
Comparative Proteomic Analysis of Potato Roots from Resistant and Susceptible Cultivars to Spongospora subterranea Zoospore Root Attachment In Vitro

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
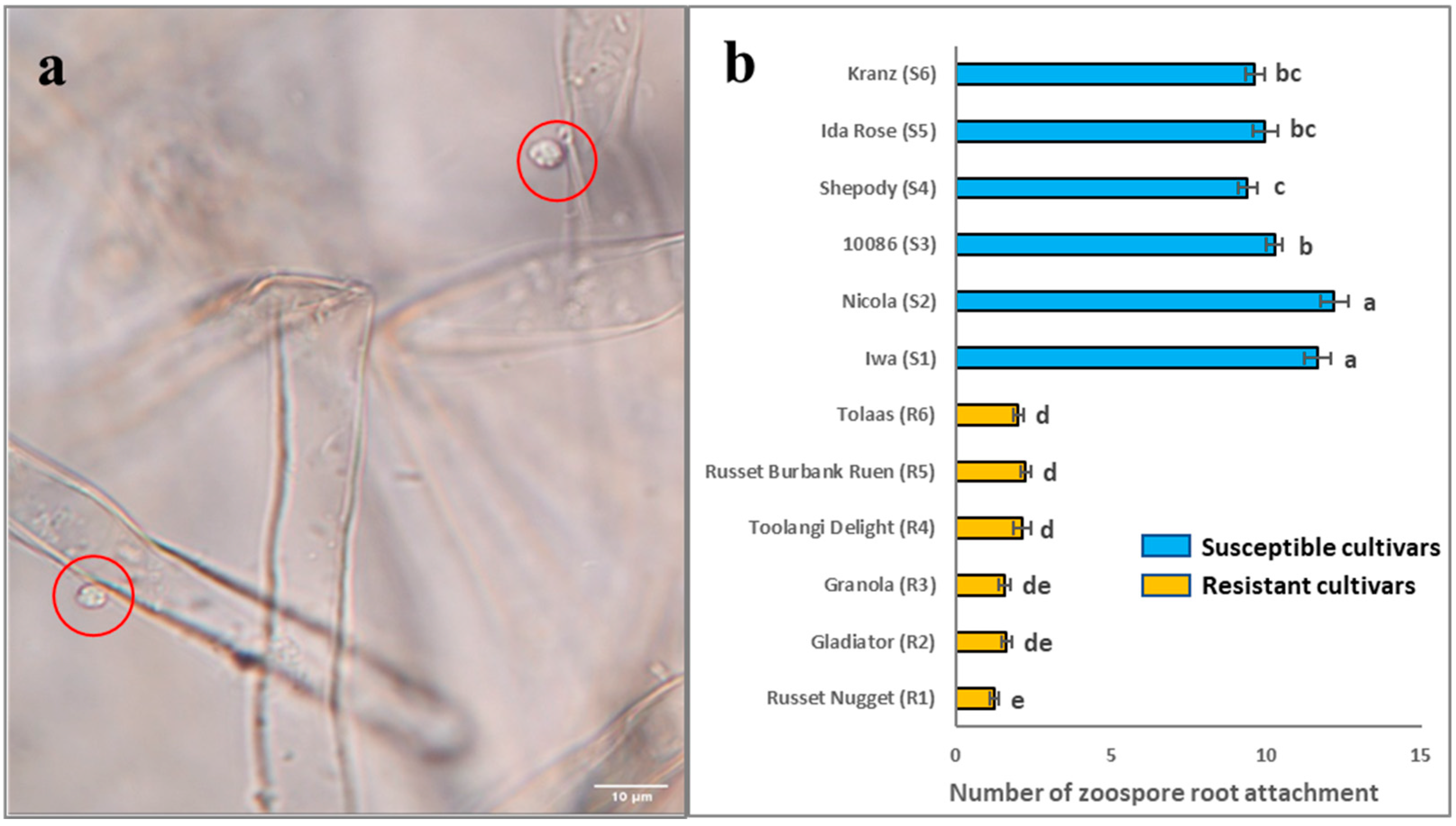
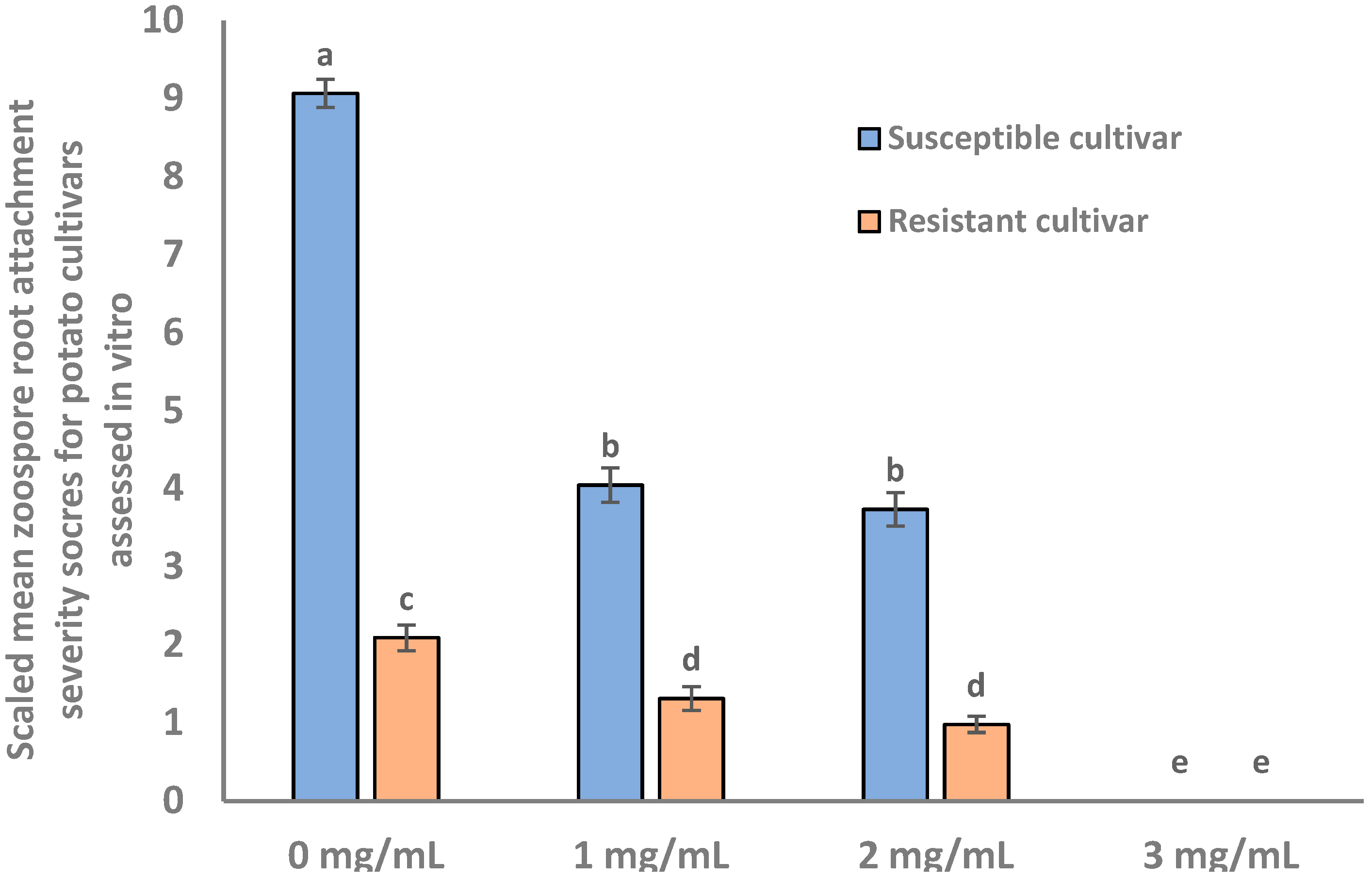
2.1. Root Attachment of Different Potato Cultivars Subjected to S. subterranea Infection
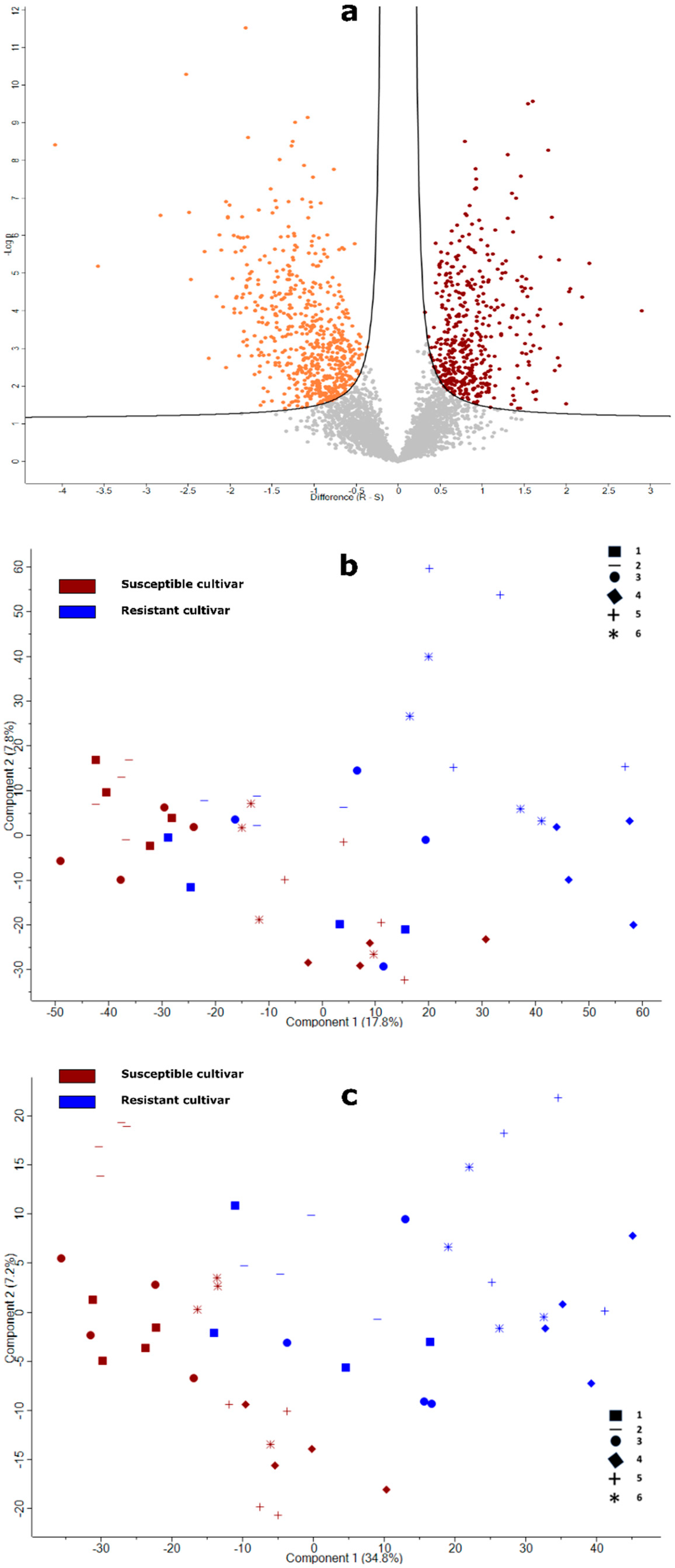
2.2. Overview of the Proteins in Potato Roots Identified by Label-Free Quantitative Proteomics
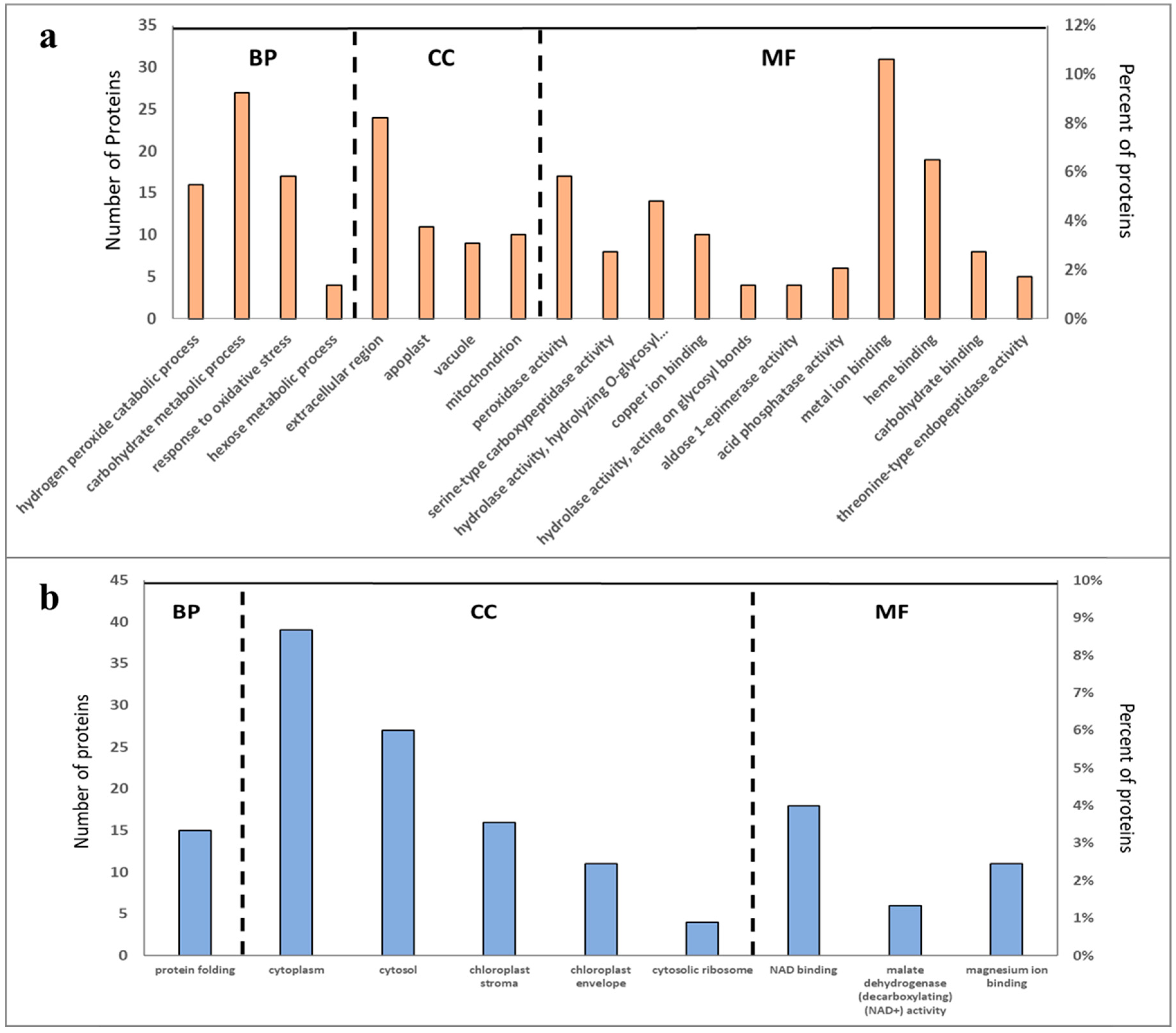
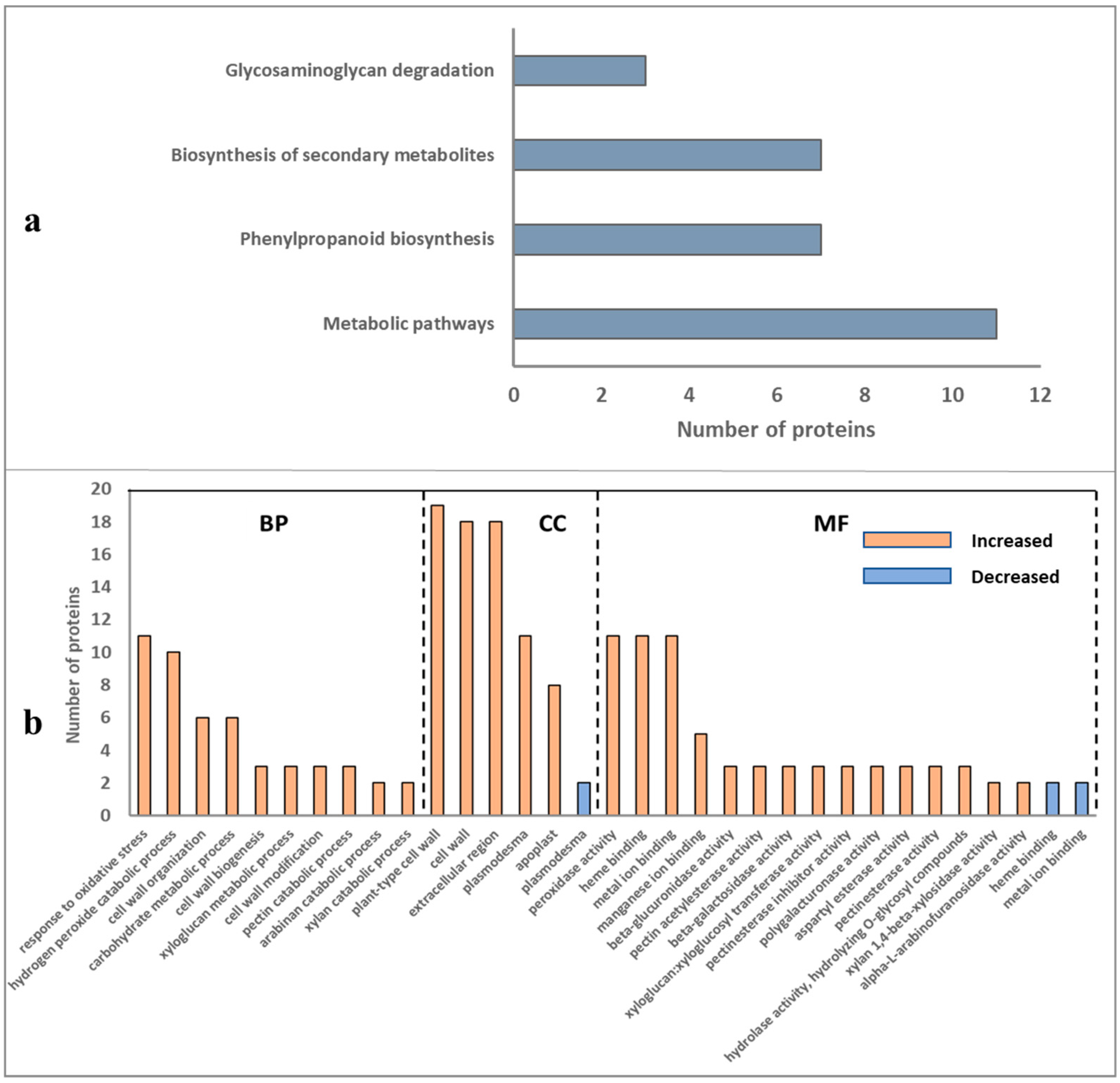
2.3. Overall Functional Classification of Differentially Abundant Proteins
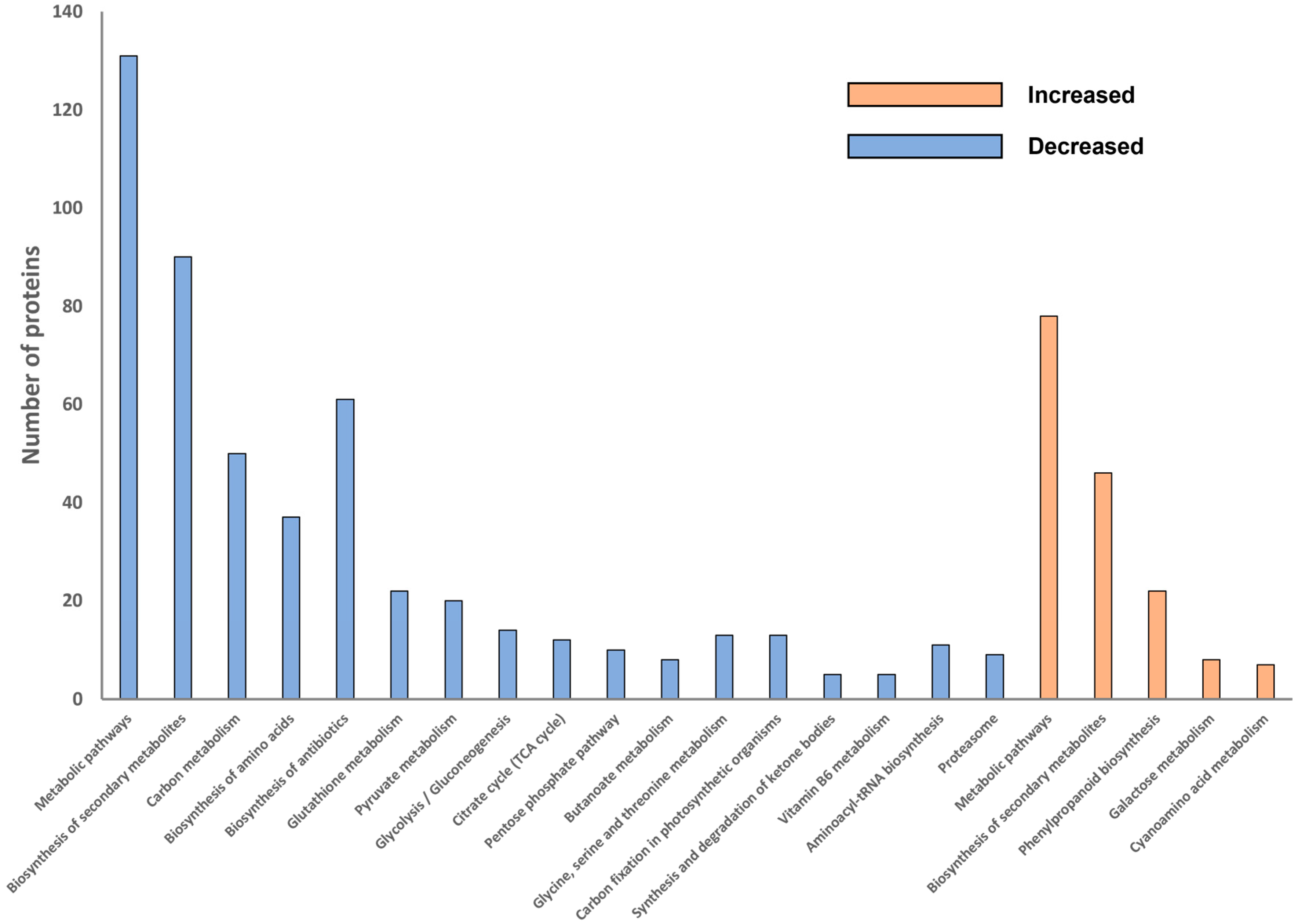
2.4. Overall Pathway Analysis of Differentially Abundant Proteins
2.5. Differentially Abundant Proteins of Root Cell Wall and Pathway Analysis
2.6. Effects of Pectinase Treatment of Potato Roots on Zoospore Root Attachment
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Spongospora subterranea Inoculum Preparation and Zoospore Germination
4.3. Zoospore Root Attachment Assay
4.4. Protein Extraction and Peptide Sample Preparation
4.5. Sample Proteomic Analysis, Data Processing and Analysis
4.6. Pectinase Treatment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Harrison, J.G.; Searle, R.J.; Williams, N.A. Powdery scab disease of potato—A review. Plant Pathol. 1997, 46, 1–25. [Google Scholar] [CrossRef]
- Falloon, R.E.; Genet, R.A.; Wallace, A.R.; Butler, R.C. Susceptibility of potato (Solanum tuberosum) cultivars to powdery scab (caused by Spongospora subterranea f. sp. subterranea), and relationships between tuber and root infection. Australas. Plant Pathol. 2003, 32, 377–385. [Google Scholar] [CrossRef]
- Merz, U. Powdery scab of potato—Occurrence, life cycle and epidemiology. Am. J. Potato Res. 2008, 85, 241–246. [Google Scholar] [CrossRef]
- Merz, U.; Falloon, R.E. Review: Powdery scab of potato-increased knowledge of pathogen biology and disease epidemiology for effective disease management. Potato Res. 2009, 52, 17–37. [Google Scholar] [CrossRef]
- Tegg, R.S.; Thangavel, T.; Balendres, M.A.; Wilson, C.R. Grading Seed Potato Lots to Remove Tubers with Powdery Scab Damage may not Eliminate the Pathogen Threat. Am. J. Potato Res. 2016, 93, 231–238. [Google Scholar] [CrossRef]
- Wilson, C.R. Plant pathogens—The great thieves of vegetable value. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes, Brisbane, Australia, 25 November 2016; Volume 1123, pp. 7–15. [Google Scholar]
- Balendres, M.A.; Tegg, R.S.; Wilson, C.R. Key events in pathogenesis of Spongospora diseases in potato: A review. Australas. Plant Pathol. 2016, 45, 229–240. [Google Scholar] [CrossRef]
- Braithwaite, M.; Falloon, R.E.; Genet, R.A.; Wallace, A.R.; Fletcher, J.D.; Braam, W.F. Control of powdery scab of potatoes with chemical seed tuber treatments. N. Z. J. Crop Hortic. Sci. 1994, 22, 121–128. [Google Scholar] [CrossRef]
- Falloon, R.E.; Wallace, A.R.; Braithwaite, M.; Genet, R.A.; Nott, H.M.; Fletcher, J.D.; Braam, W.F. Assessment of seed tuber, in-furrow, and foliar chemical treatments for control of powdery scab (Spongospora subterranea f. sp. subterranea) of potato. N. Z. J. Crop Hortic. Sci. 1996, 24, 341–353. [Google Scholar] [CrossRef]
- Tuncer, G. The effect of irrigation and nitrogen on powdery scab and yield of potatoes. Potato Res. 2002, 45, 153–161. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Shah, F.A.; Falloon, R.E.; Butler, R.C.; Lister, R.A. Low amounts of Spongospora subterranea sporosorus inoculum cause severe powdery scab, root galling and reduced water use in potato (Solanum tuberosum). Australas. Plant Pathol. 2012, 41, 219–228. [Google Scholar] [CrossRef]
- Falloon, R.E. Control of powdery scab of potato: Towards integrated disease management. Am. J. Potato Res. 2008, 85, 394. [Google Scholar] [CrossRef]
- Balotf, S.; Wilson, R.; Tegg, R.S.; Nichols, D.S.; Wilson, C.R. In planta transcriptome and proteome profiles of Spongospora subterranea in resistant and susceptible host environments illuminates regulatory principles underlying host-pathogen interaction. Biology 2021, 10, 840. [Google Scholar] [CrossRef] [PubMed]
- Balotf, S.; Wilson, R.; Tegg, R.S.; Nichols, D.S.; Wilson, C.R. Shotgun proteomics as a powerful tool for the study of the proteomes of plants, their pathogens, and plant and pathogen interactions. Proteomes 2022, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Gupta, R.; Krishna, R.; Kim, S.T.; Lee, D.Y.; Hwang, D.-J.; Bae, S.-C.; Ahn, I.-P. Proteome Analysis of Disease Resistance against Ralstonia solanacearum in Potato Cultivar CT206-10. Plant Pathol. J. 2016, 32, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Gao, J.; Zhang, Y.; Wang, Z.; Zhang, D.; Chen, Q.; Ye, X.; Xu, Y.; Yang, G.; Yan, L.; et al. Quantitative Proteomics of Potato Leaves Infected with Phytophthora infestans Provides Insights into Coordinated and Altered Protein Expression during Early and Late Disease Stages. Int. J. Mol. Sci. 2019, 20, 136. [Google Scholar] [CrossRef]
- Szajko, K.; Plich, J.; Przetakiewicz, J.; Sołtys-Kalina, D.; Marczewski, W. Comparative proteomic analysis of resistant and susceptible potato cultivars during Synchytrium endobioticum infestation. Planta 2020, 251, 4. [Google Scholar] [CrossRef]
- Yu, X.; Tegg, R.S.; Eyles, A.; Wilson, A.J.; Wilson, C.R. Development and validation of a novel rapid in-vitro assay for resistance of potato cultivars to Spongospora subterranea zoospore root attachment. Plant Pathol. 2022. Submitted and in revision. [Google Scholar]
- Ledingham, G.A. Occurrence of zoosporangia in Spongospora subterranea, (wallroth) lagerheim. Nature 1935, 135, 394. [Google Scholar] [CrossRef]
- Braselton, J.P. Ultrastructural karyology of Spongospora subterranea (Plasmodiophoromycetes). Can. J. Bot. 1992, 70, 1228–1233. [Google Scholar] [CrossRef]
- Mehdy, M.C. Active oxygen species in plant defense against pathogens. Plant Physiol. 1994, 105, 467–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Tian, Z.D.; Liu, J.; Xie, C.H. A novel leucine-rich repeat receptor-like kinase gene in potato, StLRPK1, is involved in response to diverse stresses. Mol. Biol. Rep. 2009, 36, 2365–2374. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.S.; Shortt, B.J.; Lawrence, E.B.; Leon, J.; Fitzsimmons, K.C.; Levine, E.B.; Raskin, I.; Shah, D.M. Activation of host defense mechanisms by elevated production of H2O2 in transgenic plants. Plant Physiol. 1997, 115, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Feucht, W. Tissue-specific oxidative browning of polyphenols by peroxidase in cherry shoots. Gartenbauwissenschaft 1980, 45, 68–73. [Google Scholar]
- Boordman, N.; Conn, E.; Hatch, M.; Stumpf, P. The Biochemistry of Plants: A Comprehensive Treatise; Academic Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum Lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Goldberg, R.; Imberty, A.; Liberman, M.; Prat, R. Relationships between peroxidatic activities and cell wall plasticity. In Molecular and Physiological Aspects of Plant Peroxidases; Greppin, H., Penel, C., Gaspar, T., Eds.; University M. Curie-Sklowdowska: Lublin, Poland, 1986. [Google Scholar]
- Beffa, R.; Martin, H.V.; Pilet, P.-E. In vitro oxidation of indoleacetic acid by soluble auxin-oxidases and peroxidases from maize roots. Plant Physiol. 1990, 94, 485–491. [Google Scholar] [CrossRef]
- Young, S.A.; Guo, A.; Guikema, J.A.; White, F.F.; Leach, J.E. Rice cationic peroxidase accumulates in xylem vessels during incompatible interactions with Xanthomonas oryzae pv oryzae. Plant Physiol. 1995, 107, 1333–1341. [Google Scholar] [CrossRef]
- Saikia, R.; Kumar, R.; Singh, T.; Srivastava, A.K.; Arora, D.K.; Lee, M.-W. Induction of defense related enzymes and pathogenesis related proteins in Pseudomonas fluorescens-treated chickpea in response to infection by Fusarium oxysporum f. sp. ciceri. Mycobiology 2004, 32, 47–53. [Google Scholar] [CrossRef]
- Lim, S.; Chisholm, K.; Coffin, R.H.; Peters, R.D.; Al-Mughrabi, K.I.; Wang-Pruski, G.; Pinto, D.M. Protein profiling in potato (Solanum tuberosum L.) leaf tissues by differential centrifugation. J. Proteome Res. 2012, 11, 2594–2601. [Google Scholar] [CrossRef]
- Zhao, Q.; Dixon, R.A. Altering the cell wall and its impact on plant disease: From forage to bioenergy. Annu. Rev. Phytopathol. 2014, 52, 69–91. [Google Scholar] [CrossRef]
- Li, P.; Fan, R.; Peng, Z.; Qing, Y.; Fang, Z. Transcriptome analysis of resistance mechanism to potato wart disease. Open Life Sci. 2021, 16, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Song, T.; Chu, M.; Yu, F.; Kumar, S.; Karunakaran, C.; Peng, G. Evaluating changes in cell-wall components associated with clubroot resistance using fourier transform infrared spectroscopy and RT-PCR. Int. J. Mol. Sci. 2017, 18, 2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciaghi, S.; Schwelm, A.; Neuhauser, S. Transcriptomic response in symptomless roots of clubroot infected kohlrabi (Brassica oleracea var. gongylodes) mirrors resistant plants. BMC Plant Biol. 2019, 19, 288. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, S.A.; Strelkov, S.E.; Links, M.G.; Clarke, W.E.; Robinson, S.J.; Djavaheri, M.; Malinowski, R.; Haddadi, P.; Kagale, S.; Parkin, I.A.P.; et al. The compact genome of the plant pathogen Plasmodiophora brassicae is adapted to intracellular interactions with host Brassica spp. BMC Genom. 2016, 17, 272. [Google Scholar] [CrossRef]
- Balotf, S.; Wilson, R.; Nichols, D.; Tegg, R.; Wilson, C. Multi-omics reveals mechanisms of resistance to potato root infection by Spongospora subterranea. Sci. Rep. 2022, 12, 10804. [Google Scholar] [CrossRef]
- Gou, J.-Y.; Miller, L.M.; Hou, G.; Yu, X.-H.; Chen, X.-Y.; Liu, C.-J. Acetylesterase-mediated deacetylation of pectin impairs cell elongation, pollen germination, and plant reproduction. Plant Cell 2012, 24, 50–65. [Google Scholar] [CrossRef]
- Maldonado, M.; de Saad, A.S. Production of pectinesterase and polygalacturonase by Aspergillus niger in submerged and solid state systems. J. Ind. Microbiol. Biotechnol. 1998, 20, 34–38. [Google Scholar] [CrossRef]
- Byrt, P.N.; Irving, H.R.; Grant, B.R. The effect of organic compounds on the encystment, viability and germination of zoospores of Phytophthora cinnamomi. J. Gen. Microbiol. 1982, 128, 2343–2351. [Google Scholar] [CrossRef]
- Irving, H.R.; Grant, B.R. The effects of pectin and plant root surface carbohydrates on encystment and development of Phytophthora cinnamomi zoospores. J. Gen. Microbiol. 1984, 130, 1015–1018. [Google Scholar] [CrossRef]
- Grant, B.R.; Irving, H.R.; Radda, M. The effect of pectin and related-compounds on encystment and germination of Phytophthora palmivora zoospores. J. Gen. Microbiol. 1985, 131, 669–676. [Google Scholar] [CrossRef]
- Hughes, C.S.; Moggridge, S.; Muller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Balotf, S.; Wilson, R.; Tegg, R.S.; Nichols, D.S.; Wilson, C.R. Quantitative proteomics provides an insight into germination-related proteins in the obligate biotrophic plant pathogen Spongospora subterranea. Environ. Microbiol. Rep. 2021, 13, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Dey, S.; Uddin, M.; Barua, R.; Hossain, M. Extracellular pectinase from a novel bacterium Chryseobacterium indologenes strain SD and its application in fruit juice clarification. Enzym. Res. 2018, 2018, 3859752. [Google Scholar] [CrossRef] [PubMed]
- Roosdiana, A.; Prasetyawan, S.; Mahdi, C.; Sutrisno, S. Production and characterization of Bacillus firmus pectinase. J. Pure Appl. Chem. Res. 2013, 2, 35–41. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein Description | Fold Change | Adjusted p-Value |
|---|---|---|---|
| M1C976 | Peroxidase | 1.9 | 0.00 |
| M1B051 | Germin-like protein | 1.7 | 0.00 |
| M1BUZ0 | Germin-like protein | 1.5 | 0.00 |
| M1A147 | Beta-galactosidase | 1.5 | 0.00 |
| M1B041 | Germin-like protein | 1.1 | 0.00 |
| M1BJ45 | Pectinesterase | 1.1 | 0.01 |
| M1B6G3 | Peroxidase | 1.1 | 0.00 |
| M1AQZ8 | Xyloglucan | 1.1 | 0.02 |
| M1BFU7 | Germin-like protein | 1.1 | 0.00 |
| M1D0Z2 | Heparanase | 1.0 | 0.00 |
| M1BRR7 | Pectin acetylesterase | 1.0 | 0.00 |
| M1AWV7 | Polygalacturonase | 1.0 | 0.00 |
| M1C8D8 | Pectin acetylesterase | 1.0 | 0.00 |
| M0ZQ51 | Xyloglucan endotransglucosylase/hydrolase | 0.9 | 0.03 |
| M0ZJ69 | Peroxidase | 0.9 | 0.01 |
| M1A385 | Pectin acetylesterase | 0.9 | 0.00 |
| M1AZG9 | Glycoside hydrolase family 28 protein | 0.9 | 0.00 |
| M1DTA0 | Pectinesterase | 0.9 | 0.02 |
| M1BUZ2 | Germin-like protein | 0.9 | 0.00 |
| M1AIV9 | Pectinesterase | 0.9 | 0.00 |
| M1B6G2 | Peroxidase | 0.9 | 0.01 |
| M1CV50 | Expansin | 0.9 | 0.01 |
| M1CI69 | Beta-galactosidase | 0.8 | 0.00 |
| M0ZGW4 | Polygalacturonase | 0.8 | 0.05 |
| M1BTK5 | Peroxidase | 0.8 | 0.01 |
| M1CE55 | Peroxidase | 0.8 | 0.01 |
| M1A2Z2 | Peroxidase | 0.8 | 0.00 |
| M1BYZ4 | Peroxidase | 0.8 | 0.01 |
| M1AKA7 | Periplasmic beta-glucosidase | 0.8 | 0.01 |
| M0ZJ70 | Peroxidase | 0.8 | 0.03 |
| M1ARG0 | Heparanase | 0.7 | 0.02 |
| M1BGD4 | Xyloglucan | 0.7 | 0.03 |
| M0ZHI6 | Beta-galactosidase | 0.7 | 0.04 |
| M1CAK9 | Heparanase-2 | 0.6 | 0.00 |
| M1CWU3 | LEXYL2 protein | 0.6 | 0.02 |
| M1D155 | Peroxidase | 0.6 | 0.05 |
| M1CCK1 | Peroxidase | 0.5 | 0.05 |
| M1D1V1 | Hemoglobin | −1.1 | 0.01 |
| M1AY17 | Peroxidase | −1.6 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Wilson, R.; Balotf, S.; Tegg, R.S.; Eyles, A.; Wilson, C.R. Comparative Proteomic Analysis of Potato Roots from Resistant and Susceptible Cultivars to Spongospora subterranea Zoospore Root Attachment In Vitro. Molecules 2022, 27, 6024. https://doi.org/10.3390/molecules27186024
Yu X, Wilson R, Balotf S, Tegg RS, Eyles A, Wilson CR. Comparative Proteomic Analysis of Potato Roots from Resistant and Susceptible Cultivars to Spongospora subterranea Zoospore Root Attachment In Vitro. Molecules. 2022; 27(18):6024. https://doi.org/10.3390/molecules27186024
Chicago/Turabian StyleYu, Xian, Richard Wilson, Sadegh Balotf, Robert S. Tegg, Alieta Eyles, and Calum R. Wilson. 2022. "Comparative Proteomic Analysis of Potato Roots from Resistant and Susceptible Cultivars to Spongospora subterranea Zoospore Root Attachment In Vitro" Molecules 27, no. 18: 6024. https://doi.org/10.3390/molecules27186024
APA StyleYu, X., Wilson, R., Balotf, S., Tegg, R. S., Eyles, A., & Wilson, C. R. (2022). Comparative Proteomic Analysis of Potato Roots from Resistant and Susceptible Cultivars to Spongospora subterranea Zoospore Root Attachment In Vitro. Molecules, 27(18), 6024. https://doi.org/10.3390/molecules27186024