

Low Temperature Plasma Suppresses Lung Cancer Cells Growth via VEGF/VEGFR2/RAS/ERK Axis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
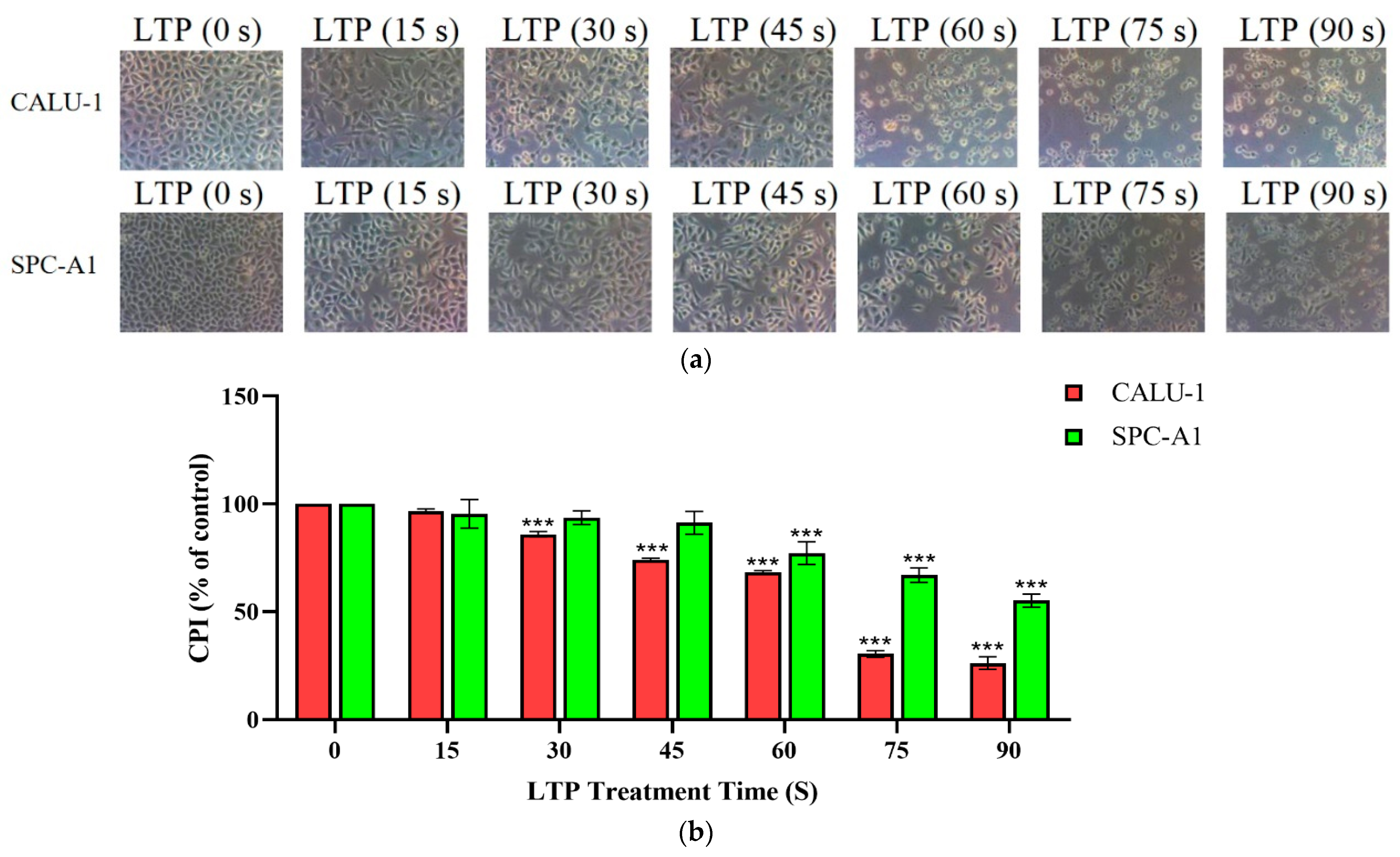
2.1. LTP Impaired Survival Abilities of CALU-1 and SPC-A1 Cells
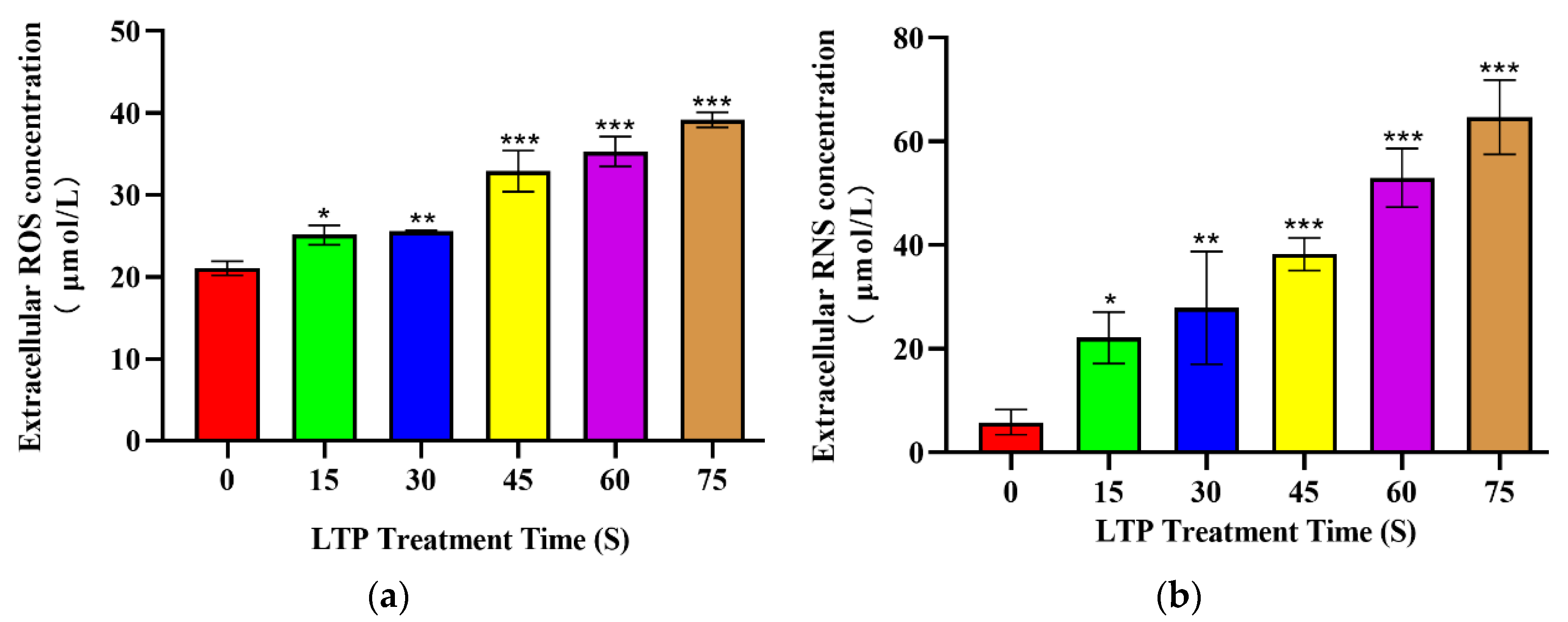
2.2. LTP Increased Extracellular ROS and RNS Levels
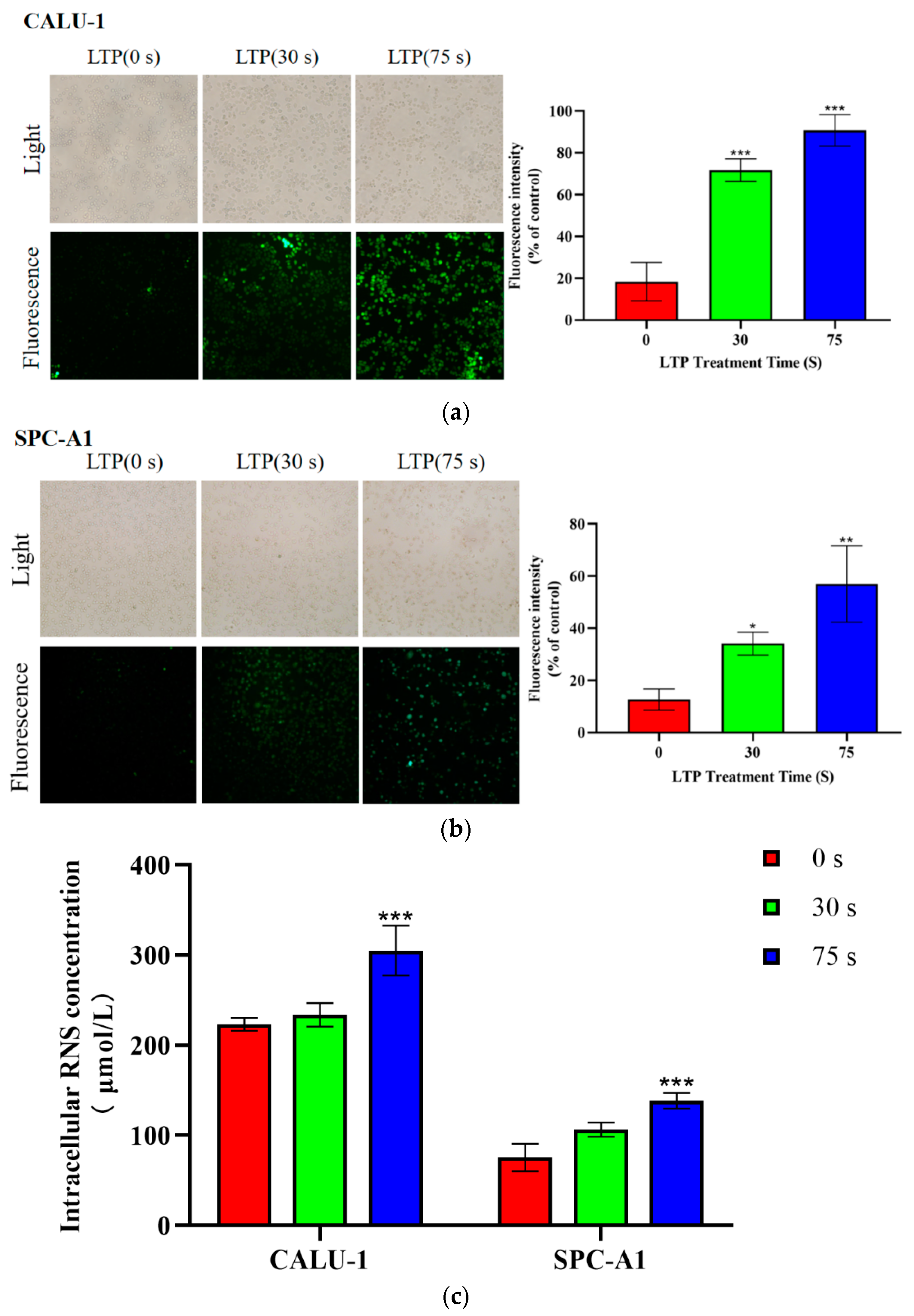
2.3. LTP Increased Intracellular ROS and RNS Generation
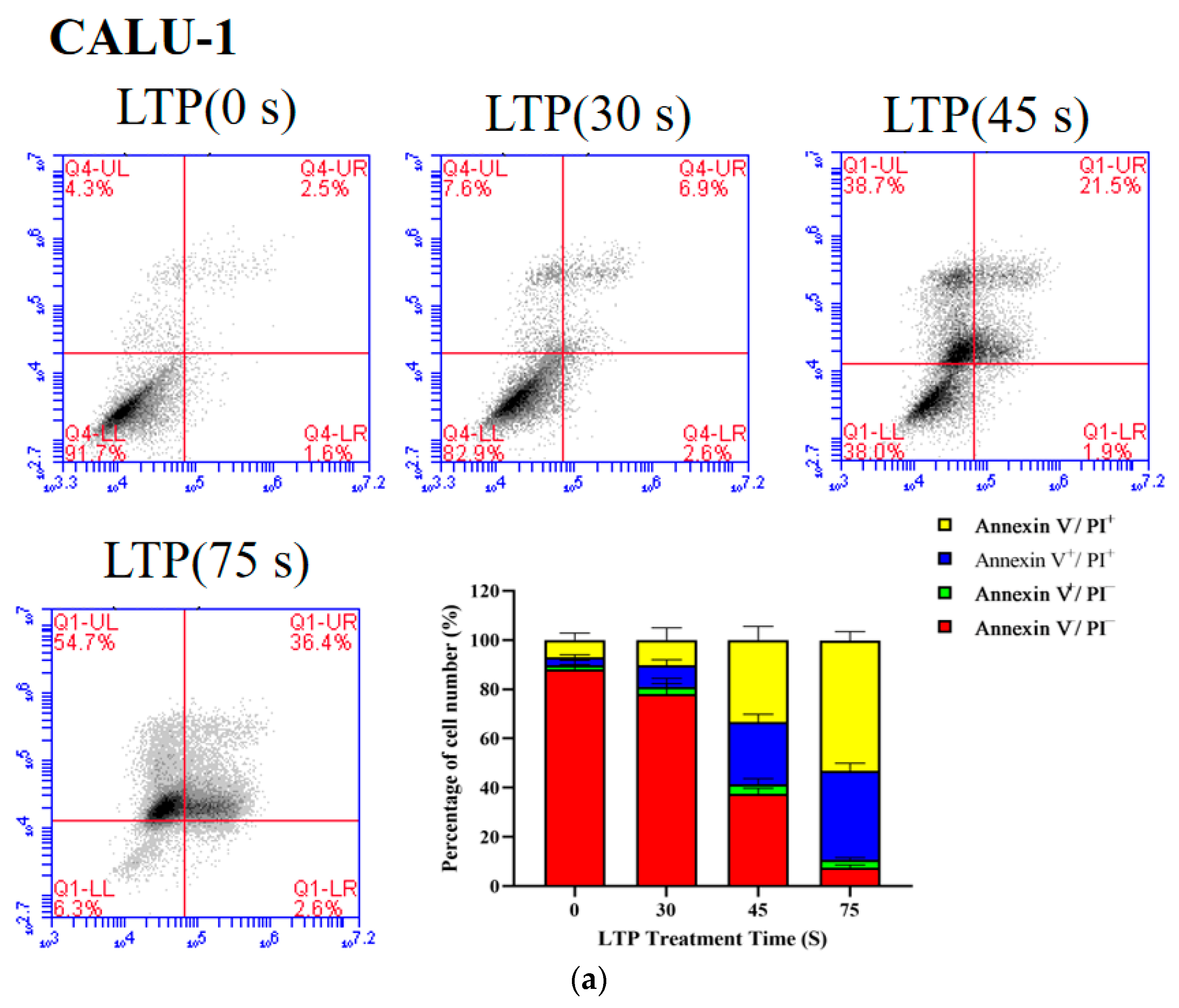
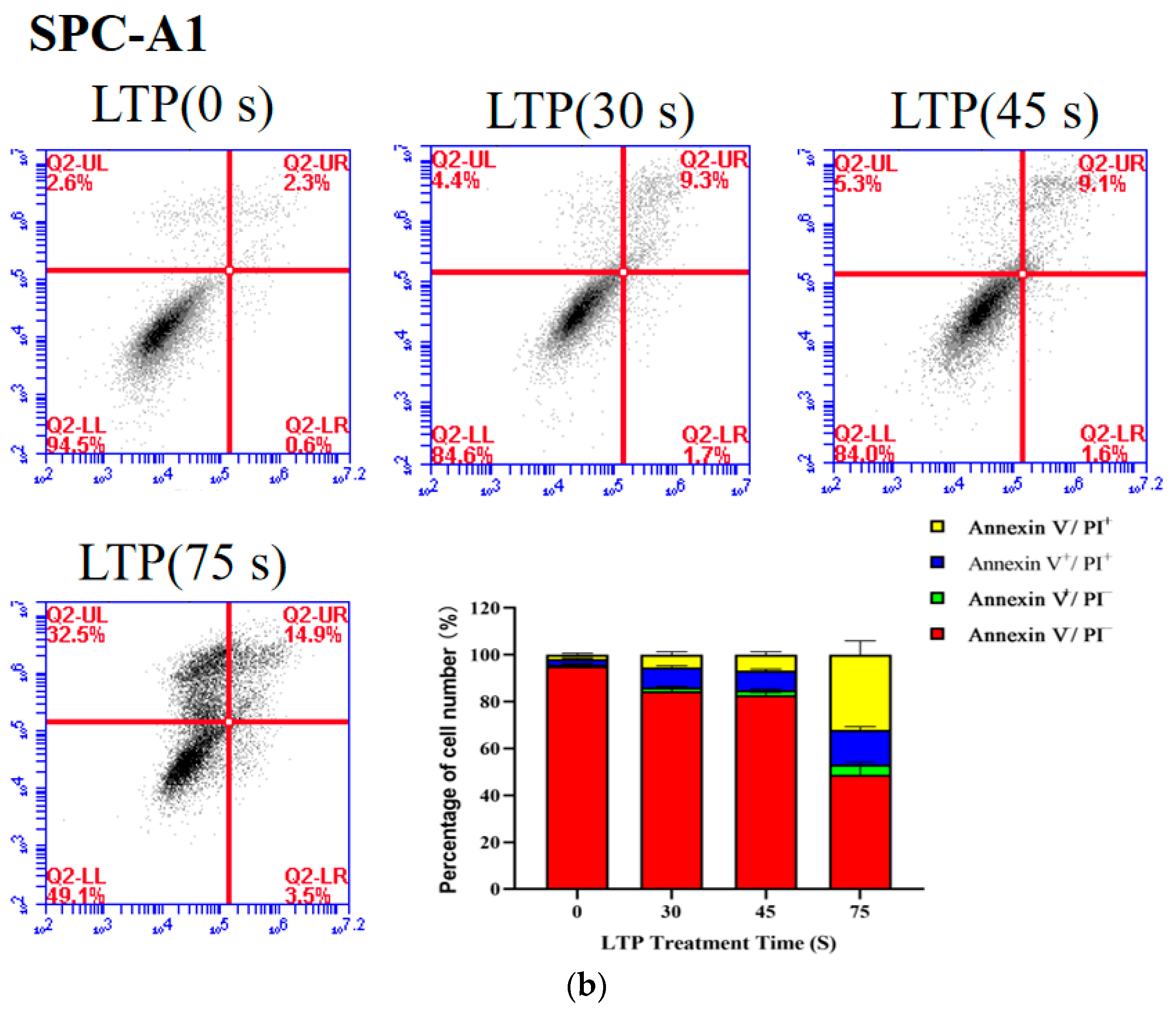
2.4. LTP Induced CALU-1 and SPC-A1 Cell Death
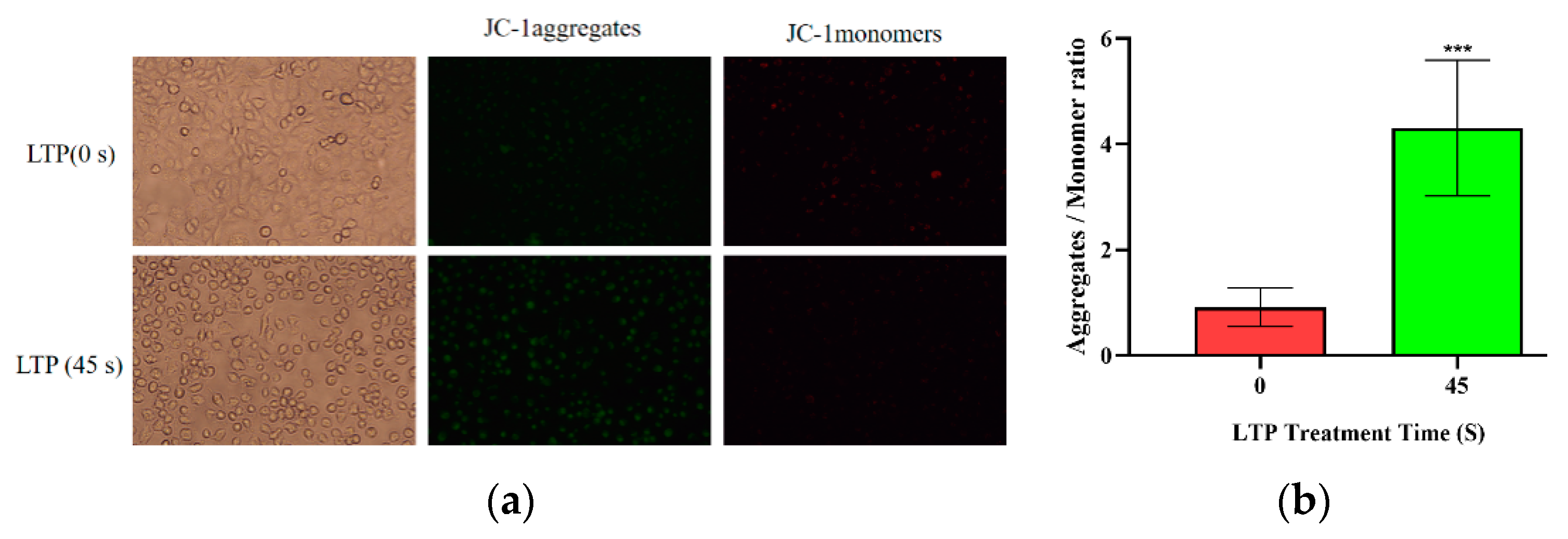
2.5. LTP Resulted in Depolarization of the Mitochondrial Membrane Potential of CALU-1 Cells
2.6. LTP Inhibited the Migration of CALU-1 Cells Migration
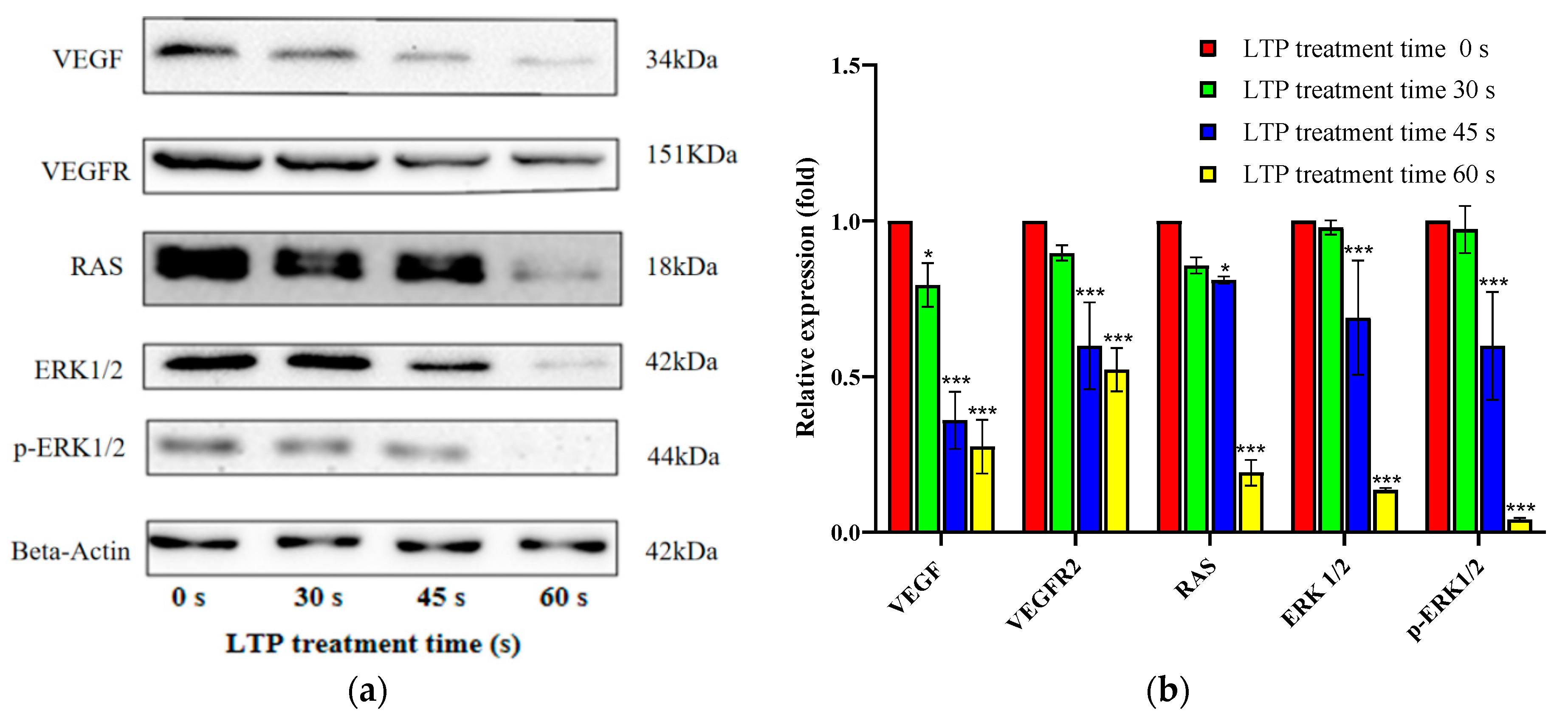
2.7. LTP Induced CALU-1 Cell Death via VEGF/VEGFR2/RAS/ERK Axis
3. Materials and Methods
3.1. NSCLC Cell Lines and Cultures
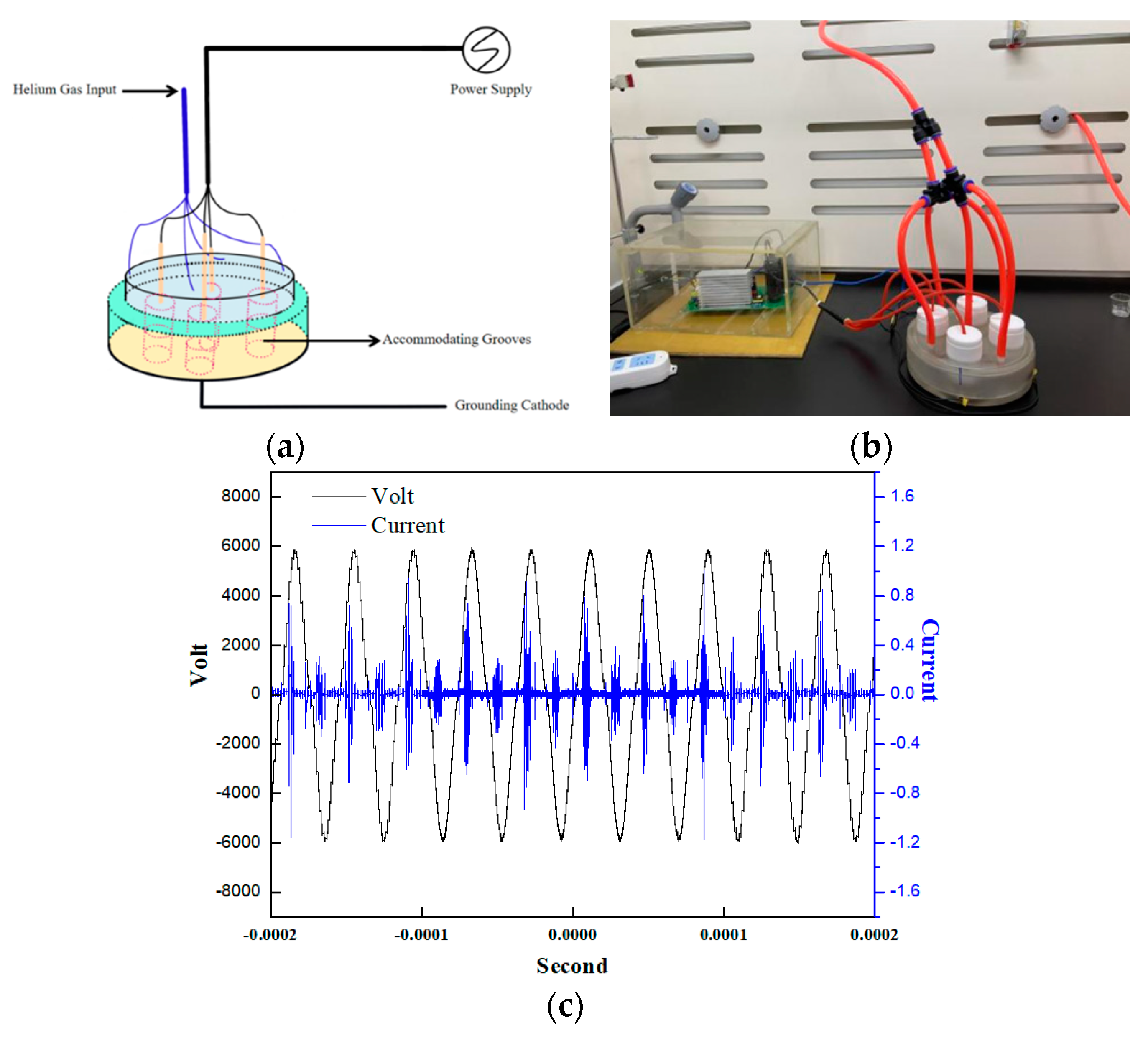
3.2. LTP Device
3.3. Cell Viability Assay
3.4. Extracellular Reactive Species Detection
3.5. Intracellular Reactive Species Detection
3.6. Flow Cytometer Analysis
3.7. JC-1 Detection
3.8. Scratch Assay
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, W.Q.; Zheng, R.S.; Baade, P.D.; Zhang, S.W.; Zeng, H.M.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer Statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, S.; Chiang, C.T.; Tsurutani, J.; Shiga, H.; Arwood, M.L.; Komiya, T.; Gills, J.J.; Memmott, R.M.; Dennis, P.A. Rapamycin downregulates thymidylate synthase and potentiates the activity of pemetrexed in non-small cell lung cancer. Oncotarget 2014, 5, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.R.; Yang, P.G.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Ahmed, M.; Liu, Z.J.; Afzal, K.S.; Weeks, D.; Lobo, S.M.; Kruskal, J.B.; Lenkinski, R.E.; Goldberg, S.N. Radiofrequency ablation: Effect of surrounding tissue composition on coagulation necrosis in a canine tumor model. Radiology 2004, 230, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Barlesi, F.; Pujol, J.L.; Daures, J.P. Should chemotherapy (Cx) for advanced non-small cell lung cancer (NSCLC) be platinum-based? A literature-based meta-analysis of randomized trials. J. Clin. Oncol. 2005, 23, 673. [Google Scholar] [CrossRef]
- Song, Y.H.; Chai, Q.; Wang, N.L.; Yang, F.F.; Wang, G.H.; Hu, J.Y. X-rays induced IL-8 production in lung cancer cells via p38/MAPK and NF-kappa B pathway. Int. J. Radiat. Biol. 2020, 96, 1374–1381. [Google Scholar] [CrossRef]
- Okumus, D.; Sarihan, S.; Gozcu, S.; Sigirli, D. The relationship between dosimetric factors, side effects, and survival in patients with non-small cell lung cancer treated with definitive radiotherapy. Med. Dosim. 2017, 42, 169–176. [Google Scholar] [CrossRef]
- Xiong, W.J.; Xu, Q.F.; Xu, Y.; Sun, C.J.; Li, N.; Zhou, L.; Liu, Y.M.; Zhou, X.J.; Wang, Y.S.; Wang, J.; et al. Stereotactic body radiation therapy for post-pulmonary lobectomy isolated lung metastasis of thoracic tumor: Survival and side effects. BMC Cancer 2014, 14, 719. [Google Scholar] [CrossRef]
- Ming, X.; Feng, Y.M.; Yang, C.W.; Wang, W.; Wang, P.; Deng, J. Radiation-induced heart disease in lung cancer radiotherapy: A dosimetric update. Medicine 2016, 95, e5051. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, H.; Hao, J.F.; Zhao, W.P.; Wu, Z.H.; Qiu, R.; Wang, X.H. Effect of p53 on lung carcinoma cells irradiated by carbon ions or X-rays. Nucl. Sci. Tech. 2009, 20, 146–151. [Google Scholar]
- Fridman, G.; Shereshevsky, A.; Jost, M.M.; Brooks, A.D.; Fridman, A.; Gutsol, A.; Vasilets, V.; Friedman, G. Floating Electrode Dielectric Barrier Discharge Plasma in Air Promoting Apoptotic Behavior in Melanoma Skin Cancer Cell Lines. Plasma Chem. Plasma Process. 2007, 27, 163–176. [Google Scholar] [CrossRef]
- Duarte, S.; Panariello, B.H. Comprehensive biomedical applications of low temperature plasmas. Arch. Biochem. Biophys. 2020, 693, 108560. [Google Scholar] [CrossRef]
- Keidar, M. Plasma for cancer treatment. Plasma Sources Sci. Technol. 2015, 24, 033001. [Google Scholar] [CrossRef]
- Von Woedtke, T.; Schmidt, A.; Bekeschus, S.; Wende, K.; Weltmann, K.D. Plasma Medicine: A Field of Applied Redox Biology. In Vivo 2019, 33, 1011–1026. [Google Scholar] [CrossRef]
- Girard, P.-M.; Arbabian, A.; Fleury, M.; Bauville, G.; Puech, V.; Dutreix, M.; Sousa, J.S. Synergistic Effect of H2O2 and NO2 in Cell Death Induced by Cold Atmospheric He Plasma. Sci. Rep. 2016, 6, 29098. [Google Scholar] [CrossRef] [PubMed]
- Kalghatgi, S.; Kelly, C.M.; Cerchar, E.; Torabi, B.; Alekseev, O.; Fridman, A.; Friedman, G.; Azizkhan-Clifford, J. Effects of Non-Thermal Plasma on Mammalian Cells. PLoS ONE 2011, 6, e16270. [Google Scholar] [CrossRef]
- Zhao, Z.W.; Zhang, D.K.; Wu, F.Z.; Tu, J.F.; Song, J.J.; Xu, M.; Ji, J.S. Sophoridine suppresses lenvatinib-resistant hepatocellular carcinoma growth by inhibiting RAS/MEK/ERK axis via decreasing VEGFR2 expression. J. Cell. Mol. Med. 2021, 25, 549–560. [Google Scholar] [CrossRef]
- Ramakrishnan, V.; Timm, M.; Haug, J.L.; Kimlinger, T.K.; Halling, T.; Wellik, L.E.; Witzig, T.E.; Rajkumar, S.V.; Adjei, A.A.; Kumar, S. Sorafenib, a multikinase inhibitor, is effective in vitro against non-hodgkin lymphoma and synergizes with the mTOR inhibitor rapamycin. Am. J. Hematol. 2012, 87, 277–283. [Google Scholar] [CrossRef]
- Song, Y.; Dai, F.; Zhai, D.; Dong, Y.; Zhang, J.; Lu, B.; Luo, J.; Liu, M.; Yi, Z. Usnic acid inhibits breast tumor angiogenesis and growth by suppressing VEGFR2-mediated AKT and ERK1/2 signaling pathways. Angiogenesis 2012, 15, 421–432. [Google Scholar] [CrossRef]
- Huang, M.; Huang, B.; Li, G.; Zeng, S. Apatinib affect VEGF-mediated cell proliferation, migration, invasion via blocking VEGFR2/RAF/MEK/ERK and PI3K/AKT pathways in cholangiocarcinoma cell. BMC Gastroenterol. 2018, 18, 169. [Google Scholar] [CrossRef]
- Yue, F.; Zhou, M.Y.; Sun, F.; Kong, Z.; Wang, J.; Sun, Z.Q.; Hu, L.J.; Wang, J.L.; Hua, Q.; Yu, J.P. Apatinib mesylate regulates the Ras/Raf/MEK/ERK and JAK2/STAT3 signaling pathways to affect the proliferation, migration and apoptosis of esophageal cancer cells and the growth of transplanted tumors in tumor-bearing mice. Chin. J. Clin. Oncol. 2019, 41, 263–275. [Google Scholar]
- Amiri, P.; Aikawa, M.E.; Dove, J.; Stuart, D.D.; Poon, D.; Pick, T.; Ramurthy, S.; Subramanian, S.; Levine, B.; Costales, A.; et al. CHIR-265 is a potent selective inhibitor of c-Raf/B-Raf/B-mut-Raf that effectively inhibits proliferation and survival of cancer cell lines with Ras/Raf pathway mutations. Cancer Res. 2006, 66, 4855. [Google Scholar]
- Belenchia, R.; Broggi, M.; Georgelos, K.; Mcnabola, A.; Novicki, E.N.; Rowley, R.B.; Trombley, S.; Wilkie, D.; Wilhelm, S.; Taylor, I. Baseline phosphorylated ERK levels in renal cell carcinoma patients from a phase II study of BAY 43-9006. Cancer Res. 2004, 64, 848. [Google Scholar]
- Chatterjee, S.; Rhee, Y.; Chung, P.S.; Ge, R.F.; Ahn, J.C. Sulforaphene enhances the efficacy of photodynamic therapy in anaplastic thyroid cancer through Ras/RAF/MEK/ERK pathway suppression. J. Photochem. Photobiol. B. 2018, 179, 46–53. [Google Scholar] [CrossRef]
- Iseki, S.; Nakamura, K.; Hayashi, M.; Tanaka, H.; Kondo, H.; Kajiyama, H.; Kano, H.; Kikkawa, F.; Hori, M. Selective killing of ovarian cancer cells through induction of apoptosis by nonequilibrium atmospheric pressure plasma. Appl. Phys. Lett. 2012, 100, 113702. [Google Scholar] [CrossRef]
- Guerrero-Preston, R.; Ogawa, T.; Uemura, M.; Shumulinsky, G.; Valle, B.L.; Pirini, F.; Ravi, R.; Sidransky, D.; Keidar, M.; Trink, B. Cold atmospheric plasma treatment selectively targets head and neck squamous cell carcinoma cells. Int. J. Mol. Med. 2014, 34, 941–946. [Google Scholar] [CrossRef]
- Semmler, M.L.; Bekeschus, S.; Schaefer, M.; Bernhardt, T.; Fischer, T.; Witzke, K.; Seebauer, C.; Rebl, H.; Grambow, E.; Vollmar, B.; et al. Molecular Mechanisms of the Efficacy of Cold Atmospheric Pressure Plasma (CAP) in Cancer Treatment. Cancers 2020, 12, 269. [Google Scholar] [CrossRef]
- Wang, Q.; Zheng, X.L.; Yang, L.; Shi, F.; Gao, L.B.; Zhong, Y.J.; Sun, H.; He, F.; Lin, Y.; Wang, X. Reactive oxygen species-mediated apoptosis contributes to chemosensitization effect of saikosaponins on cisplatin-induced cytotoxicity in cancer cells. J. Exp. Clin. Cancer Res. 2010, 29, 159. [Google Scholar] [CrossRef]
- Chen, G.; Chen, Z.; Wen, D.; Wang, Z.; Li, H.; Zeng, Y.; Dotti, G.; Wirz, R.E.; Gu, Z. Transdermal cold atmospheric plasma-mediated immune checkpoint blockade therapy. Proc. Natl. Acad. Sci. USA 2020, 117, 3687–3692. [Google Scholar] [CrossRef]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Karki, S.B.; Gupta, T.T.; Yildirim-Ayan, E.; Eisenmann, K.M.; Ayan, H. Miniature Non-thermal Plasma Induced Cell Cycle Arrest and Apoptosis in Lung Carcinoma Cells. Plasma Chem. Plasma Process. 2019, 40, 99–117. [Google Scholar] [CrossRef]
- Yan, D.; Wang, Q.; Malyavko, A.; Zolotukhin, D.B.; Adhikari, M.; Sherman, J.H.; Keidar, M. The anti-glioblastoma effect of cold atmospheric plasma treatment: Physical pathway v.s. chemical pathway. Sci. Rep. 2020, 10, 11788. [Google Scholar] [CrossRef] [PubMed]
- Chernets, N.; Kurpad, D.S.; Alexeev, V.; Rodrigues, D.B.; Freeman, T.A. Reaction Chemistry Generated by Nanosecond Pulsed Dielectric Barrier Discharge Treatment is Responsible for the Tumor Eradication in the B16 Melanoma Mouse Model. Plasma Process. Polym. 2015, 12, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, N.K.; Kaushik, N.; Yoo, K.C.; Uddin, N.; Kim, J.S.; Lee, S.J.; Choi, E.H. Data on combination effect of PEG-coated gold nanoparticles and non-thermal plasma inhibit growth of solid tumors. Data Brief 2016, 9, 318–323. [Google Scholar] [CrossRef]
- Kim, S.J.; Joh, H.M.; Chung, T.H. Production of intracellular reactive oxygen species and change of cell viability induced by atmospheric pressure plasma in normal and cancer cells. Appl. Phys. Lett. 2013, 103, 153705. [Google Scholar] [CrossRef]
- Zuo, X.; Wei, Y.; Chen, L.W.; Meng, Y.D. Non-equilibrium atmospheric pressure microplasma jet: An approach to endoscopic therapies. Phys. Plasmas 2013, 20, 5. [Google Scholar] [CrossRef]
- Lobanova, E.G.; Kondrat’eva, E.V. Measurement of mitochondrial membrane potential in leukocyte suspension by fluorescent spectroscopy. Bull. Exp. Biol. Med. 2014, 157, 288–290. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef]
- Wu, S.F.; Noren Hooten, N.; Freeman, D.W.; Mode, N.A.; Zonderman, A.B.; Evans, M.K. Extracellular vesicles in diabetes mellitus induce alterations in endothelial cell morphology and migration. J. Transl. Med. 2020, 18, 230. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, J.; Jiang, D.; Wuren, T.; Ma, S.; Du, Y.; Yi, X.; Wu, S. The plasma level changes of VEGF and soluble VEGF receptor-1 are associated with high-altitude pulmonary edema. J. Med Investig. 2018, 65, 64–68. [Google Scholar] [CrossRef]
- Hu, C.X.; Zhuang, W.; Qiao, Y.; Liu, B.; Liu, L.; Hui, K.Y.; Jiang, X.D. Effects of combined inhibition of STAT3 and VEGFR2 pathways on the radiosensitivity of non-small-cell lung cancer cells. OncoTargets Ther. 2019, 12, 933–944. [Google Scholar] [CrossRef]
- Watanabe, H.; Ichihara, E.; Kayatani, H.; Makimoto, G.; Ninomiya, K.; Nishii, K.; Higo, H.; Ando, C.; Okawa, S.; Nakasuka, T. VEGFR2 blockade augments the effects of tyrosine kinase inhibitors by inhibiting angiogenesis and oncogenic signaling in oncogene-driven non-small-cell lung cancers. Cancer Sci. 2021, 112, 1853–1864. [Google Scholar] [CrossRef]
- Fey, D.; Matallanas, D.; Rauch, J.; Rukhlenko, O.S.; Kholodenko, B.N. The complexities and versatility of the RAS-to-ERK signalling system in normal and cancer cells. Semin. Cell Dev. Biol. 2016, 58, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Hilger, R.A.; Scheulen, M.E.; Strumberg, D. The Ras-Raf-MEK-ERK pathway in the treatment of cancer. Onkologie 2002, 25, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Friday, B.B.; Adjei, A.A. Advances in targeting the Ras/Raf/MEK/Erk mitogen-activated protein kinase cascade with MEK inhibitors for cancer therapy. Clin. Cancer Res. 2008, 14, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Li, G.; Liu, J.; Xu, D.; Shi, X.; Zhang, G. Inhibitory effect of non-thermal plasma synergistic Tegafur on pancreatic tumor cell line BxPc-3 proliferation. Plasma Process. Polym. 2019, 16, e1800165. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhang, Y.; Bao, J.; Chen, J.; Song, W. Low Temperature Plasma Suppresses Lung Cancer Cells Growth via VEGF/VEGFR2/RAS/ERK Axis. Molecules 2022, 27, 5934. https://doi.org/10.3390/molecules27185934
Zhou Y, Zhang Y, Bao J, Chen J, Song W. Low Temperature Plasma Suppresses Lung Cancer Cells Growth via VEGF/VEGFR2/RAS/ERK Axis. Molecules. 2022; 27(18):5934. https://doi.org/10.3390/molecules27185934
Chicago/Turabian StyleZhou, Yuanyuan, Yan Zhang, Jie Bao, Jinwu Chen, and Wencheng Song. 2022. "Low Temperature Plasma Suppresses Lung Cancer Cells Growth via VEGF/VEGFR2/RAS/ERK Axis" Molecules 27, no. 18: 5934. https://doi.org/10.3390/molecules27185934
APA StyleZhou, Y., Zhang, Y., Bao, J., Chen, J., & Song, W. (2022). Low Temperature Plasma Suppresses Lung Cancer Cells Growth via VEGF/VEGFR2/RAS/ERK Axis. Molecules, 27(18), 5934. https://doi.org/10.3390/molecules27185934