
Phenolic Profile, Antioxidant and Enzyme Inhibitory Activities of Leaves from Two Cassia and Two Senna Species


 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Polyphenolic and Flavonoid Contents
2.2. Chemical Phenolic Profile by HPLC–MS/MS
2.3. Antioxidant Activity
2.4. Enzyme Inhibition Activity
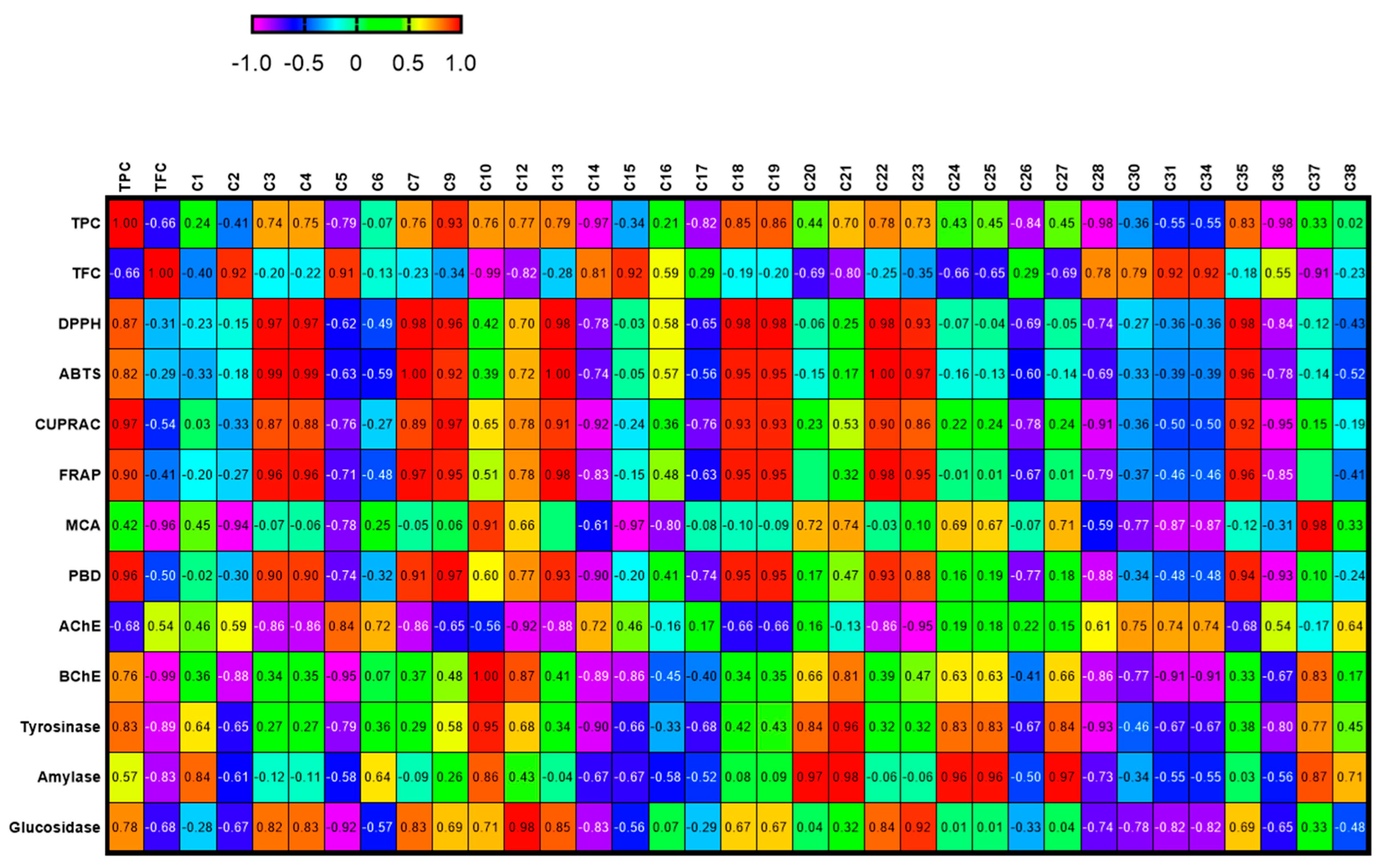
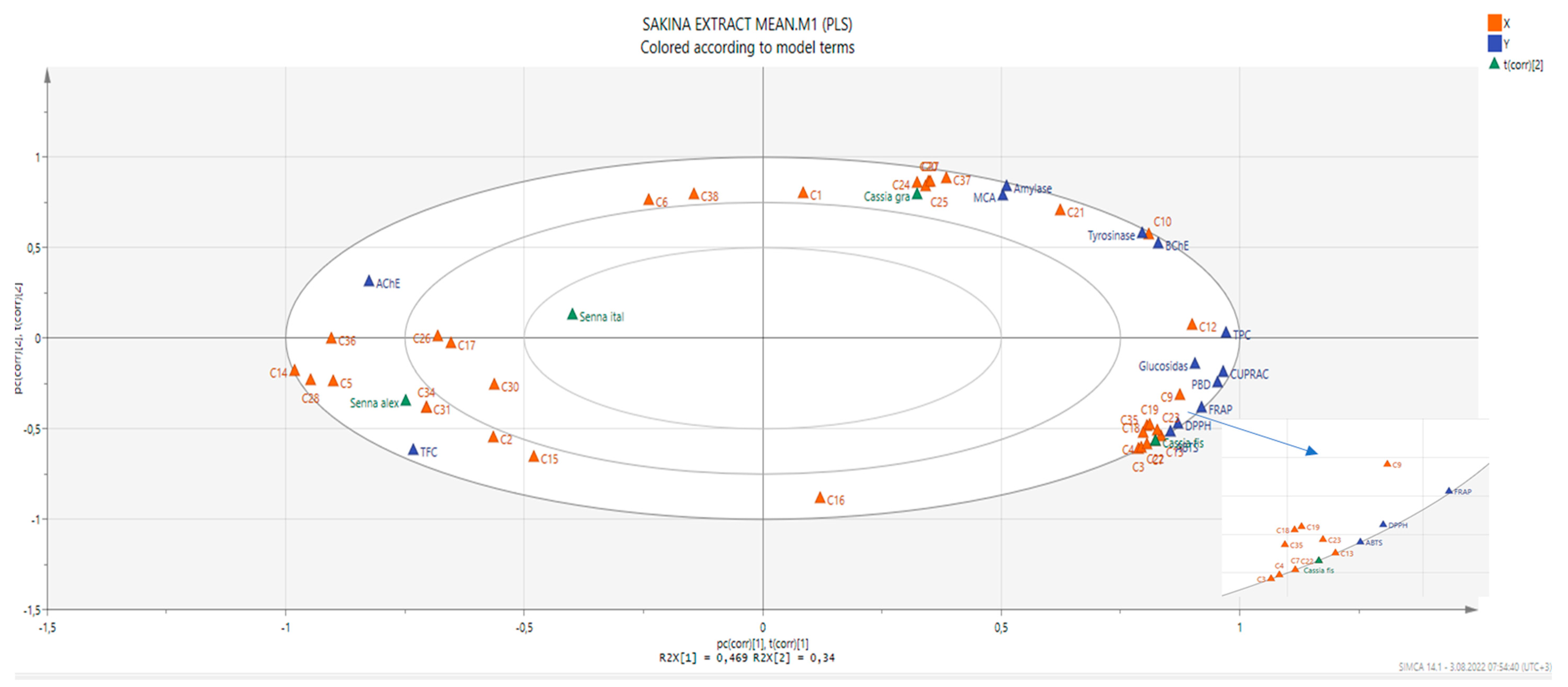
2.5. Data Mining
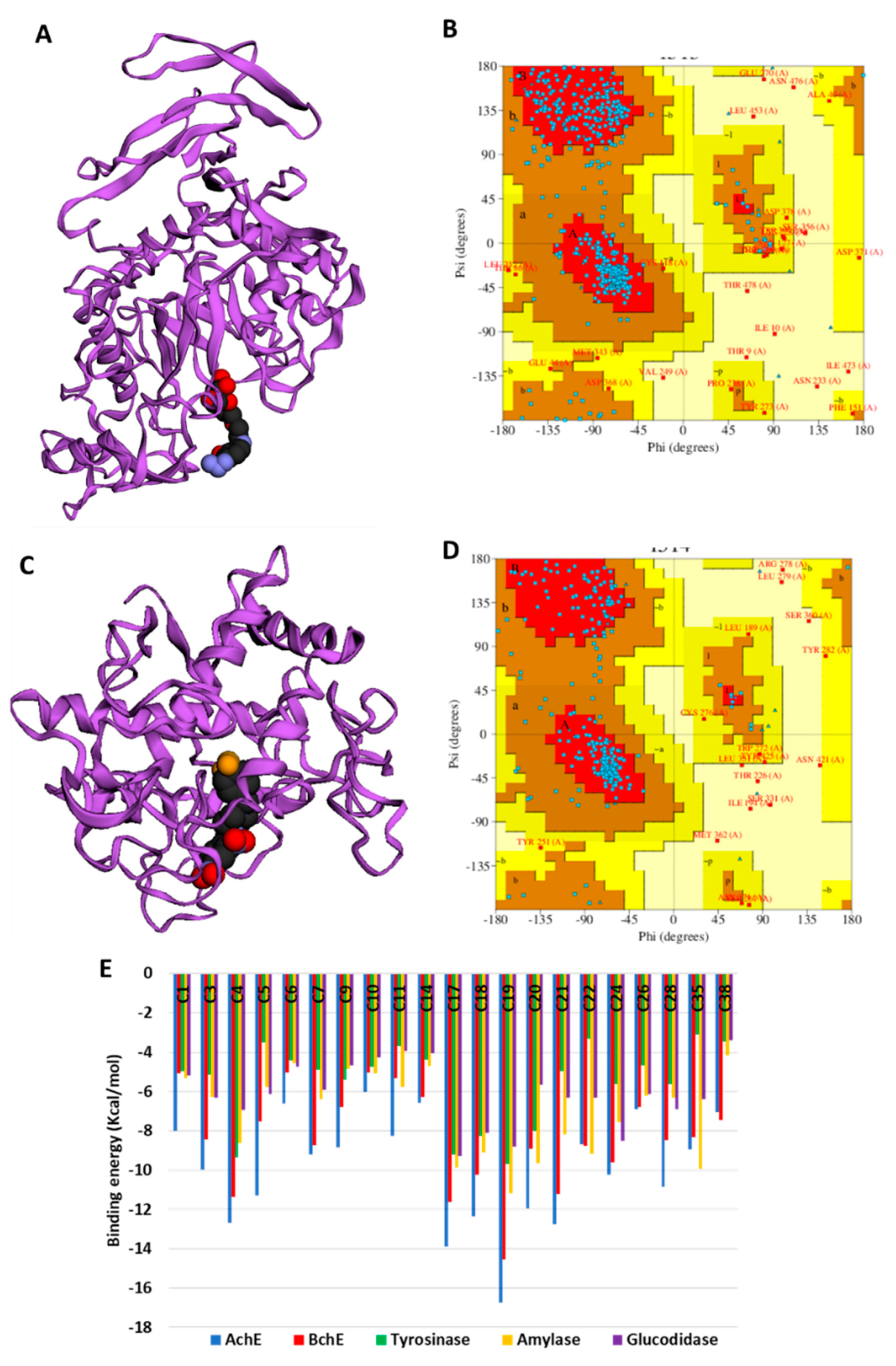
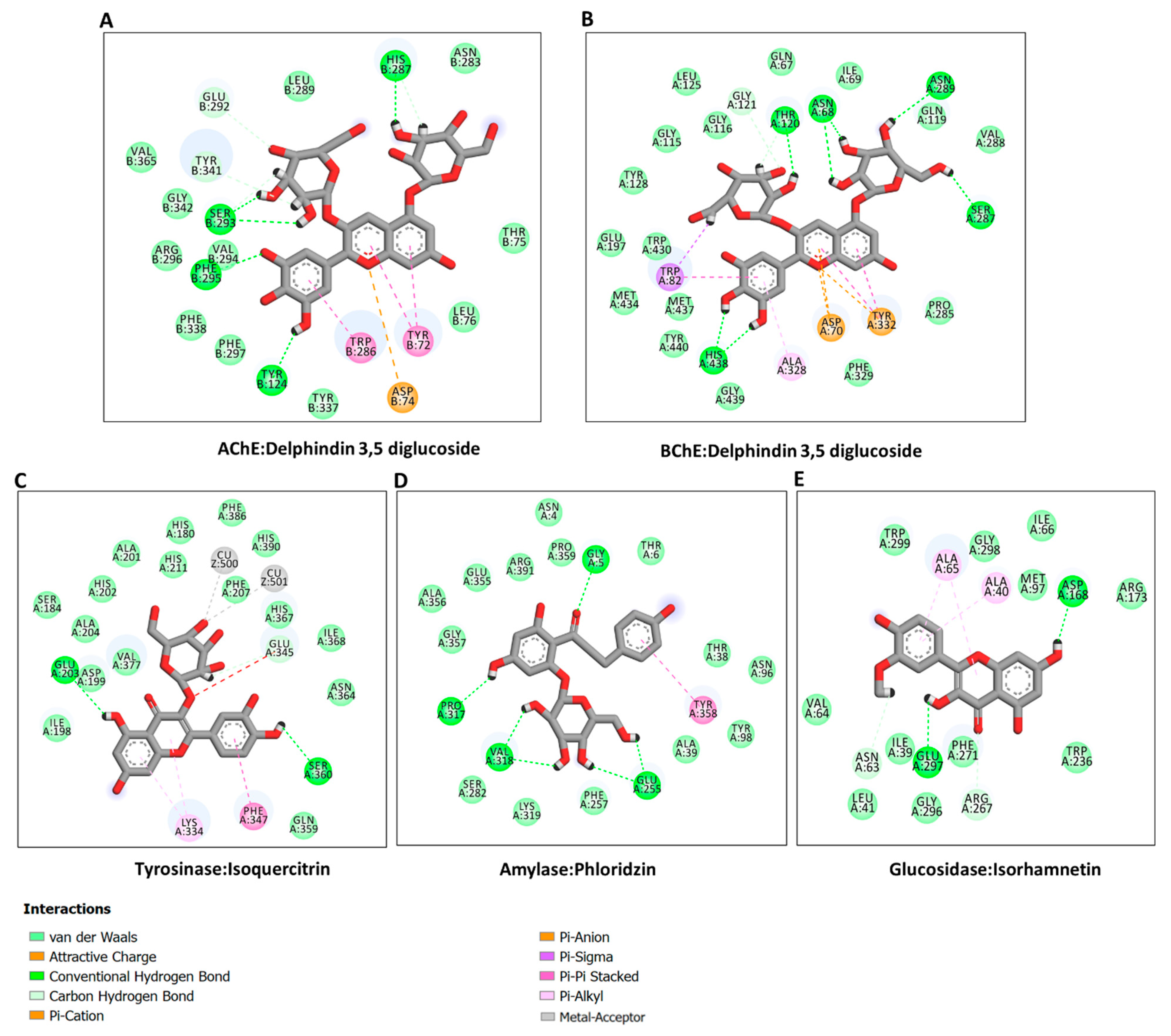
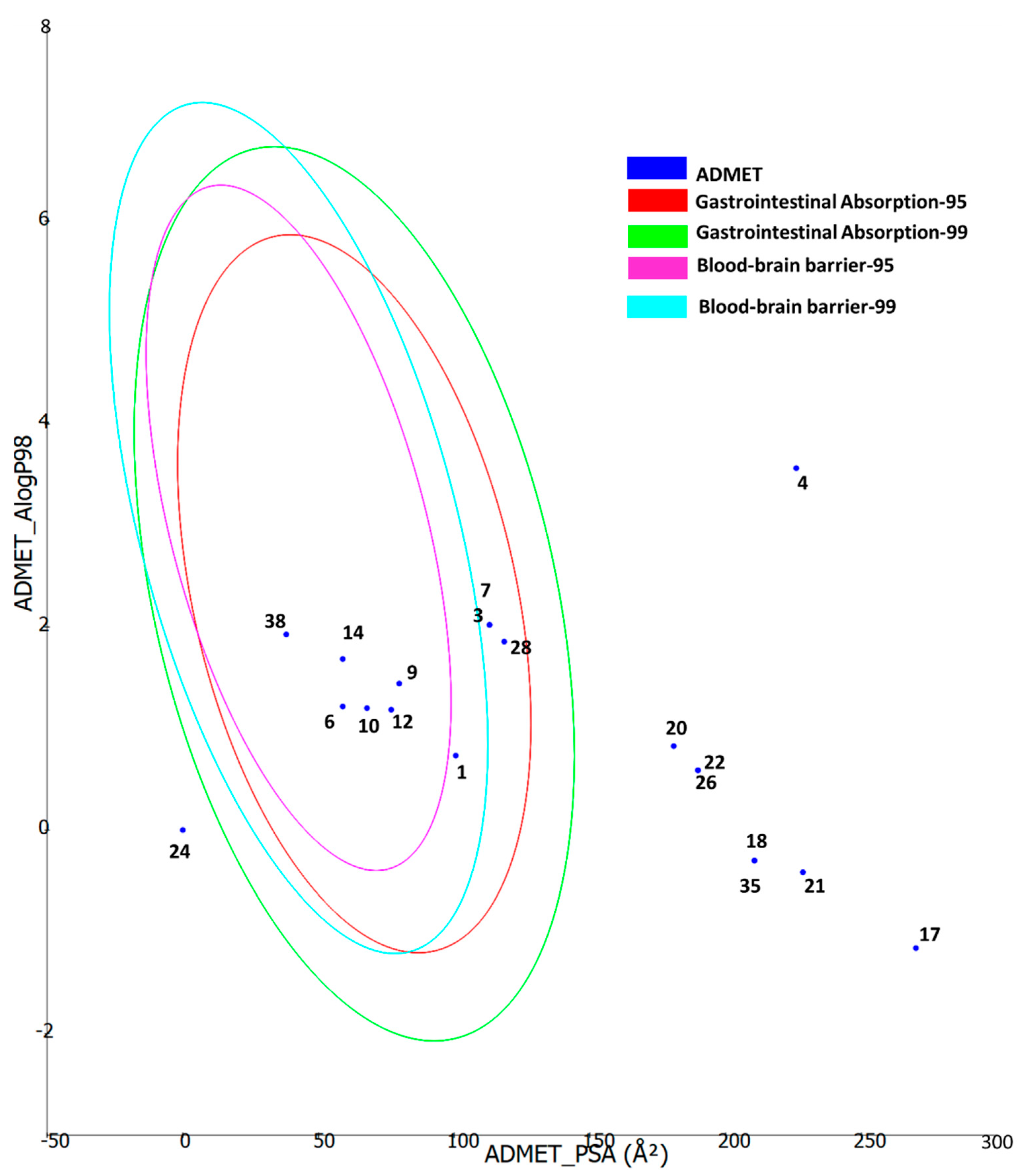
2.6. Molecular Modeling
3. Materials and Methods
3.1. Plant Materials
3.2. Preparation of Extracts
3.3. Determination of Total Polyphenol and Flavonoid Contents
3.4. Phytochemical Analysis by HPLC–MS/MS System
3.5. Antioxidant Assays
3.6. Enzyme Inhibitory Assays
3.7. Molecular Modeling
3.8. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling plant natural chemical diversity for drug discovery purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Al-Nemi, R.; Makki, A.A.; Sawalha, K.; Hajjar, D.; Jaremko, M. Untargeted Metabolomic Profiling and Antioxidant Capacities of Different Solvent Crude Extracts of Ephedra foeminea. Metabolites 2022, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Veiga, M.; Costa, E.M.; Silva, S.; Pintado, M. Impact of plant extracts upon human health: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 873–886. [Google Scholar] [CrossRef]
- Iqbal, A.; Sher, A.A.; Muhammad, N.; Badshah, S.L.; Emwas, A.-H.; Jaremko, M. Extraction and Fractionation of Prokinetic Phytochemicals from Chrozophora tinctoria and Their Bioactivities. Molecules 2022, 27, 4321. [Google Scholar] [CrossRef]
- Lavanya, B.; Maheswaran, A.; Vimal, N.; Vignesh, K.; Uvarani, K.; Varsha, R. An overall view of cassia species phytochemical constituents and its pharmacological uses. Int. J. Pharm. Sci. Res. 2018, 3, 47–50. [Google Scholar]
- Khurm, M.; Wang, X.; Zhang, H.; Hussain, S.N.; Qaisar, M.N.; Hayat, K.; Saqib, F.; Zhang, X.; Zhan, G.; Guo, Z. The genus Cassia, L.: Ethnopharmacological and phytochemical overview. Phytother. Res. 2021, 35, 2336–2385. [Google Scholar] [CrossRef]
- do Nascimento, M.N.G.; Martins, M.M.; Cunha, L.C.S.; de Souza Santos, P.; Goulart, L.R.; de Souza Silva, T.; Martins, C.H.G.; de Morais, S.A.L.; Pivatto, M. Antimicrobial and cytotoxic activities of Senna and Cassia species (Fabaceae) extracts. Ind. Crops Prod. 2020, 148, 112081. [Google Scholar] [CrossRef]
- de Albuquerque Melo, G.M.; Silva, M.C.R.; Guimaraes, T.P.; Pinheiro, K.M.; da Matta, C.B.B.; de Queiroz, A.C.; Pivatto, M.; da Silva Bolzani, V.; Alexandre-Moreira, M.S.; Viegas, C., Jr. Leishmanicidal activity of the crude extract, fractions and major piperidine alkaloids from the flowers of Senna spectabilis. Phytomedicine 2014, 21, 277–281. [Google Scholar] [CrossRef]
- Selegato, D.M.; Monteiro, A.F.; Vieira, N.C.; Cardoso, P.; Pavani, V.D.; Bolzani, V.S.; Castro-Gamboa, I. Update: Biological and chemical aspects of Senna spectabilis. J. Braz. Chem. Soc. 2017, 28, 415–426. [Google Scholar] [CrossRef]
- Sanoria, S.; Qadrie, Z.L.; Gautam, S.P.; Barwal, A. Cassia Fistula: Botany, phytochemistry and pharmacological leverages—A review. Int. J. Pharm. Pharm. Sci. 2020, 12, 1–7. [Google Scholar] [CrossRef]
- Abdalla, W.E.; Guma’a, A.G.N.; El Ghazali, G.E.B.; Khalid, H.E.S. An Updated Species Check-list for the Genus Cassia L. sensu lato in the Sudan. Nat. Resour. Res. 2016, 4, 1–12. [Google Scholar]
- Yagi, S.; Yagi, A. Important Medicinal Plants—Sudan, in Medicinal and Aromatic Plants of the World, in Encyclopedia of Life Support Systems (EOLSS); Eolss Publishers: Paris, France, 2021. [Google Scholar]
- Musa, S.M.; Fathelrhman, E.A.; Elsheikh, A.E.; Lubna, A.M.N.A.; Abdel, L.E.M.; Sakina, M.Y. Ethnobotanical study of medicinal plants in the Blue Nile State, South-eastern Sudan. J. Med. Plants Res. 2011, 5, 4287–4297. [Google Scholar]
- Anis, M.; Siddique, I.; Naz, R.; Ahmed, M.R.; Aref, I.M. Advances in Micropropagation of a Highly Important Cassia species—A Review. In New Perspectives in Plant Protection; Bandani, A.R., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Lodha, S.R.; Joshi, S.V.; Vyas, B.A.; Upadhye, M.C.; Kirve, M.S.; Salunke, S.S.; Kadu, S.K.; Rogye, M.V. Assessment of the antidiabetic potential of Cassia grandis using an in vivo model. J. Adv. Pharm. Technol. Res. 2010, 1, 330–333. [Google Scholar] [CrossRef]
- Yang, M.; Ma, Y.; Wang, Z.; Khan, A.; Zhou, W.; Zhao, T.; Cao, J.; Cheng, G.; Cai, S. Phenolic constituents, antioxidant and cytoprotective activities of crude extract and fractions from cultivated artichoke inflorescence. Ind. Crops Prod. 2020, 143, 111433. [Google Scholar] [CrossRef]
- Tzekiat, L.; Chiang, L.K. Total phenolics, total tannins and antioxidant activity of Cassia fistula L. extracts of bark, stem, leaf and root under different age classes. Asian J. Pharm. Res. Health Care 2013, 5, 52–57. [Google Scholar]
- Fuentes, J.A.M.; Fernández, I.M.; Fernández, H.Z.; Sánchez, J.L.; Alemán, R.S.; Navarro-Alarcon, M.; Borrás-Linares, I.; Maldonado, S.A.S. Quantification of bioactive molecules, minerals and bromatological analysis in Carao (Cassia grandis). J. Agric. Sci. 2020, 12, 88–94. [Google Scholar] [CrossRef]
- Laghari, A.Q.; Memon, S.; Nelofar, A.; Laghari, A.H. Extraction, identification and antioxidative properties of the flavonoid-rich fractions from leaves and flowers of Cassia angustifolia. Am. J. Anal. Chem. 2011, 2, 871–878. [Google Scholar] [CrossRef]
- Gololo, S.S.; Mapfumari, N.S.; Mogale, M.A. Comparative qualitative phytochemical analysis of the leaves of sena italica collected from different areas in Limpopo Province, South Africa. Int. J. Pharm. Pharm. Sci. 2018, 10, 67–71. [Google Scholar] [CrossRef]
- Gritsanapan, W.; Tantisewie, B.; Jirawongse, V. Chemical-Constituents of Cassia-Timorensis and Cassia-Grandis. J. Sci. Soc. Thail. 1984, 10, 189–190. [Google Scholar] [CrossRef]
- Verma, R.P.; Sinha, K.S. Anthraquinone ß-D-glucoside from Cassia grandis. Int. J. Pharmacogn. 1996, 34, 290–294. [Google Scholar] [CrossRef]
- Kalidhar, M.S.B. Chemical Examination of the Stems of Cassia grandis L. Indian J. Pharm. Sci. 1998, 60, 59. [Google Scholar]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Handa, T.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Phytochemical profile and antioxidant capacity of coffee plant organs compared to green and roasted coffee beans. Antioxidants 2020, 9, 93. [Google Scholar] [CrossRef] [PubMed]
- Al-Haidari, R.A.; Al-Oqail, M.M. New benzoic acid derivatives from Cassia italica growing in Saudi Arabia and their antioxidant activity. Saudi Pharm. J. 2020, 28, 1112–1117. [Google Scholar] [CrossRef]
- Fernández-Poyatos, M.d.P.; Zengin, G.; Salazar-Mendías, C.; Ruiz-Medina, A.; Sinan, K.I.; Llorent-Martínez, E.J. Study on Three Sarcocapnos Species as Potential Sources of Bioactive Compounds: Relation between Phenolic Content and Bioactivity by Multivariate Analysis. J. Anal. Methods Chem. 2020, 2020, 8885169. [Google Scholar] [CrossRef]
- Ismail, B.B.; Pu, Y.; Guo, M.; Ma, X.; Liu, D. LC-MS/QTOF identification of phytochemicals and the effects of solvents on phenolic constituents and antioxidant activity of baobab (Adansonia digitata) fruit pulp. Food Chem. 2019, 277, 279–288. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, Y.J.; Lee, H.J.; Lee, C.Y. Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef]
- Gülçin, İ. Antioxidant activity of caffeic acid (3, 4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef]
- Calderon-Montano, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Negro, C.; Longo, L.; Vasapollo, G.; Bellis, L.; Miceli, A. Biochemical, antioxidant and anti-inflammatory properties of pomegranate fruits growing in Southern Italy (Salento, Apulia). Acta Aliment. 2012, 41, 190–199. [Google Scholar] [CrossRef]
- Yagi, S.; Drouart, N.; Bourgaud, F.; Henry, M.; Chapleur, Y.; Laurain-Mattar, D. Antioxidant and antiglycation properties of Hydnora johannis roots. S. Afr. J. Bot. 2013, 84, 124–127. [Google Scholar] [CrossRef]
- Das, A.B.; Goud, V.V.; Das, C. 9-Phenolic compounds as functional ingredients in beverages. In Value-Added Ingredients and Enrichments of Beverages; Academic Press: Cambridge, MA, USA, 2019; pp. 285–323. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, L.; Wang, X.; Lan, R.; Wang, M.; Du, G.; Guan, W.; Liu, J.; Brennan, M.; Guo, H.; et al. Antioxidant activity evaluation of dietary flavonoid hyperoside using saccharomyces cerevisiae as a model. Molecules 2019, 24, 788. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.E. Enzyme inhibitors as the attractive targets for the treatment of various diseases. Curr. Med. Chem. 2019, 26, 3206–3207. [Google Scholar] [CrossRef] [PubMed]
- Prada, A.L.; Amado, J.R.R.; Keita, H.; Zapata, E.P.; Carvalho, H.; Lima, E.S.; de Sousa, T.P.; Carvalho, J.C.T. Cassia grandis fruit extract reduces the blood glucose level in alloxan-induced diabetic rats. Biomed. Pharm. 2018, 103, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Alhawarri, M.B.; Dianita, R.; Razak, K.N.A.; Mohamad, S.; Nogawa, T.; Wahab, H.A. Antioxidant, Anti-Inflammatory, and Inhibition of Acetylcholinesterase Potentials of Cassia timoriensis DC. Flowers. Molecules 2021, 26, 2594. [Google Scholar] [CrossRef]
- Jabir, N.R.; Khan, F.R.; Tabrez, S. Cholinesterase targeting by polyphenols: A therapeutic approach for the treatment of Alzheimer’s disease. CNS Neurosci. Ther. 2018, 24, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D. Anticholinesterase activity of selected phenolic acids and flavonoids-interaction testing in model solutions. Ann. Agric. Environ. Med. 2015, 22, 690–694. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K.; Zapp, J. Activity-guided isolation of cholinesterase inhibitors quercetin, rutin and kaempferol from Prunus persica fruit. Z. Für Nat. C 2020, 75, 87–96. [Google Scholar] [CrossRef]
- Si, Y.-X.; Yin, S.-J.; Oh, S.; Wang, Z.-J.; Ye, S.; Yan, L.; Yang, J.-M.; Park, Y.-D.; Lee, J.; Qian, G.-Y. An integrated study of tyrosinase inhibition by rutin: Progress using a computational simulation. J. Biomol. Struct. Dyn. 2012, 29, 999–1012. [Google Scholar] [CrossRef]
- Si, Y.-X.; Wang, Z.-J.; Park, D.; Jeong, H.O.; Ye, S.; Chung, H.Y.; Yang, J.-M.; Yin, S.-J.; Qian, G.-Y. Effects of isorhamnetin on tyrosinase: Inhibition kinetics and computational simulation. Biosci. Biotechnol. Biochem. 2012, 76, 1091–1097. [Google Scholar] [CrossRef]
- Einstein, J.W.; Mohd Rais, M.; Mohd, M.A. Comparative Evaluation of the Antidiabetic Effects of Different Parts of Cassia fistula Linn, a Southeast Asian Plant. J. Chem. 2013, 2013, 714063. [Google Scholar] [CrossRef]
- Jani, D.K.; Goswami, S. Antidiabetic activity of Cassia angustifolia Vahl. and Raphanus sativus Linn. leaf extracts. J. Tradit. Complement. Med. 2020, 10, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Malematja, R.O.; Bagla, V.P.; Njanje, I.; Mbazima, V.; Poopedi, K.W.; Mampuru, L.; Mokgotho, M.P. Potential Hypoglycaemic and Antiobesity Effects of Senna italica Leaf Acetone Extract. Evid.-Based Complement. Altern. Med. 2018, 2018, 5101656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praparatana, R.; Maliyam, P.; Barrows, L.R.; Puttarak, P. Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem. Molecules 2022, 27, 2393. [Google Scholar] [CrossRef]
- Qi, Y. Anti-diabetic activities of phenolic compounds in muscadine against alpha-glucosidase and pancreatic lipase. LWT—Food Sci. Technol. 2012, 46, 164–168. [Google Scholar] [CrossRef]
- Sarian, M.N.; Ahmed, Q.U.; Mat So’ad, S.Z.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Syed Mohamad, S.N.A.; Khatib, A.; Latip, J. Antioxidant and Antidiabetic Effects of Flavonoids: A Structure-Activity Relationship Based Study. BioMed Res. Int. 2017, 2017, 8386065. [Google Scholar] [CrossRef]
- Wairata, J.; Fadlan, A.; Setyo Purnomo, A.; Taher, M.; Ersam, T. Total phenolic and flavonoid contents, antioxidant, antidiabetic and antiplasmodial activities of Garcinia forbesii King: A correlation study. Arab. J. Chem. 2022, 15, 103541. [Google Scholar] [CrossRef]
- Halim, M.A.; Kanan, K.A.; Nahar, T.; Rahman, M.J.; Ahmed, K.S.; Hossain, H.; Mozumder, N.H.M.R.; Ahmed, M. Metabolic profiling of phenolics of the extracts from the various parts of blackberry plant (Syzygium cumini L.) and their antioxidant activities. LWT 2022, 167, 113813. [Google Scholar] [CrossRef]
- Lin, Y.; Tang, D.; Liu, X.; Cheng, J.; Wang, X.; Guo, D.; Zou, J.; Yang, H. Phenolic profile and antioxidant activity of longan pulp of different cultivars from South China. LWT 2022, 165, 113698. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Angeloni, S.; Abouelenein, D.; Acquaticci, L.; Xiao, J.; Sagratini, G.; Maggi, F.; Vittori, S.; Caprioli, G. A new HPLC-MS/MS method for the simultaneous determination of 36 polyphenols in blueberry, strawberry and their commercial products and determination of antioxidant activity. Food Chem. 2022, 367, 130743. [Google Scholar] [CrossRef] [PubMed]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Gerlits, O.; Ho, K.-Y.; Cheng, X.; Blumenthal, D.; Taylor, P.; Kovalevsky, A.; Radić, Z. A new crystal form of human acetylcholinesterase for exploratory room-temperature crystallography studies. Chem.-Biol. Interact. 2019, 309, 108698. [Google Scholar] [CrossRef]
- Rosenberry, T.L.; Brazzolotto, X.; Macdonald, I.R.; Wandhammer, M.; Trovaslet-Leroy, M.; Darvesh, S.; Nachon, F. Comparison of the Binding of Reversible Inhibitors to Human Butyrylcholinesterase and Acetylcholinesterase: A Crystallographic, Kinetic and Calorimetric Study. Molecules 2017, 22, 2098. [Google Scholar] [CrossRef]
- Ielo, L.; Deri, B.; Germanò, M.P.; Vittorio, S.; Mirabile, S.; Gitto, R.; Rapisarda, A.; Ronsisvalle, S.; Floris, S.; Pazy, Y.; et al. Exploiting the 1-(4-fluorobenzyl)piperazine fragment for the development of novel tyrosinase inhibitors as anti-melanogenic agents: Design, synthesis, structural insights and biological profile. Eur. J. Med. Chem. 2019, 178, 380–389. [Google Scholar] [CrossRef]
- Maurus, R.; Begum, A.; Williams, L.K.; Fredriksen, J.R.; Zhang, R.; Withers, S.G.; Brayer, G.D. Alternative Catalytic Anions Differentially Modulate Human α-Amylase Activity and Specificity. Biochemistry 2008, 47, 3332–3344. [Google Scholar] [CrossRef]
- Karade, S.S.; Hill, M.L.; Kiappes, J.L.; Manne, R.; Aakula, B.; Zitzmann, N.; Warfield, K.L.; Treston, A.M.; Mariuzza, R.A. N-Substituted Valiolamine Derivatives as Potent Inhibitors of Endoplasmic Reticulum α-Glucosidases I and II with Antiviral Activity. J. Med. Chem. 2021, 64, 18010–18024. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Martínez-Rosell, G.; Giorgino, T.; De Fabritiis, G. PlayMolecule ProteinPrepare: A Web Application for Protein Preparation for Molecular Dynamics Simulations. J. Chem. Inf. Modeling 2017, 57, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Miteva, M.A.; Guyon, F.; Tuffery, P. Frog2: Efficient 3D conformation ensemble generator for small compounds. Nucleic Acids Res. 2010, 38, W622–W627. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Kurt-Celep, I.; Zheleva-Dimitrova, D.; Gevrenova, R.; Uba, A.I.; Zengin, G.; Yıldıztugay, E.; Picot-Allain, C.M.N.; Lorenzo, J.M.; Mahomoodally, M.F.; Montesano, D. An In-Depth Study on the Metabolite Profile and Biological Properties of Primula auriculata Extracts: A Fascinating Sparkle on the Way from Nature to Functional Applications. Antioxidants 2022, 11, 1377. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Uba, A.I.; Ocal, M.; Sharifi-Rad, M.; Caprioli, G.; Angeloni, S.; Altunoglu, Y.C.; Baloglu, M.C.; Yıldıztugay, E. Integration of in vitro and in silico approaches to assess three Astragalus species from Turkey flora: A novel spotlight from lab bench to functional applications. Food Biosci. 2022, 49, 101858. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Name | TPC (mg GAE/g) | TFC (mg RE/g) |
|---|---|---|
| Cassia fistula | 49.75 ± 0.37 a | 39.15 ± 0.29 c |
| Cassia grandis | 42.23 ± 0.27 b | 30.38 ± 0.22 d |
| Senna alexandrina | 22.25 ± 0.29 c | 59.24 ± 0.08 a |
| Senna italica | 21.54 ± 0.28 c | 41.47 ± 0.28 b |
| No | Compound | C. fistula | C. grandis | S. alexandrina | S. italica |
|---|---|---|---|---|---|
| 1 | Gallic acid | 37.80 | 88.03 | 53.98 | 37.53 |
| 2 | Neochlorogenic acid | 0.15 | n.d. | 0.69 | n.d. |
| 3 | Catechin | 1361.85 | 32.94 | 3.19 | n.d. |
| 4 | Procyanidin B2 | 1767.78 | 62.43 | n.d. | 17.20 |
| 5 | Chlorogenic acid | 2.29 | 3.55 | 11.45 | 5.54 |
| 6 | 4-Hydroxy benzoic acid | 200.01 | 466.62 | 364.13 | 258.74 |
| 7 | Epicatechin | 8928.75 | 492.80 | 5.72 | n.d. |
| 8 | 3-Hydroxybenzoic acid | n.d. | n.d. | n.d. | n.d. |
| 9 | Caffeic acid | 101.65 | 76.69 | 63.16 | 48.40 |
| 10 | Vanillic acid | 179.59 | 207.39 | 102.62 | 155.01 |
| 11 | Resveratrol | n.d. | n.d. | n.d. | n.d. |
| 12 | Syringic acid | 112.42 | 85.02 | 24.59 | 78.69 |
| 13 | Procyanidin A2 | 35.90 | 7.34 | 3.63 | 4.60 |
| 14 | P-Coumaric acid | 177.89 | 198.16 | 354.55 | 314.18 |
| 15 | Ferulic acid | 21.43 | 5.52 | 49.04 | 7.73 |
| 16 | 3,5-Dicaffeoylquinic acid | 0.67 | n.d. | 0.61 | n.d. |
| 17 | Rutin | 684.39 | 2.89 | 6381.85 | 17,285.02 |
| 18 | Isoquercitrin | 6526.84 | 3169.68 | 2752.63 | 1042.35 |
| 19 | Delphindin 3,5 diglucoside | 4907.12 | 2423.86 | 2071.99 | 810.07 |
| 20 | Phloridzin | 101.26 | 2106.51 | 0.56 | 10.85 |
| 21 | Naringin | 163.84 | 444.87 | n.d. | 16.14 |
| 22 | Quercitrin | 1563.25 | 139.76 | 5.30 | 2.82 |
| 23 | Myricetin | 4.26 | 1.28 | 0.71 | 1.56 |
| 24 | Kaempferol-3-glucoside | 3398.12 | 47,360.04 | 3019.95 | 1082.75 |
| 25 | Ellagic acid | 10.62 | 116.46 | 8.61 | n.d. |
| 26 | Quercetin | 106.15 | 126.86 | 522.83 | 1207.88 |
| 27 | Phloretin | 0.89 | 16.65 | 0.05 | n.d. |
| 28 | Isorhamnetin | 7.83 | 6.92 | 454.47 | 411.96 |
| 29 | Delphindin3-galactoside | n.d. | n.d. | n.d. | n.d. |
| 30 | Cyanidin-3-glucoside | 18.27 | 24.23 | 56.93 | 11.18 |
| 31 | Petunidin-3-glucoside | n.d. | n.d. | 2.51 | n.d. |
| 32 | Pelargonidin-3-rutinoside | n.d. | n.d. | n.d. | n.d. |
| 33 | Pelargonidin-3-glucoside | n.d. | n.d. | n.d. | n.d. |
| 34 | Malvidin-3-galactoside | n.d. | n.d. | 14.19 | n.d. |
| 35 | Hyperoside | 8270.74 | 3634.40 | 3299.95 | 1293.52 |
| 36 | Hesperidin | n.d. | 12.34 | 56.04 | 74.06 |
| 37 | Kaempferol | 28.04 | 49.89 | 16.81 | 37.98 |
| 38 | Trans-cinnamic acid | 177.68 | 3851.66 | 2095.98 | 803.88 |
| Total content | 38,897.47 | 65,214.79 | 21,798.71 | 25,019.64 | |
| Plant name | DPPH (mg TE */g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | MCA (mg EDTAE **/g) | PBD (mmol TE/g) |
|---|---|---|---|---|---|---|
| Cassia fistula | 77.36 ± 0.69 a | 218.44 ± 13.8 a | 215.33 ± 4.37 a | 106.34 ± 5.88 a | 5,68 ± 0.55 c | 1.94 ± 0.13 a |
| Cassia grandis | 34.74 ± 0.26 b | 78.43 ± 0.08 b | 152.79 ± 2.19 b | 62.68 ± 0.24 b | 11.38 ± 0.96 a | 1.48 ± 0.12 b |
| Senna alexandrina | 24.74 ± 0.64 c | 54.49 ± 1.28 c | 88.45 ± 2.11 c | 44.02 ± 0.92 c | na | 1.11 ± 0.05 c |
| Senna italica | 18.60 ± 0.20 d | 51.98± 0.05 c | 84.60 ± 2.54 c | 44.58 ± 0.26 c | 8.11 ± 0.72 b | 1.07 ± 0.11 c |
| Plant name | AChE (mg GALAE */g) | BChE (mg GALAE/g) | Tyrosinase (mg KAE **/g) | Amylase (mmol ACAE ***/g) | Glucosidase (mmol ACAE/g) |
|---|---|---|---|---|---|
| Cassia fistula | 1.71 ± 0.20 b | 0.92 ± 0.02 a | 34.51 ± 0.93 b | 0.40 ± 0.02 b | 1.45 ± 0.01 a |
| Cassia grandis | 2.17 ± 0.22 a | 1.15 ± 0.18 a | 46.58 ± 0.32 a | 0.48 ± 0.02 a | 1.44 ± 0.00 ab |
| Senna alexandrina | 2.41 ± 0.07 a | na | 15.12 ± 0.59 d | 0.37 ± 0.02 b | 1.43 ± 0.00 b |
| Senna italica | 2.05 ± 0.11 ab | 0.64 ± 0.02 b | 21.17 ± 0.38 c | 0.39 ± 0.01 b | 1.44 ± 0.01 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omer, H.A.A.; Caprioli, G.; Abouelenein, D.; Mustafa, A.M.; Uba, A.I.; Ak, G.; Ozturk, R.B.; Zengin, G.; Yagi, S. Phenolic Profile, Antioxidant and Enzyme Inhibitory Activities of Leaves from Two Cassia and Two Senna Species. Molecules 2022, 27, 5590. https://doi.org/10.3390/molecules27175590
Omer HAA, Caprioli G, Abouelenein D, Mustafa AM, Uba AI, Ak G, Ozturk RB, Zengin G, Yagi S. Phenolic Profile, Antioxidant and Enzyme Inhibitory Activities of Leaves from Two Cassia and Two Senna Species. Molecules. 2022; 27(17):5590. https://doi.org/10.3390/molecules27175590
Chicago/Turabian StyleOmer, Haifa A. A., Giovanni Caprioli, Doaa Abouelenein, Ahmed M. Mustafa, Abdullahi Ibrahim Uba, Gunes Ak, Refiye Beyza Ozturk, Gokhan Zengin, and Sakina Yagi. 2022. "Phenolic Profile, Antioxidant and Enzyme Inhibitory Activities of Leaves from Two Cassia and Two Senna Species" Molecules 27, no. 17: 5590. https://doi.org/10.3390/molecules27175590
APA StyleOmer, H. A. A., Caprioli, G., Abouelenein, D., Mustafa, A. M., Uba, A. I., Ak, G., Ozturk, R. B., Zengin, G., & Yagi, S. (2022). Phenolic Profile, Antioxidant and Enzyme Inhibitory Activities of Leaves from Two Cassia and Two Senna Species. Molecules, 27(17), 5590. https://doi.org/10.3390/molecules27175590