

Characterization of Polyphenolic Compounds from Bacopa procumbens and Their Effects on Wound-Healing Process

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Phytochemical Screening of Bacopa procumbens Aqueous Fraction
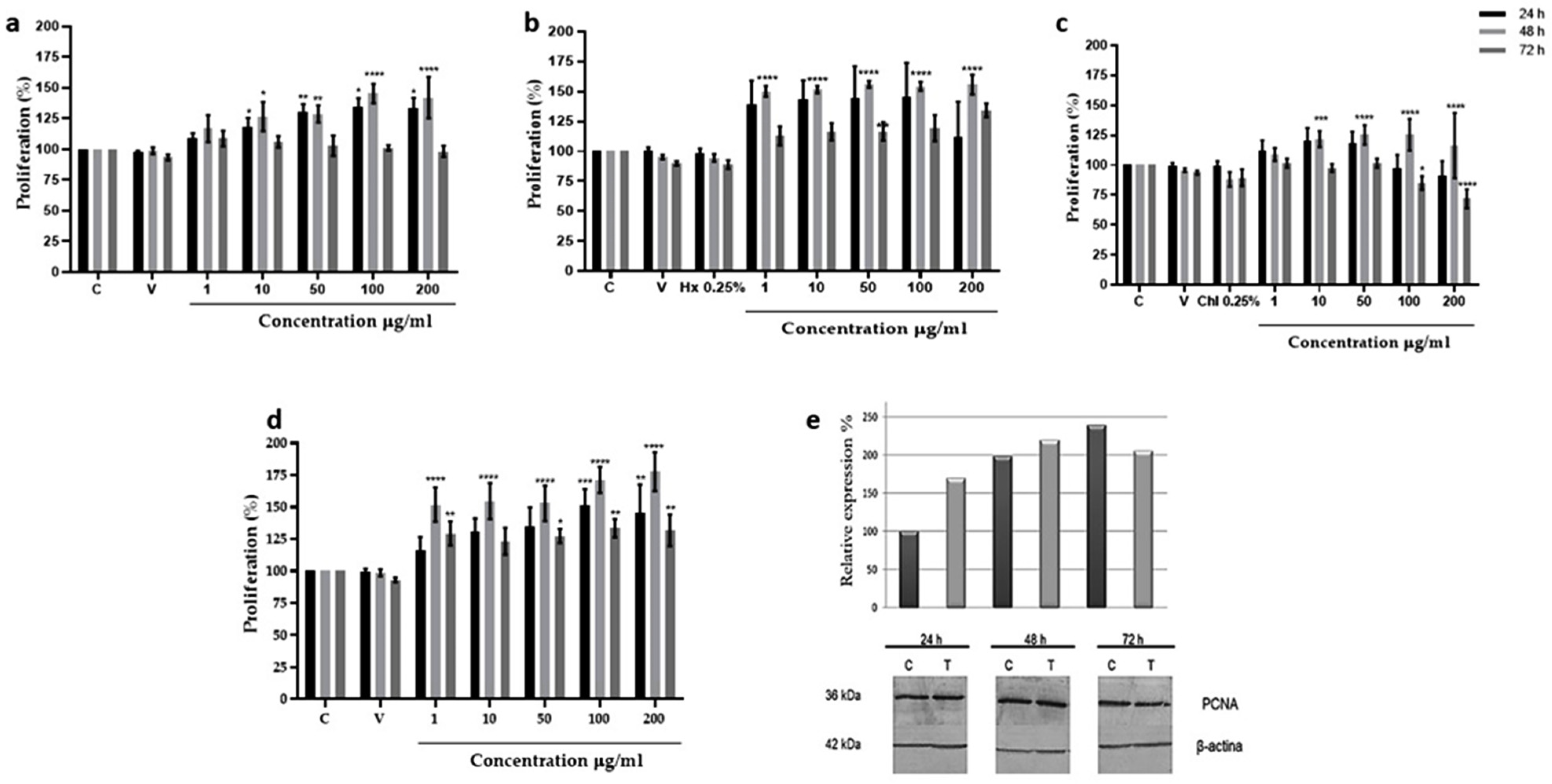
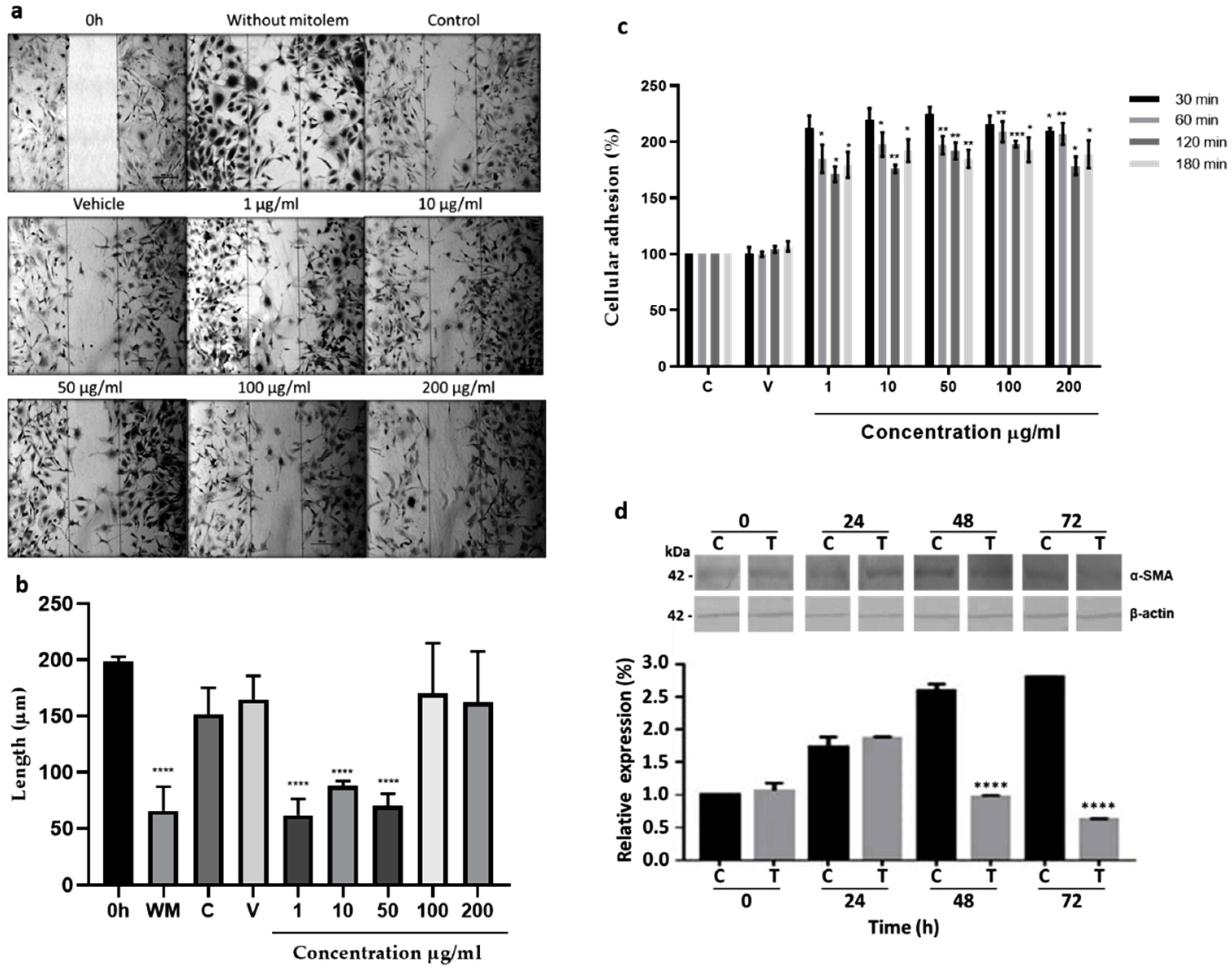
2.2. In Vitro Assays
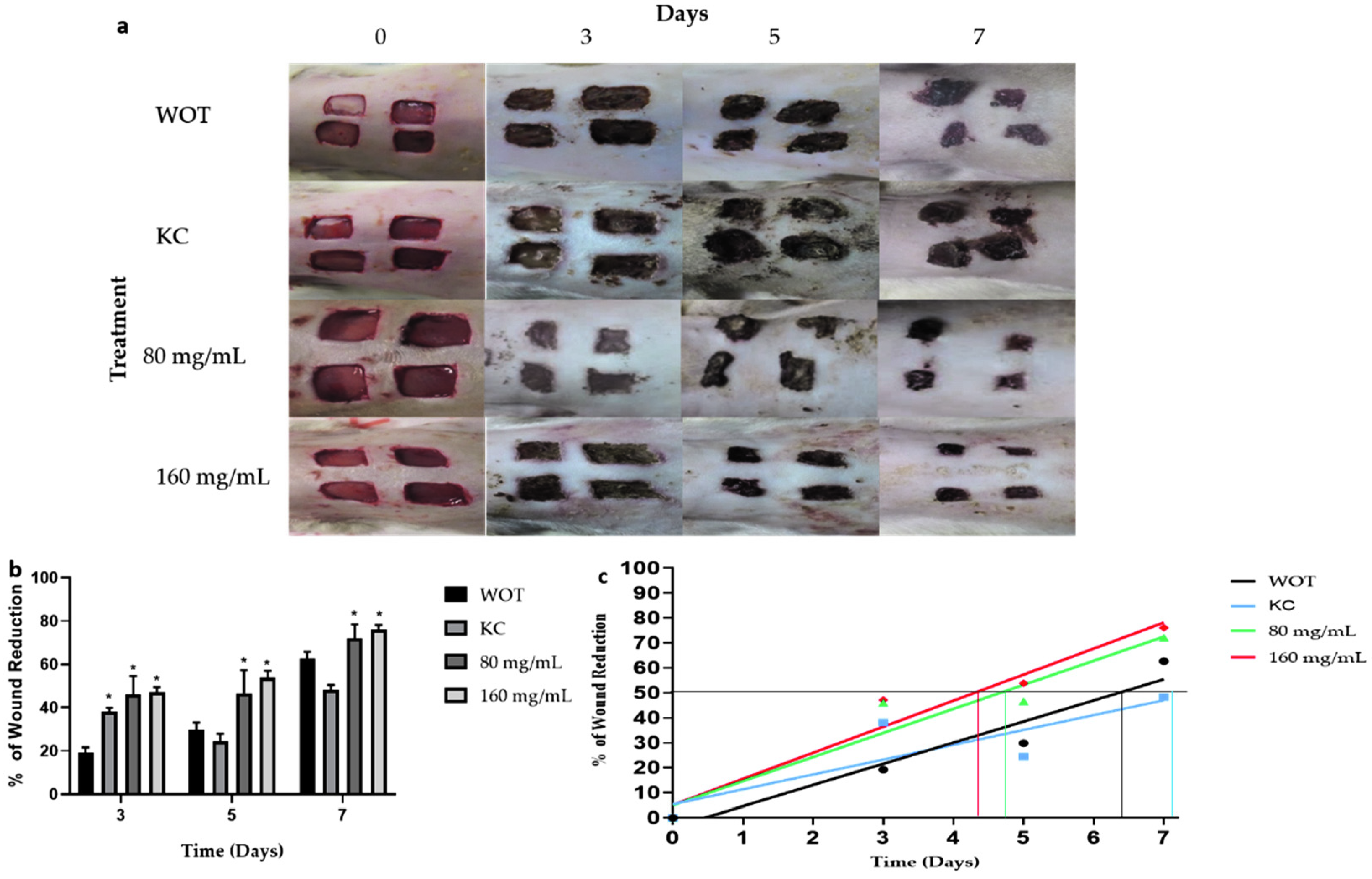
2.3. Macroscopic Wound Repair Effects of Polyphenolic Compounds from Bacopa procumbens
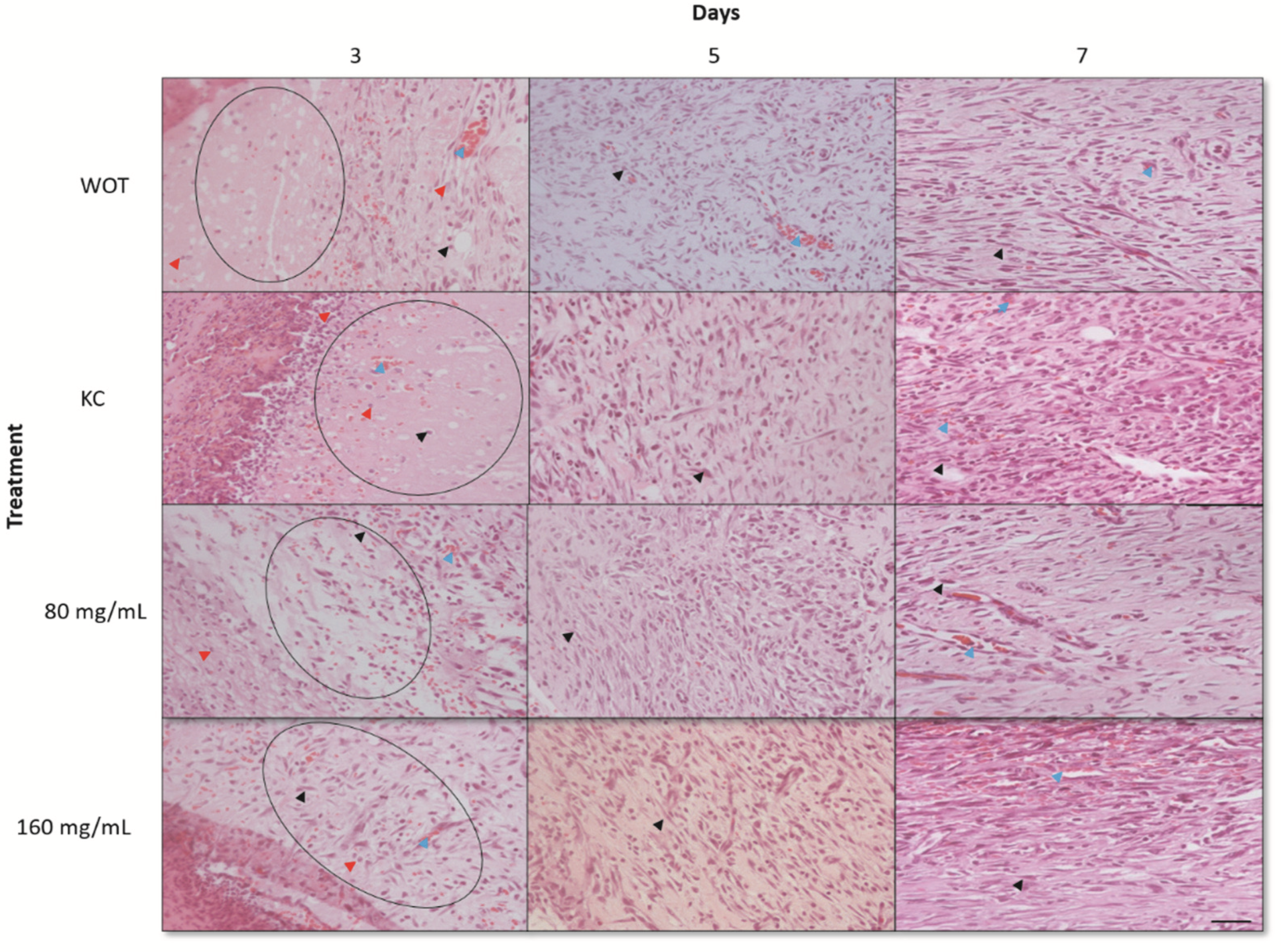
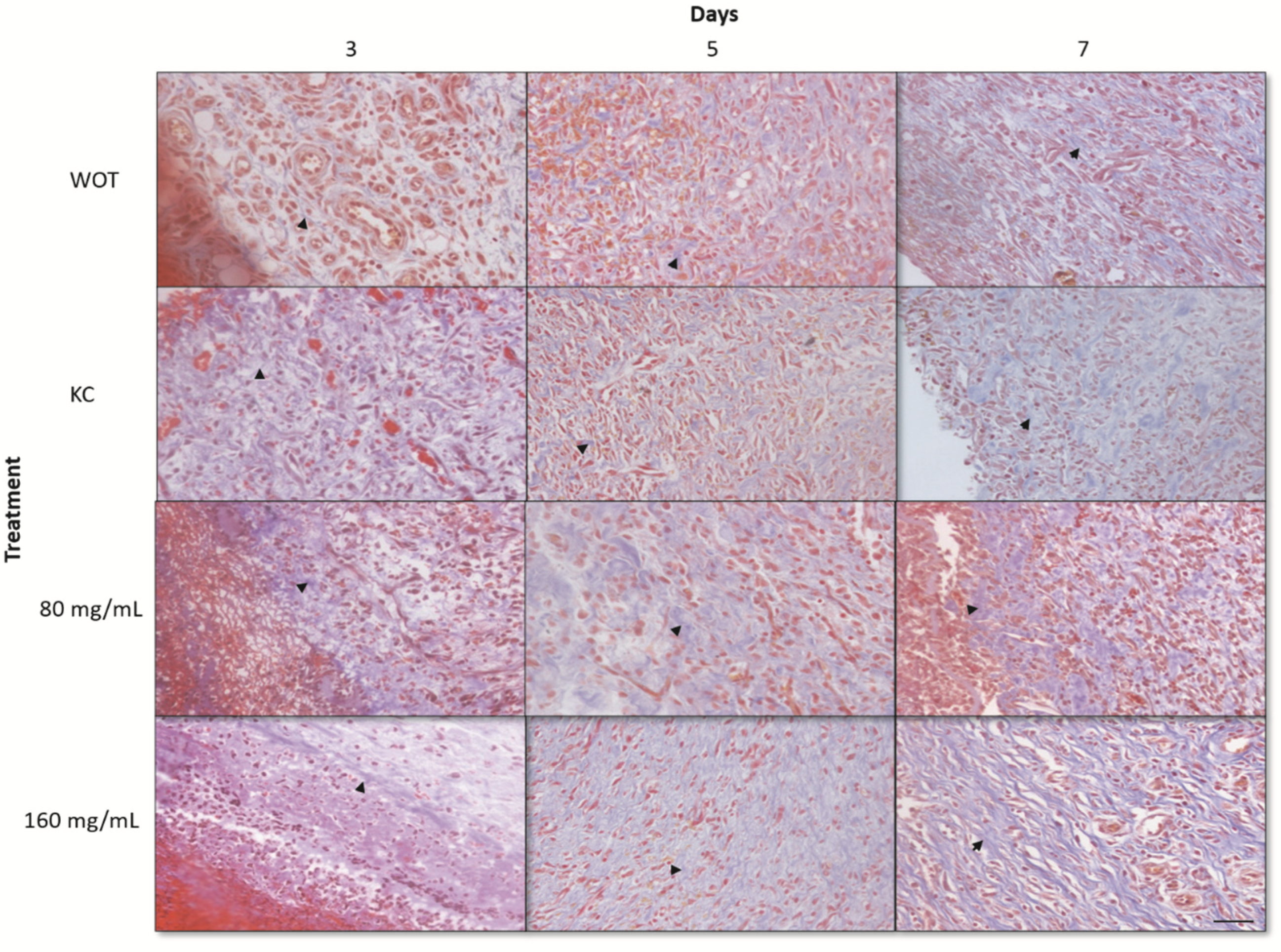
2.4. Histological Findings of Wound Healing from Different Groups
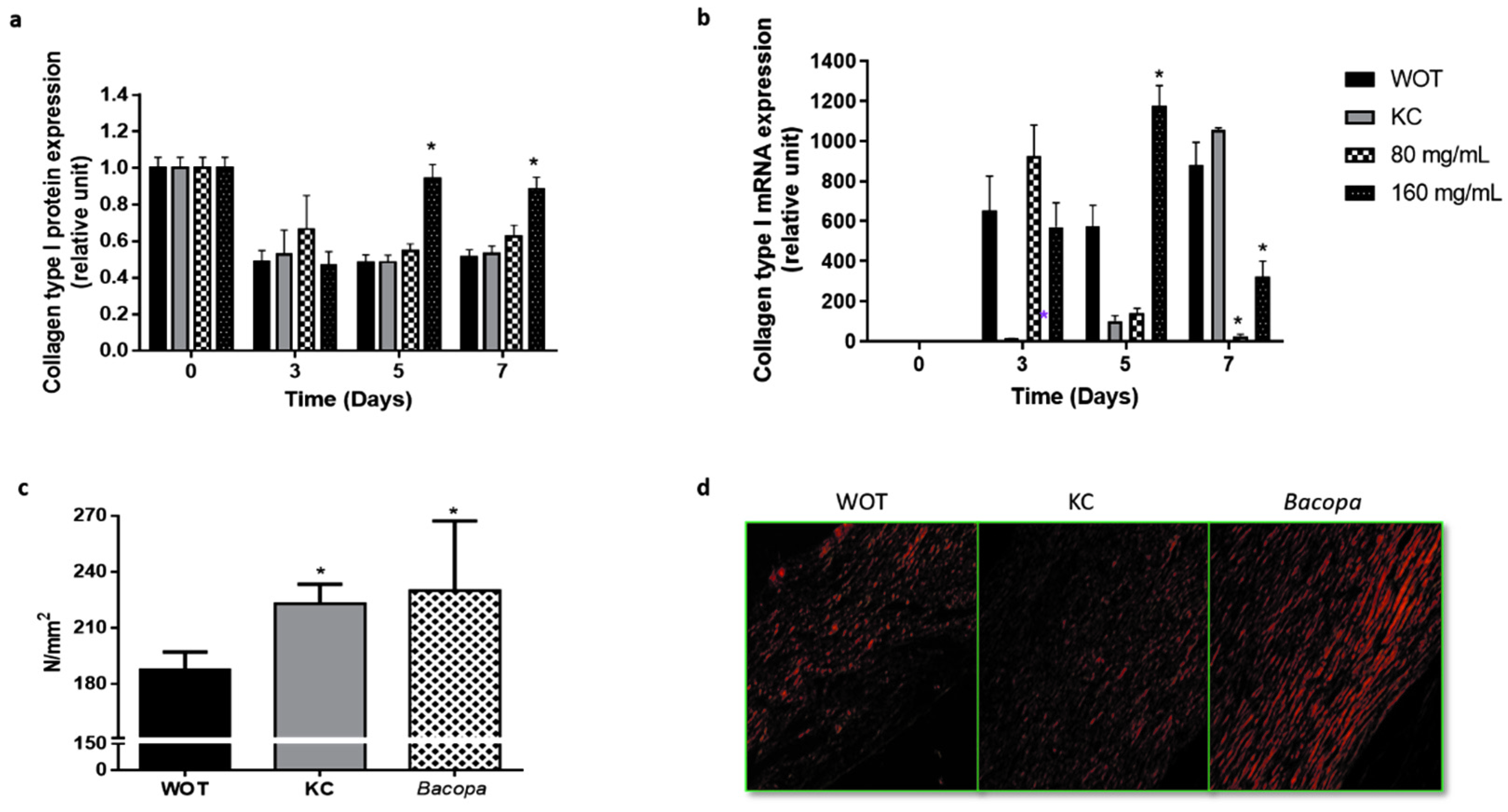
2.5. Bacopa Treatment Produced Scars with Better Mechanical Properties
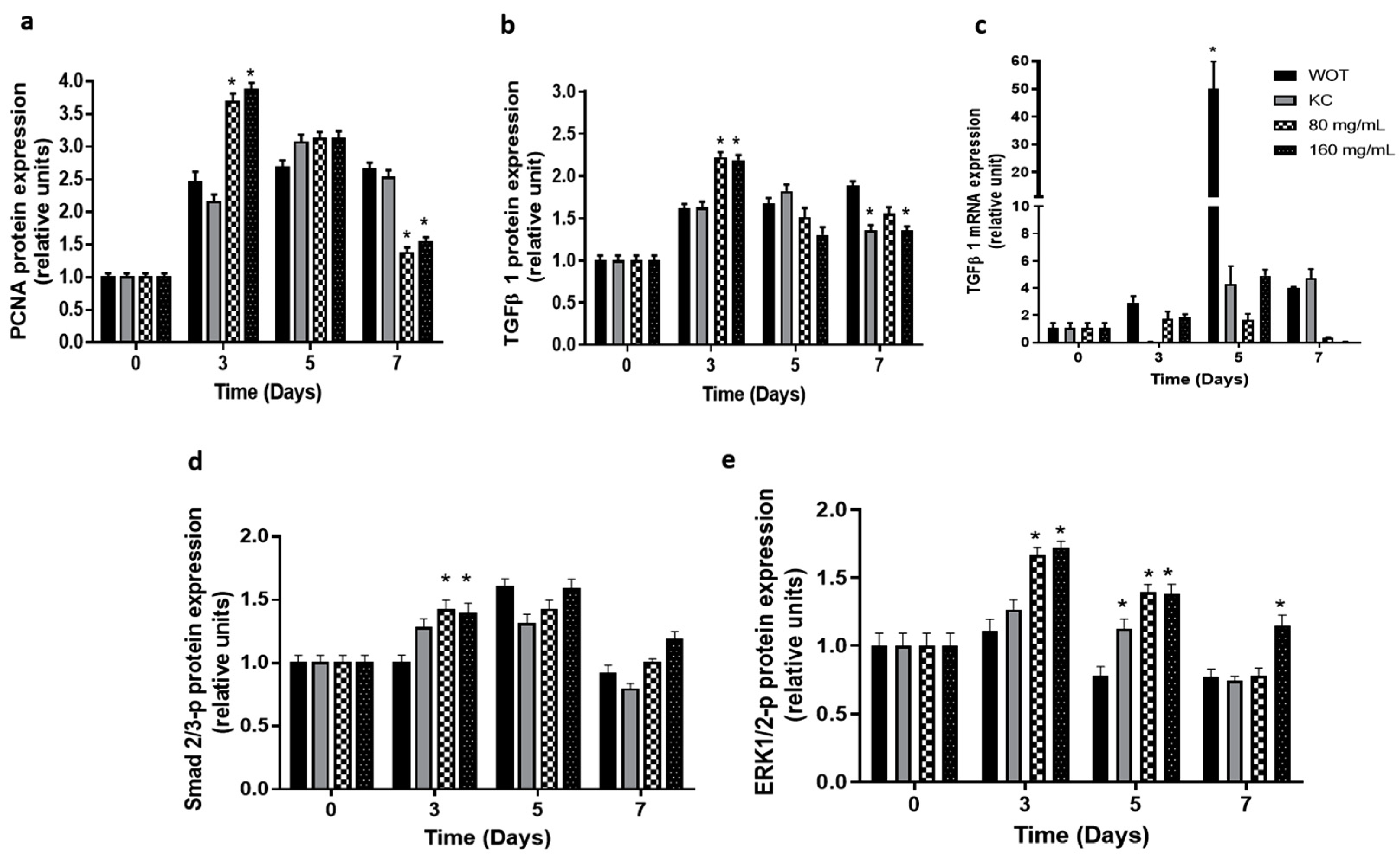
2.6. Key Molecular Effectors of Wound Repair Induced by Bacopa
3. Discussion
4. Materials and Methods
4.1. Preparation of Aquoethanolic Extract and Hydrogel Formulation
4.2. In Vitro Model
4.2.1. Cell Culture
4.2.2. Cell Viability Assay
4.2.3. In Vitro Scratch Wound-Healing Assay
4.2.4. Cell Adhesion Assay
4.2.5. Western Blotting Analysis
4.3. In Vivo Skin Wound Model
4.3.1. Animals
4.3.2. Immunohistochemistry
4.3.3. Quantitative Real-Time PCR
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Singer, A.J.; Clark, R.A. Cutaneous Wound Healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Baum, C.L.; Arpey, C.J. Normal Cutaneous Wound Healing: Clinical Correlation with Cellular and Molecular Events. Dermatol. Surg. 2006, 31, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.H.; Liu, X.; To, M.K.; Yeung, K.; Ho, C.-M.; Wong, K.K. Modulation of collagen alignment by silver nanoparticles results in better mechanical properties in wound healing. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 497–504. [Google Scholar] [CrossRef] [PubMed]
- A Border, W.; Ruoslahti, E. Transforming growth factor-beta in disease: The dark side of tissue repair. J. Clin. Investig. 1992, 90, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrecchia, F.; Mauviel, A. Transforming growth factor-β and fibrosis. World J. Gastroenterol. 2007, 13, 3056–3062. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Nguyen, C.; Zhang, X.; Khorasani, H.; Wang, J.Z.; Zara, J.N.; Chu, F.; Yin, W.; Pang, S.; Le, A.; et al. Delayed Wound Closure in Fibromodulin-Deficient Mice Is Associated with Increased TGF-β3 Signaling. J. Investig. Dermatol. 2011, 131, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Massagué, J. How cells read TGF-β signals. Nat. Rev. Mol. Cell Biol. 2000, 1, 169–178. [Google Scholar] [CrossRef]
- Mecott-Rivera, G.; Aguilar-Baqueiro, J.A.; Bracho, S.; Miranda-Maldonado, I.; Franco-Márquez, R.; Castro-Govea, Y.; Dorsey-Treviño, E.G.; García-Pérez, M.M. Pirfenidone increases the epithelialization rate of skin graft donor sites. Burns 2018, 44, 2051–2058. [Google Scholar] [CrossRef]
- Molina-Mendoza, J.L.; Galván-Villanueva, R.; Patiño-Siciliano, A.; Fernández-Nava, R. Medicinal plants and preliminary floristic list of the municipality of Huasca de Ocampo, Hidalgo, Mexico. Polibotanica 2012, 34, 259–291. [Google Scholar]
- Velidandla, S.; Gaikwad, P.; Ealla, K.K.R.; Bhorgonde, K.D.; Hunsingi, P.; Kumar, A. Histochemical analysis of polarizing colors of collagen using Picrosirius Red staining in oral submucous fibrosis. J. Int. Oral Health 2014, 6, 33–38. [Google Scholar] [PubMed]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.E. Non-Smad pathways in TGF-β signaling. Cell Res. 2009, 19, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, B.; Vijayakumar, M.; Govindarajan, R.; Pushpangadan, P. Ethnopharmacological approaches to wound healing—Exploring medicinal plants of India. J. Ethnopharmacol. 2007, 114, 103–113. [Google Scholar] [CrossRef]
- Abbasi, A.M.; Khan, M.; Ahmad, M.; Zafar, M.; Jahan, S.; Sultana, S. Ethnopharmacological application of medicinal plants to cure skin diseases and in folk cosmetics among the tribal communities of North-West Frontier Province, Pakistan. J. Ethnopharmacol. 2010, 128, 322–335. [Google Scholar] [CrossRef]
- Pathak, A.; Kulshreshtha, D.; Maurya, R. Chemical constituents of Bacopa procumbens. Nat. Prod. Res. 2005, 19, 131–136. [Google Scholar] [CrossRef]
- González-Cortazar, M.; López-Gayou, V.; Tortoriello, J.; Domínguez-Mendoza, B.E.; Ríos-Cortes, A.M.; Delgado-Macuil, R.; Hernández-Beteta, E.E.; Blé-González, E.A.; Zamilpa, A. Antimicrobial gastrodin derivatives isolated from Bacopa procumbens. Phytochem. Lett. 2019, 31, 33–38. [Google Scholar] [CrossRef]
- Chen, L.-Y.; Huang, C.-N.; Liao, C.-K.; Chang, H.-M.; Kuan, Y.-H.; Tseng, T.-J.; Yen, K.-J.; Yang, K.-L.; Lin, H.-C. Effects of Rutin on Wound Healing in Hyperglycemic Rats. Antioxidants 2020, 9, 1122. [Google Scholar] [CrossRef]
- Sun, X.; Wang, X.; Zhao, Z.; Chen, J.; Li, C.; Zhao, G. Paeoniflorin accelerates foot wound healing in diabetic rats though activating the Nrf2 pathway. Acta Histochem. 2020, 122, 151649. [Google Scholar] [CrossRef]
- Al-Roujayee, A.S. Naringenin improves the healing process of thermally-induced skin damage in rats. J. Int. Med. Res. 2017, 45, 570–582. [Google Scholar] [CrossRef] [Green Version]
- Sharath, R.; Harish, B.; Krishna, V.; Sathyanarayana, B.; Swamy, H.K. Wound healing and protease inhibition activity of Bacoside-A, isolated from Bacopa monnieri wettest. Phytother. Res. 2010, 24, 1217–1222. [Google Scholar] [CrossRef]
- Murthy, S.; Gautam, M.K.; Goel, S.; Purohit, V.; Sharma, H.; Goel, R.K. Evaluation of In Vivo Wound Healing Activity of Bacopa monniera on Different Wound Model in Rats. BioMed Res. Int. 2013, 2013, 972028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.C.W.; Lau, K.M.; Liu, C.L.; To, M.H.; Kwok, H.F.; Lai, K.K.; Lau, C.P.; Ko, C.H.; Leung, P.C.; Fung, K.P.; et al. The in vivo and in vitro diabetic wound healing effects of a 2-herb formula and its mechanisms of action. J. Ethnopharmacol. 2011, 134, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Freiesleben, S.H.; Soelberg, J.; Nyberg, N.T.; Jäger, A.K. Determination of the Wound Healing Potentials of Medicinal Plants Historically Used in Ghana. Evid.-Based Complement. Altern. Med. 2017, 2017, 9480791. [Google Scholar] [CrossRef] [Green Version]
- Pitz, H.D.S.; Pereira, A.; Blasius, M.B.; Voytena, A.P.L.; Affonso, R.C.L.; Fanan, S.; Trevisan, A.C.D.; Ribeiro-Do-Valle, R.M.; Maraschin, M. In Vitro Evaluation of the Antioxidant Activity and Wound Healing Properties of Jaboticaba (Plinia peruviana) Fruit Peel Hydroalcoholic Extract. Oxidative Med. Cell. Longev. 2016, 2016, 3403586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alerico, G.C.; Beckenkamp, A.; Vignoli-Silva, M.; Buffon, A.; von Poser, G.L. Proliferative effect of plants used for wound healing in Rio Grande do Sul state, Brazil. J. Ethnopharmacol. 2015, 176, 305–310. [Google Scholar] [CrossRef]
- Yamada, K.M. Fibronectin peptides in cell migration and wound repair. J. Clin. Investig. 2000, 105, 1507–1509. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.B.; Pang, B.; Gardner, D.J.; Niknam-Benia, S.; Soundarajan, V.; Bramos, A.; Perrault, D.P.; Banks, K.; Lee, G.K.; Baker, R.Y.; et al. Topical Fibronectin Improves Wound Healing of Irradiated Skin. Sci. Rep. 2017, 7, 3876. [Google Scholar] [CrossRef]
- De Martino, L.; Mencherini, T.; Mancini, E.; Aquino, R.P.; De Almeida, L.F.R.; De Feo, V. In Vitro Phytotoxicity and Antioxidant Activity of Selected Flavonoids. Int. J. Mol. Sci. 2012, 13, 5406–5419. [Google Scholar] [CrossRef]
- Raimondi, L.; Banchelli, G.; Dalmazzi, D.; Mulinacci, N.; Romani, A.; Vincieri, F.F.; Pirisino, R. Sedum telephium L. Polysaccharide Content Affects MRC5 Cell Adhesion to Laminin and Fibronectin. J. Pharm. Pharmacol. 2000, 52, 585–591. [Google Scholar] [CrossRef]
- Gál, P.; Vasilenko, T.; Kováč, I.; Kostelníková, M.; Jakubčo, J.; Szabo, P.; Dvořánková, B.; Sabol, F.; Gabius, H.-J.; Smetana, K., Jr. Atropa belladonna L. Water Extract: Modulator of Extracellular Matrix Formation in Vitro and in Vivo. Physiol. Res. 2012, 61, 241–250. [Google Scholar] [CrossRef] [PubMed]
- di Martino, O.; Tito, A.; De Lucia, A.; Cimmino, A.; Cicotti, F.; Apone, F.; Colucci, G.; Calabrò, V. Hibiscus syriacus Extract from an Established Cell Culture Stimulates Skin Wound Healing. BioMed Res. Int. 2017, 2017, 7932019. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.H.; Lee, G.W.; Park, K.H.; Mohamed, M.A.A.; Bang, M.H.; Baek, Y.S.; Son, Y.; Chung, D.K.; Baek, N.-I.; Kim, J. The stimulatory effects of Stewartia koreana extract on the proliferation and migration of fibroblasts and the wound healing activity of the extract in mice. Int. J. Mol. Med. 2014, 34, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Talekar, Y.P.; Apte, K.G.; Paygude, S.V.; Tondare, P.R.; Parab, P.B. Studies on wound healing potential of polyherbal formulation using in vitro and in vivo assays. J. Ayurveda Integr. Med. 2017, 8, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, Y. Dissection of Key Events in Tubular Epithelial to Myofibroblast Transition and Its Implications in Renal Interstitial Fibrosis. Am. J. Pathol. 2001, 159, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.M.; Chen, L.; Bond, J.E.; Medina, M.A.; Ren, L.; Kokosis, G.; Selim, A.M.; Levinson, H. Myofibroblasts contribute to but are not necessary for wound contraction. Lab. Investig. 2015, 95, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Van De Water, L.; Varney, S.; Tomasek, J.J. Mechanoregulation of the Myofibroblast in Wound Contraction, Scarring, and Fibrosis: Opportunities for New Therapeutic Intervention. Adv. Wound Care 2013, 2, 122–141. [Google Scholar] [CrossRef] [Green Version]
- Suntar, I.; Akkol, E.K.; Keleş, H.; Yesilada, E.; Sarker, S.D. Exploration of the wound healing potential of Helichrysum graveolens (Bieb.) Sweet: Isolation of apigenin as an active component. J. Ethnopharmacol. 2013, 149, 103–110. [Google Scholar] [CrossRef]
- Nazac, A.; Bancelin, S.; Teig, B.; Ibrahim, B.H.; Fernandez, H.; Schanne-Klein, M.-C.; De Martino, A. Optimization of Picrosirius red staining protocol to determine collagen fiber orientations in vaginal and uterine cervical tissues by Mueller polarized microscopy. Microsc. Res. Tech. 2015, 78, 723–730. [Google Scholar] [CrossRef]
- Ganeshkumar, M.; Ponrasu, T.; Krithika, R.; Iyappan, K.; Gayathri, V.S.; Suguna, L. Topical application of Acalypha indica accelerates rat cutaneous wound healing by up-regulating the expression of Type I and III collagen. J. Ethnopharmacol. 2012, 142, 14–22. [Google Scholar] [CrossRef]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [PubMed]
- Finnson, K.W.; McLean, S.; Di Guglielmo, G.M.; Philip, A. Dynamics of Transforming Growth Factor Beta Signaling in Wound Healing and Scarring. Adv. Wound Care 2013, 2, 195–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakytny, R.; Brunner, G. Extracellular regulation of TGF-β activity in wound repair: Growth factor latency as a sensor mechanism for injury. Thromb. Haemost. 2004, 92, 253–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Fong, C.C.; Yu, W.K.; Chen, Y.; Wei, F.; Koon, C.M.; Lau, K.M.; Leung, P.C.; Lau, C.B.S.; Fung, K.P.; et al. Herbal formula Astragali Radix and Rehmanniae Radix exerted wound healing effect on human skin fibroblast cell line Hs27 via the activation of transformation growth factor (TGF-β) pathway and promoting extracellular matrix (ECM) deposition. Phytomedicine 2012, 20, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Suh, N.; Roberts, A.B.; Reffey, S.B.; Miyazono, K.; Itoh, S.; Dijke, P.T.; Heiss, E.; E Place, A.; Risingsong, R.; Williams, C.R.; et al. Synthetic triterpenoids enhance transforming growth factor beta/Smad signaling. Cancer Res. 2003, 63, 1371–1376. [Google Scholar] [PubMed]
- Huh, J.-E.; Nam, D.-W.; Baek, Y.-H.; Kang, J.W.; Park, D.-S.; Choi, D.-Y.; Lee, J.-D. Formononetin accelerates wound repair by the regulation of early growth response factor-1 transcription factor through the phosphorylation of the ERK and p38 MAPK pathways. Int. Immunopharmacol. 2011, 11, 46–54. [Google Scholar] [CrossRef]
- Kim, M.; Lee, H.J.; Randy, A.; Yun, J.H.; Oh, S.-R.; Nho, C.W. Stellera chamaejasme and its constituents induce cutaneous wound healing and anti-inflammatory activities. Sci. Rep. 2017, 7, srep42490. [Google Scholar] [CrossRef] [Green Version]
- Dhanani, T.; Shah, S.; Gajbhiye, N.A.; Kumar, S. Effect of extraction methods on yield, phytochemical constituents and antioxidant activity of Withania somnifera. Arab. J. Chem. 2017, 10, S1193–S1199. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, L.G.; Wu, X.; Guan, J.L. Wound-Healing Assay. In Cell Migration, Guan, JL. Ed.; Methods in Molecular Biology™; Humana Press: Totowa, NJ, USA, 2005; Volume 294, pp. 23–30. [Google Scholar] [CrossRef]
- Rafiee, S.; Nekouyian, N.; Sarabandi, F.; Chavoshi-Nejad, M.; Mohsenikia, M.; Yadollah-Damavandi, S.; Seifaee, A.; Jangholi, E.; Eghtedari, D.; Najafi, H.; et al. Effect of Topical Linum usitatissimum on Full Thickness Excisional Skin Wounds. Trauma Mon. 2016, 22. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Zapién, G.J.; Mata-Miranda, M.M.; García-Sánchez, F.J.; Campos-Soto, S.S.; Sánchez-Brito, M.; Martínez-Cuazitl, A. Biomolecular Characterization by FTIR Microspectroscopy in the Modeling Phase of Wound Cicatrization in a Murine Model of Excisional Injury. Int. J. Morphol. 2019, 37, 1234–1244. [Google Scholar] [CrossRef]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol Treats UVB-Induced Photoaging by Anti-MMP Expression, through Anti-Inflammatory, Antioxidant, and Antiapoptotic Properties, and Treats Photoaging by Upregulating VEGF-B Expression. Oxidative Med. Cell. Longev. 2022, 2022, 6037303. [Google Scholar] [CrossRef] [PubMed]
- Rittié, L. Method for Picrosirius Red-Polarization Detection of Collagen Fibers in Tissue Sections. In Fibrosis; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2017; Volume 1627, pp. 395–407. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RT (min) | [M−H]– | Reference Mass |
|---|---|---|---|
| Arbutin | 0.386 | 271.0821 | 272.0896 |
| o-Hydroxybenzoic acid | 0.924 | 137.0242 | 138.0317 |
| m-Hydroxybenzoic acid | 0.959 | 137.0245 | 138.0317 |
| Feruloyl glucose | 1.161 | 355.1033 | 356.1107 |
| Esculetin | 1.263 | 177.0188 | 178.0266 |
| 3-O-Caffeoyl shikimic acid | 1.937 | 335.0771 | 336.0845 |
| Catalposide | 2.88 | 481.134 | 482.1424 |
| Homovanillyl alcohol | 3.082 | 167.0711 | 168.0786 |
| Genistein | 3.285 | 269.0452 | 270.0528 |
| p-Coumaric acid | 3.318 | 163.0396 | 164.0473 |
| Z-Astringin | 3.486 | 405.118 | 406.1264 |
| Naringenin-C-hexoside | 3.588 | 433.1126 | 434.1213 |
| Astilbin | 3.823 | 449.1092 | 450.1162 |
| m-Coumaric acid | 3.857 | 163.0398 | 164.0473 |
| Equol 7-O-glucuronide | 4.092 | 417.1183 | 418.1264 |
| Syringaresinol-glucoside | 4.093 | 579.2096 | 580.2156 |
| p-Hydroxybenzoic acid | 4.194 | 137.0243 | 138.0317 |
| Daidzein 7-O-glucuronide | 4.531 | 429.084 | 430.09 |
| 4′-Methoxyapigenin rutinoside | 5.069 | 591.1709 | 592.1792 |
| Prunetin | 5.071 | 283.0602 | 284.0685 |
| Paeoniflorin | 5.104 | 479.1539 | 480.1632 |
| Orobol | 5.204 | 285.0409 | 286.0477 |
| Phloretic acid | 5.339 | 165.0551 | 166.063 |
| Methyl ferulate | 5.34 | 207.0658 | 208.0736 |
| Urolithin C | 5.745 | 243.0296 | 244.0372 |
| Koparin | 5.912 | 299.0551 | 300.0634 |
| Coumarin | 6.115 | 145.0298 | 146.0368 |
| Stevenin | 6.822 | 283.0606 | 284.0685 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Cuazitl, A.; Gómez-García, M.d.C.; Hidalgo-Alegria, O.; Flores, O.M.; Núñez-Gastélum, J.A.; Martínez, E.S.M.; Ríos-Cortés, A.M.; Garcia-Solis, M.; Pérez-Ishiwara, D.G. Characterization of Polyphenolic Compounds from Bacopa procumbens and Their Effects on Wound-Healing Process. Molecules 2022, 27, 6521. https://doi.org/10.3390/molecules27196521
Martínez-Cuazitl A, Gómez-García MdC, Hidalgo-Alegria O, Flores OM, Núñez-Gastélum JA, Martínez ESM, Ríos-Cortés AM, Garcia-Solis M, Pérez-Ishiwara DG. Characterization of Polyphenolic Compounds from Bacopa procumbens and Their Effects on Wound-Healing Process. Molecules. 2022; 27(19):6521. https://doi.org/10.3390/molecules27196521
Chicago/Turabian StyleMartínez-Cuazitl, Adriana, María del Consuelo Gómez-García, Oriana Hidalgo-Alegria, Olivia Medel Flores, José Alberto Núñez-Gastélum, Eduardo San Martín Martínez, Ada María Ríos-Cortés, Mario Garcia-Solis, and David Guillermo Pérez-Ishiwara. 2022. "Characterization of Polyphenolic Compounds from Bacopa procumbens and Their Effects on Wound-Healing Process" Molecules 27, no. 19: 6521. https://doi.org/10.3390/molecules27196521
APA StyleMartínez-Cuazitl, A., Gómez-García, M. d. C., Hidalgo-Alegria, O., Flores, O. M., Núñez-Gastélum, J. A., Martínez, E. S. M., Ríos-Cortés, A. M., Garcia-Solis, M., & Pérez-Ishiwara, D. G. (2022). Characterization of Polyphenolic Compounds from Bacopa procumbens and Their Effects on Wound-Healing Process. Molecules, 27(19), 6521. https://doi.org/10.3390/molecules27196521