste streams. Food Chem. 2016, 201, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, G.; Ahmad, E.; Khan, M.V.; Ashraf, M.T.; Bhat, R.; Khan, R.H. Impact of structural stability of cold adapted Candida antarctica lipase B (CaLB): In relation to pH, chemical and thermal denaturation. RSC Adv. 2015, 5, 20115–20131. [Google Scholar] [CrossRef]
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef]
- Bennamoun, L.; Hiligsmann, S.; Dakhmouche, S.; Ait-Kaki, A.; Labbani, F.-Z.K.; Nouadri, T.; Meraihi, Z.; Turchetti, B.; Buzzini, P.; Thonart, P. Production and Properties of a Thermostable, pH—Stable Exo-Polygalacturonase Using Aureobasidium pullulans Isolated from Saharan Soil of Algeria Grown on Tomato Pomace. Foods 2016, 5, 72. [Google Scholar] [CrossRef] [Green Version]
- Arcus, V.L.; Prentice, E.J.; Hobbs, J.; Mulholland, A.; van der Kamp, M.; Pudney, C.; Parker, E.; Schipper, L.A. On the Temperature Dependence of Enzyme-Catalyzed Rates. Biochemistry 2016, 55, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q. On the indirect relationship between protein dynamics and enzyme activity. Prog. Biophys. Mol. Biol. 2017, 125, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Barnby, F.M.; Morpeth, F.F.; Pyle, D.L. Endopolygalacturonase production from Kluyveromyces marxianus. I. Resolution, purification, and partial characterisation of the enzyme. Enz Microb Tech. 1990, 11, 891–897. [Google Scholar] [CrossRef]
- Dubey, A.K.; Yadav, S.; Kumar, M.; Anand, G.; Yadav, D. Molecular Biology of Microbial Pectate Lyase: A Review. Br. Biotechnol. J. 2016, 13, 1–26. [Google Scholar] [CrossRef]
- Zhu, M.; He, H.; Fan, M.; Ma, H.; Ren, H.; Zeng, J.; Gao, H. Application and optimization of solid-state fermentation process for enhancing polygalacturonase production by Penicillium expansum. Int. J. Agric. Biol. Eng. 2018, 11, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Lin, J.; Wang, C.; Du, X.; Cai, J. Purification and characterization of exo-polygalacturonase from Zygoascus hellenicus V25 and its potential application in fruit juice clarification. Food Sci. Biotechnol. 2016, 25, 1379–1385. [Google Scholar] [CrossRef]
- Gainvors, A.; Frézier, V.; Lemaresquier, H.; Lequart, C.; Aigle, M.; Belarbi, A. Detection of polygalacturonase, pectin-lyase and pectin-esterase activities in aSaccharomyces cerevisiae strain. Yeast 1994, 10, 1311–1319. [Google Scholar] [CrossRef]
- Pili, J.; Danielli, A.; Nyari, N.L.D.; Zeni, J.; Cansian, R.L.; Backes, G.T.; Valduga, E. Biotechnological potential of agro-industrial waste in the synthesis of pectin lyase from Aspergillus brasiliensis. Food Sci. Technol. Int. 2018, 24, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Gummadi, S.N.; Kumar, D.S. Batch and fed batch production of pectin lyase and pectate lyase by novel strain Debaryomyces nepalensis in bioreactor. Bioresour Technol. 2008, 99, 874–881. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coded Variable (Level) | pH | Agitation (rpm) | Peptone (% w/w) | Temperature (°C) | PFRF (% w/w) | |
|---|---|---|---|---|---|---|
| Fractional factorial | −1 | 5 | 0 | 0.0 | 21.0 | 1.0 |
| 0 | 7 | 100 | 0.5 | 28.0 | 6.0 | |
| +1 | 9 | 200 | 1.0 | 35.0 | 11.0 | |
| CCRD 1 | −α (−1.68) | - | - | 0.0 | 12.0 | 4.0 |
| −1 | - | - | 0.4 | 15.2 | 15.0 | |
| 0 | - | - | 1.0 | 20.0 | 28.0 | |
| +1 | - | - | 1.6 | 24.7 | 41.0 | |
| +α (+1.68) | - | - | 2.0 | 28.0 | 50.0 | |
| CCRD 2 | −α (−1.41) | - | - | - | 12.0 | 4.0 |
| −1 | - | - | - | 15.5 | 12.0 | |
| 0 | - | - | - | 24.0 | 32.0 | |
| +1 | - | - | - | 32.5 | 52.0 | |
| +α (+1.41) | - | - | - | 52.0 | 60.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takeyama, M.M.; de Carvalho, M.C.; Carvalho, H.S.; Silva, C.R.; Uetanabaro, A.P.T.; da Costa, A.M.; Evaristo, J.A.M.; Nogueira, F.C.S.; Fai, A.E.C.; Koblitz, M.G.B. Pectinases Secretion by Saccharomyces cerevisiae: Optimization in Solid-State Fermentation and Identification by a Shotgun Proteomics Approach. Molecules 2022, 27, 4981. https://doi.org/10.3390/molecules27154981
Takeyama MM, de Carvalho MC, Carvalho HS, Silva CR, Uetanabaro APT, da Costa AM, Evaristo JAM, Nogueira FCS, Fai AEC, Koblitz MGB. Pectinases Secretion by Saccharomyces cerevisiae: Optimization in Solid-State Fermentation and Identification by a Shotgun Proteomics Approach. Molecules. 2022; 27(15):4981. https://doi.org/10.3390/molecules27154981
Chicago/Turabian StyleTakeyama, Matheus Mikio, Márcia Corrêa de Carvalho, Helena Sacco Carvalho, Cristiane Rodrigues Silva, Ana Paula Trovatti Uetanabaro, Andrea Miura da Costa, Joseph A. Medeiros Evaristo, Fábio César Sousa Nogueira, Ana Elizabeth Cavalcante Fai, and Maria Gabriela Bello Koblitz. 2022. "Pectinases Secretion by Saccharomyces cerevisiae: Optimization in Solid-State Fermentation and Identification by a Shotgun Proteomics Approach" Molecules 27, no. 15: 4981. https://doi.org/10.3390/molecules27154981
APA StyleTakeyama, M. M., de Carvalho, M. C., Carvalho, H. S., Silva, C. R., Uetanabaro, A. P. T., da Costa, A. M., Evaristo, J. A. M., Nogueira, F. C. S., Fai, A. E. C., & Koblitz, M. G. B. (2022). Pectinases Secretion by Saccharomyces cerevisiae: Optimization in Solid-State Fermentation and Identification by a Shotgun Proteomics Approach. Molecules, 27(15), 4981. https://doi.org/10.3390/molecules27154981