
Mass Spectrometric Identification of Licania rigida Benth Leaf Extracts and Evaluation of Their Therapeutic Effects on Lipopolysaccharide-Induced Inflammatory Response


 , , ,
, , , 

Abstract
:1. Introduction
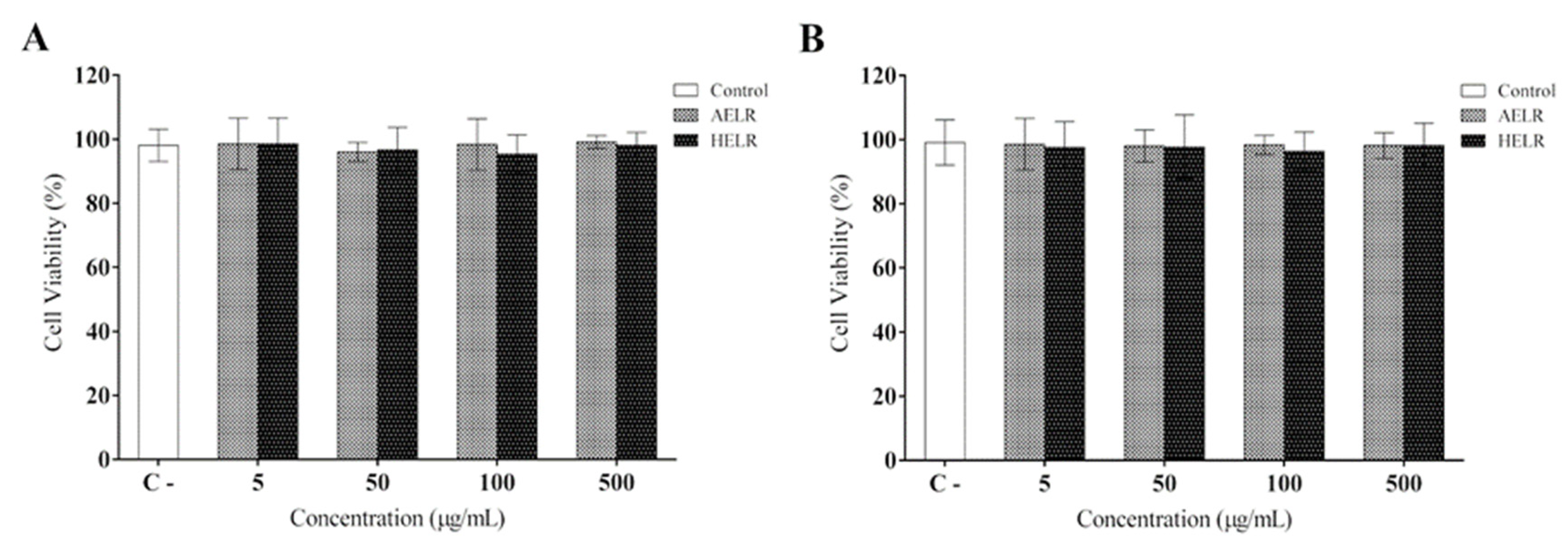
2. Results
3. Discussion
4. Materials and Methods
4.1. Collection of Plant Material and Preparation of Extracts
4.2. Phytochemical Analysis by Ultrafast Liquid Chromatography Coupled with Mass Spectrometry (LC-MS/MS)
4.3. Cell Culture and Animals
4.4. Cell Viability and Cytotoxicity Assays
4.5. Leukocyte Migration into Peritoneal Cavity and Cytokine Dosage
4.6. Cytokine Measurement (TNF-α, IL1-β, and IL-6)
4.7. Measurement of Nitric Oxide (NO) Production
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chan, J.T.H.; Kadri, S.; Köllner, B.; Rebl, A.; Korytář, T. RNA-seq of single fish cells—Seeking out the leukocytes mediating immunity in teleost fishes. Front. Immunol. 2022, 13, 1664–3224. [Google Scholar] [CrossRef] [PubMed]
- Morla, S. Glycosaminoglycans and glycosaminoglycan mimetics in cancer and inflammation. Int. J. Mol. Sci. 2019, 20, 1963. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Pirlamarla, P.; Bond, R.M. FDA labeling of NSAIDs: Review of nonsteroidal anti-inflammatory drugs in cardiovascular disease. Trends Cardiovasc. Med. 2016, 26, 675–680. [Google Scholar] [CrossRef]
- Ross, E.A.; Devitt, A.; Johnson, J.R. Macrophages: The good, the bad, and the gluttony. Front. Immunol. 2021, 12, 708186. [Google Scholar] [CrossRef]
- Fuijiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drugs Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Cavallion, J.M. Cytokines and macrophages. Biomed. Pharmacother. 1994, 10, 445–453. [Google Scholar] [CrossRef]
- Kamimura, D.; Ishihara, K.; Hirano, T. IL-6 signal transduction and its physiological roles: The signal orchestration model. Physiol. Biochem. Pharmacol. 2003, 149, 1–38. [Google Scholar]
- Siregar, A.S.; Werdhani, R.A.; Ascobat, P.; Nafrialdi, N.; Syam, A.F.; Hidayat, R.; Wangge, G. Development of module for the prevention of nonsteroidal anti-inflammatory drugs-associated gastrointestinal adverse reactions in the elderly at a primary health center. Int. J. Risk Saf. Med. 2021, 32, 61–73. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; The International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern approaches in the discovery and development of plant-based natural products and their analogues as potential therapeutic agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, U.P.; Medeiros, P.M.; Almeida, A.L.S.; Monteiro, J.M.; Lins Neto, E.M.F.; Melo, J.G.; Santos, J.P. Medicinal plants of the caatinga (semi-arid) vegetation of NE Brazil: A quantitative approach. J. Ethnopharmacol. 2007, 114, 325–354. [Google Scholar] [CrossRef] [PubMed]
- Feitosa, E.A.; Xavier, H.S.; Randau, K.P. Chrysobalanaceae: Traditional uses, phytochemistry and pharmacology. Rev. Bras. Farmacogn. 2012, 22, 1181–1186. [Google Scholar] [CrossRef]
- Pessoa, I.P.; Neto, J.J.L.; de Almeida, T.S.; Farias, D.F.; Vieira, L.R.; de Medeiros, J.L.; Boligon, A.A.; Peijnenburg, A.; Castelar, I.; Carvalho, A.F.U. Polyphenol composition, antioxidant activity and cytotoxicity of seeds from two underexploited wild Licania species: L. rigida and L. tomentosa. Molecules 2016, 21, 1755. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.M.; Lopes, H.T.O.; Rodrigues, F.A.; Coutinho, H.D.M.; Oliveira, L.C.C. Antioxidant potential of the Caatinga flora. Phytomed. Plus. 2022, 2, 100240. [Google Scholar] [CrossRef]
- Morais, L.V.F.; Luz, J.R.D.; Nascimento, T.E.S.; Azevedo, M.A.S.; Rocha, W.P.S.; Araujo-Silva, G.; Ururahy, M.A.G.; Chaves, G.M.; López, J.A.; Santos, E.C.G.; et al. Phenolic composition, toxicity potential, and antimicrobial activity of Licania rigida Benth (Chrysobalanaceae) leaf extracts. J. Med. Food 2022, 25, 97–109. [Google Scholar] [CrossRef]
- Santos, E.S.; Oliveira-Tintino, C.D.M.; Correia, D.B.; Alencar, C.D.C.; Sousa, M.F.; Lima, C.N.F.; Machado, S.T.S.; Gomes, A.D.S.; Garcia, F.A.O.; Menezes, I.R.A.; et al. Topical anti-inflammatory effect of hydroalcoholic extract of leaves of Licania rigida Benth. in mice. Phytomed. Plus 2021, 1, 100110. [Google Scholar] [CrossRef]
- Linus, L.O.; Wang, S.L.; Shi, N.; Hanson, C.; Lu, Y.T.; Alolga, R.N.; Liu, Q.; Njokuocha, R.C.; Qi, L.W. The new plant Parinari kerstinguii Engl: Toxicity studies and anti-inflammatory properties. J. Ethnopharmacol. 2018, 220, 101016. [Google Scholar] [CrossRef]
- Venancio, V.P.; Cipriano, P.A.; Kim, H.; Antunes, L.M.G.; Talcott, S.T.; Mertens-Talcott, S.U. Cocoplum (Chrysobalanus icaco L.) anthocyanins exert anti-inflammatory activity in human colon cancer and non-malignant colon cells. Food Funct. 2017, 8, 307–331. [Google Scholar] [CrossRef]
- Santos, E.S.; Oliveira, C.D.M.; Menezes, I.R.A.; Nascimento, E.P.; Correia, D.B.; Alencar, C.D.C.; Sousa, M.F.; Lima, C.N.F.; Monteiro, A.B.; Souza, C.P.E.; et al. Anti-inflammatory activity of herb products from Licania rigida Benth. Complement. Ther. Med. 2019, 45, 254–261. [Google Scholar] [CrossRef]
- Carnevale, N.F.; Pilon, A.C.; Silva, B.V. Chrysobalanaceae: Secondary metabolites, ethnopharmacology and pharmacological potential. Phytochem. Rev. 2013, 12, 121–146. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as potential anti-inflammatory molecules: A review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef] [PubMed]
- Da Luz, J.R.D.; do Nascimento, T.E.S.; Araujo-Silva, G.; de Rezende, A.A.; Brandão-Neto, J.; Ururahy, M.A.G.; Luchessi, A.D.; López, J.A.; Rocha, H.A.O.; das Graças Almeida, M. Licania rigida Benth leaf extracts: Assessment of toxicity and potential anticoagulant effect. S. Afr. J. Bot. 2021, 139, 217–225. [Google Scholar] [CrossRef]
- Bernal, A.J.; da Silva, M.M.G.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; De los Reyes, V.; Martín-Quirós, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L. Molnupiravir for oral treatment of COVID-19 in nonhospitalized patients. N. Engl. J. Med. 2022, 386, 509–520. [Google Scholar] [CrossRef]
- Poredos, P.; Poredos, P. Involvement of inflammation in venous thromboembolic disease: An update in the age of COVID-19. Semin. Thromb. Hemost. 2021, 48, 93–99. [Google Scholar] [CrossRef]
- Singh, A.K.; Malviya, R. Coagulation and inflammation in cancer: Limitations and prospects for treatment. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188727. [Google Scholar] [CrossRef]
- Oliveira, M.C.B.; Cruz, C.K.S.; Rocha, G.M.M.; Brito, M.G.A.; Oliveira, G.A.L. Toxicity and antibacterial activity of medicinal plants used in the treatment of respiratory diseases: An integrative review. Res. Soc. Dev. 2020, 9, e244997169. [Google Scholar] [CrossRef]
- van Wyk, A.S.; Prinsloo, G. Health, safety and quality concerns of plant-based traditional medicines and herbal remedies. S. Afr. J. Bot. 2020, 133, 54–62. [Google Scholar] [CrossRef]
- Vieira, E.O.G.; Fernandes, R.M.T. Toxic effects of medicinal plants commercialized in natura in São Luís/MA: A literature review. Res. Soc. Dev. 2021, 10, e55910514821. [Google Scholar] [CrossRef]
- Gaston, T.E.; Mendrick, D.L.; Paine, M.F.; Roe, A.L.; Yeung, C.K. Natural is not synonymous with safe: Toxicity of natural products alone and in combination with pharmaceutical agents. Regul. Toxicol. Pharmacol. 2020, 113, 104642. [Google Scholar] [CrossRef] [PubMed]
- Batista, D.; Luz, J.R.D.; Nascimento, T.E.S.; Senes-Lopes, T.F.; Galdino, O.A.; Silva, S.V.; Ferreira, M.P.; Azevedo, M.A.S.; Brandão-Neto, J.; Araujo-Silva, G.; et al. Licania rigida leaf extract: Protective effect on oxidative stress, associated with cytotoxic, mutagenic and preclinical aspects. J. Toxicol. Environ. Health Part A 2021, 20, 276–290. [Google Scholar] [CrossRef]
- Kameritsch, P.; Rrnkawitz, J. Principles of leukocyte migration strategies. Trends Cell Biol. 2020, 30, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Hong, J.; Mi, J.; Sun, B.; Zhang, J.; Li, C.; Yang, W. Polyphenols extracted from Enteromorpha clathrata alleviates inflammation in lipopolysaccharide-induced RAW 264.7 cells by inhibiting the MAPKs/NF-κB signaling pathways. J. Ethnopharmacol. 2022, 286, 114897. [Google Scholar] [CrossRef]
- Moninuola, O.O.; Milligan, W.; Lochhead, P.; Khalili, H. Systematic review with meta-analysis: Association between acetaminophen and nonsteroidal anti-inflammatory drugs (NSAIDs) and risk of Crohn’s disease and ulcerative colitis exacerbation. Aliment. Pharmacol. Ther. 2018, 47, 1428–1439. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.J.; Bian, Z.X.; Qiu, H.C.; Wang, Y.T.; Wang, Y. Biological and clinical implications of herbal medicine and natural products for the treatment of inflammatory bowel disease. Ann. N. Y. Acad. Sci. 2017, 1401, 37–38. [Google Scholar] [CrossRef]
- Trinh, H.K.T.; Pham, L.D.; Le, K.M.; Park, H.S. Pharmacogenomics of hypersensitivity to non-steroidal anti-inflammatory drugs. Front. Genet. 2021, 12, 647257. [Google Scholar] [CrossRef]
- Nunes, C.R.; Arantes, M.B.; de Faria Pereira, S.M.; da Cruz, L.L.; de Souza Passos, M.; de Moraes, L.P.; Vieira, I.J.C.; de Oliveira, D.B. Plants as sources of anti-inflammatory agents. Molecules 2020, 25, 3726. [Google Scholar] [CrossRef]
- Shazhni, J.R.A.; Renu, A.; Vijayaraghavan, P. Insights of antidiabetic, anti-inflammatory and hepatoprotective properties of antimicrobial secondary metabolites of corm extract from Caladium x hortulanum. Saudi J. Biol. Sci. 2018, 25, 1755–1761. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Valli, V.; Tomás-Cobos, L.; Tomás-Chisbert, T.; Murgui-Bosch, L.; Danesi, F.; Bordoni, A. Is cytotoxicity a determinant of the different in vitro and in vivo effects of bioactives? BMC Complement. Altern. Med. 2017, 17, 453. [Google Scholar] [CrossRef]
- Beigh, S.; Rehman, M.U.; Khan, A.; Patil, B.R.; Makeen, H.A.; Rasool, S.; Rashid, S.; Arafah, A.; Kamal, M.A. Therapeutic role of flavonoids in lung inflammatory disorders. Phytomed. Plus 2022, 2, 100221. [Google Scholar] [CrossRef]
- Alzaabi, M.M.; Hamdy, R.; Ashmawy, N.S.; Hamoda, A.M.; Alkhayat, F.; Khademi, N.N.; Al Joud, S.M.A.; El-Keblawy, A.A.; Soliman, S.M.M. Flavonoids are promising safe therapy against COVID-19. Phytochem. Rev. 2022, 21, 291–312. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Virgous, C.; Si, H. Synergistic anti-inflammatory effects and mechanisms of combined phytochemicals. J. Nutr. Biochem. 2019, 69, 19–30. [Google Scholar] [CrossRef]
- He, M.; Min, J.W.; Kong, W.L.; He, X.H.; Li, J.X.; Peng, B.W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, X.; Wang, J. Isovitexin protects against cisplatin-induced kidney injury in mice through inhibiting inflammatory and oxidative responses. Int. Immunopharmacol. 2020, 83, 106437. [Google Scholar] [CrossRef]
- Xi, S.; Shao, Z.; Li, L.; Gui, Z.; Liu, P.; Jiang, Q.; Yu, Y.; Zhou, W.; Zhou, Z.; Zhang, S.; et al. Tongbixiao pills improve gout by reducing uric acid levels and inhibiting inflammation. Dose-Response 2022, 20, 1–11. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharmacother. 2021, 133, 110985. [Google Scholar] [CrossRef]
- Toyama, M.H.; Rogero, A.; de Moraes, L.L.F.; Fernandes, G.A.; da Cruz Costa, C.R.; Belchor, M.N.; De Carli, A.M.; de Oliveira, M.A. Gallic acid as a non-selective inhibitor of α/β-hydrolase fold enzymes involved in the inflammatory process: The two sides of the same coin. Pharmaceutics 2022, 14, 368. [Google Scholar] [CrossRef]
- Rios, L.L.; Giner, R.M.; Marín, M.; Recio, M.C. A pharmacological update of ellagic acid. Planta Med. 2018, 84, 1068–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allam, G.; Mahdi, E.A.; Alzahrani, A.M.; Abuelsaad, A.S. Ellagic acid alleviates adjuvant induced arthritis by modulation of pro- and anti-inflammatory cytokines. Cent. Eur. J. Immunol. 2016, 41, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Basak, P.; Dutta, S.; Chowdhury, S.; Sil, P.C. New insights into the ameliorative effects of ferulic acid in pathophysiological conditions. Food Chem. Toxicol. 2017, 103, 41–55. [Google Scholar] [CrossRef]
- Pasquini, S.; Contri, C.; Borea, P.A.; Vincenzi, F.; Varani, K. Adenosine and inflammation: Here, there and everywhere. Int. J. Mol. Sci. 2021, 22, 7685. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Guo, A.; Li, F.; Zhang, F.; Bi, M.; Zhang, Y.; Zhu, W. Liquiritigenin enhances cyclic adenosine monophosphate production to mitigate inflammation in dendritic cells. Int. J. Immunopathol. Pharmacol. 2021, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bergantin, L.B. Diabetes and inflammatory diseases: An overview from the perspective of Ca2+/3′-5′-cyclic adenosine monophosphate signaling. World J. Diabetes 2021, 12, 767–779. [Google Scholar] [CrossRef]
- Pemovska, T.; Bigenzahn, J.; Superti-Furga, G. Recent advances in combinatorial drug screening and synergy scoring. Curr. Opin. Pharmacol. 2018, 42, 102–110. [Google Scholar] [CrossRef]
- Ma, J.; Motsinger-Reif, A. Current methods for quantifying drug synergism. Proteom. Bioinform. 2019, 1, 43–48. [Google Scholar]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-inflammatory activity of natural products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Cui, H.; Liu, G.; Zhao, X.; Li, W.; Piao, G. How can synergism of traditional medicines benefit from network pharmacology? Molecules 2017, 22, 1135. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Alhadrami, H.A.; Amin, E.; Aly, H.F.; Othman, A.M.; Rateb, M.E.; Hetta, M.H.; Abdelmohsen, U.R.; Hassan, H.M. Anti-inflammatory and antioxidant activities of terpene- and polyphenol-rich Premna odorata leaves on alcohol-inflamed female Wistar albino rat liver. Molecules 2020, 25, 3116. [Google Scholar] [CrossRef] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxid Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Aware, C.B.; Patil, D.N.; Suryawanshi, S.S.; Mali, P.R.; Rane, M.R.; Gurav, R.G.; Jadhav, J.P. Natural bioactive products as promising therapeutics: A review of natural product-based drug development. S. Afr. J. Bot. 2022, in press. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Y.; Zhou, Y.; Lian, X.; Yan, L.; Pan, T.; Jin, T.; Xie, H.; Liang, Z.; Qiu, W.; et al. NPCDR: Natural product-based drug combination and its disease-specific molecular regulation. Nucleic Acids Res. 2022, 50, D1324–D1333. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compound | Cosine | MassDiff | Mass | Molecular Formula | Ion Fragments | Adduct | Extract |
|---|---|---|---|---|---|---|---|---|
| 1 | Adenosine monophosphate | 0.89 | 0.001 | 348.071 | C10H14N5O7P | 250.09, 178.07, 136.06 | [M + H]+ | AELR |
| 2 | Gallic acid | 0.97 | 0 | 171.029 | C7H6O5 | 153.02, 135.01, 127.04, 125.02, 109.03, 107.01, 81.04 | [M + H]+ | AELR |
| 3 | 5′-Deoxy-5′-(methylsulfinyl) adenosine | 0.91 | 0.003 | 314.093 | C11H15N5O4S | 296.08, 164.06, 136.06, 97.03 | [M + H]+ | AELR |
| 4 | Ellagic acid | 0.86 | 0.038 | 303.048 | C14H6O8 | 285.00, 275.02, 257.01, 201.02 | [M + H]+ | AELR |
| 1 | Gallic acid | 0.94 | 0 | 171.029 | C7H6O5 | 153.02, 127.04, 125.02, 109.03, 107.01, 81.04 | [M + H]+ | HELR |
| 2 | Phenylalanine | 0.93 | 0 | 166.086 | C9H11NO2 | 149.06, 120.08, 103.05 | [M + H]+ | HELR |
| 3 | 2,6-Xylidine | 0.96 | 0 | 122.096 | C8H11N | 105.07, 79.06, 77.04, 51.04 | [M + H]+ | HELR |
| 4 | Pyridoxine | 0.87 | 0 | 170.081 | C8H11NO3 | 152.07, 142.05, 134.06, 124.08, 96.05 | [M + H]+ | HELR |
| 5 | DL-Indole-3-lactic acid | 0.98 | 0.001 | 188.071 | C11H11NO3 | 146.06, 118.07, 65.04 | [M + H-H2O]+ | HELR |
| 6 | 4-Aminobutanoate | 0.87 | 0 | 104.071 | C4H9NO2 | 87.05, 86.06, 69.04 | [M + H]+ | HELR |
| 7 | Isovitexin | 0.97 | 0.003 | 433.113 | C21H20O10 | 415.10, 313.07, 283.06, 217.05 | [M + H]+ | HELR |
| 8 | Ferulate | 0.93 | 0 | 177.054 | C10H10O4 | 149.06, 145.03, 117.07, 89.04, 63.03 | [M + H-H2O]+ | HELR |
| 9 | Pheophorbide A | 0.89 | 0.032 | 593.237 | C35H36N4O5 | 533.25, 461.23, 460.22, 433.24 | [M + H]+ | HELR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, T.E.S.d.; López, J.A.; Barbosa, E.A.; Ururahy, M.A.G.; Brito, A.d.S.; Araujo-Silva, G.; Luz, J.R.D.d.; Almeida, M.d.G. Mass Spectrometric Identification of Licania rigida Benth Leaf Extracts and Evaluation of Their Therapeutic Effects on Lipopolysaccharide-Induced Inflammatory Response. Molecules 2022, 27, 6291. https://doi.org/10.3390/molecules27196291
Nascimento TESd, López JA, Barbosa EA, Ururahy MAG, Brito AdS, Araujo-Silva G, Luz JRDd, Almeida MdG. Mass Spectrometric Identification of Licania rigida Benth Leaf Extracts and Evaluation of Their Therapeutic Effects on Lipopolysaccharide-Induced Inflammatory Response. Molecules. 2022; 27(19):6291. https://doi.org/10.3390/molecules27196291
Chicago/Turabian StyleNascimento, Thayse Evellyn Silva do, Jorge A. López, Eder Alves Barbosa, Marcela Abbott Galvão Ururahy, Adriana da Silva Brito, Gabriel Araujo-Silva, Jefferson Romáryo Duarte da Luz, and Maria das Graças Almeida. 2022. "Mass Spectrometric Identification of Licania rigida Benth Leaf Extracts and Evaluation of Their Therapeutic Effects on Lipopolysaccharide-Induced Inflammatory Response" Molecules 27, no. 19: 6291. https://doi.org/10.3390/molecules27196291
APA StyleNascimento, T. E. S. d., López, J. A., Barbosa, E. A., Ururahy, M. A. G., Brito, A. d. S., Araujo-Silva, G., Luz, J. R. D. d., & Almeida, M. d. G. (2022). Mass Spectrometric Identification of Licania rigida Benth Leaf Extracts and Evaluation of Their Therapeutic Effects on Lipopolysaccharide-Induced Inflammatory Response. Molecules, 27(19), 6291. https://doi.org/10.3390/molecules27196291