


Quercetin and Resveratrol Differentially Decrease Expression of the High-Affinity IgE Receptor (FcεRI) by Human and Mouse Mast Cells


Abstract
:1. Introduction
2. Results
2.1. Effects of Quercetin and Resveratrol on FcεRI Expression and TNF Release by MC/9
2.1.1. Quercetin Inhibits the Expression of FcεRI by MC/9
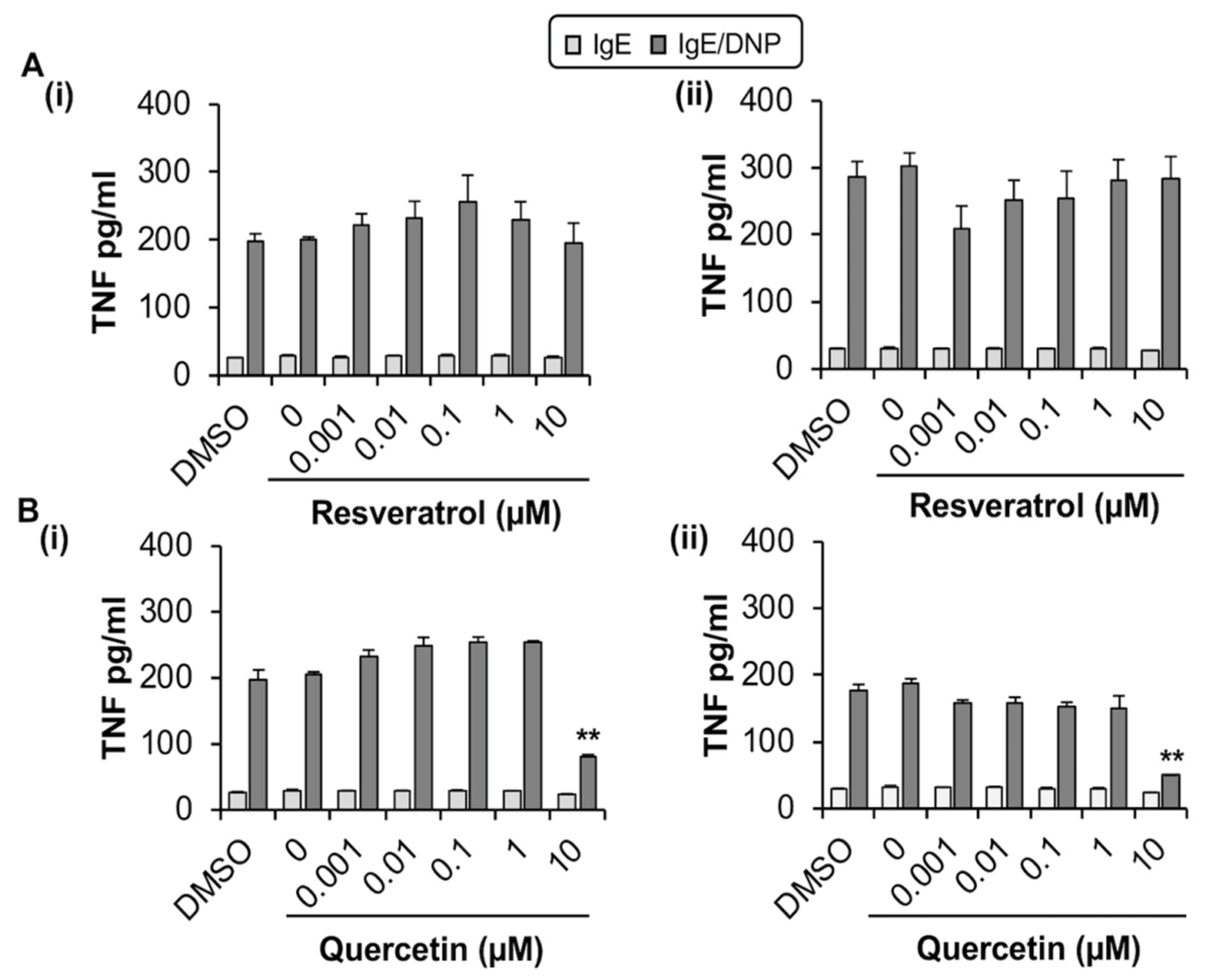
2.1.2. Quercetin Inhibits TNF Release from MC/9
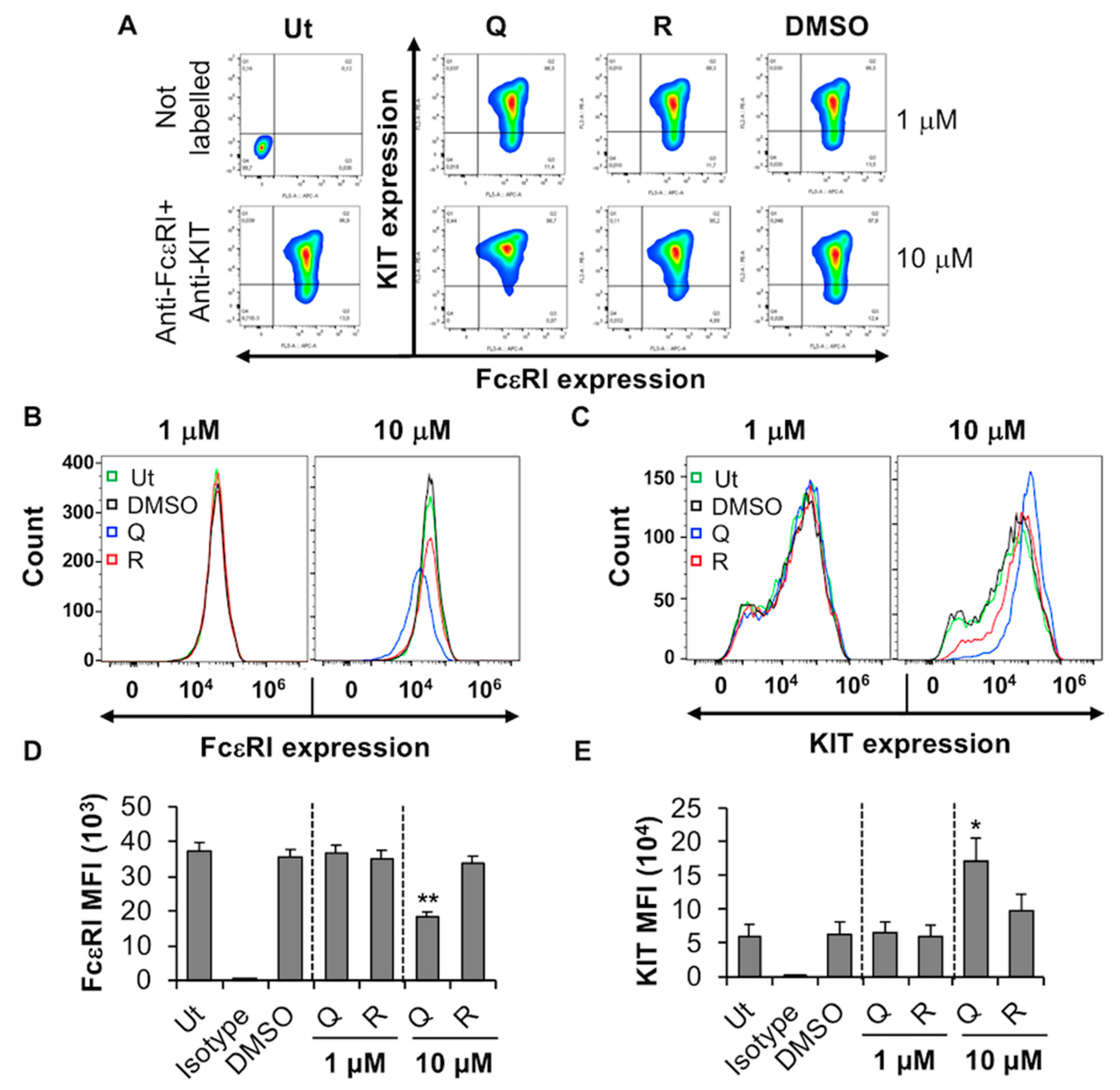
2.2. Effect of Quercetin and Resveratrol on FcεRI Expression, FcεRI-Mediated Signaling, Degranulation and TNF Release by BMMC
2.2.1. Quercetin and Resveratrol Inhibit the Expression of FcεRI by BMMC
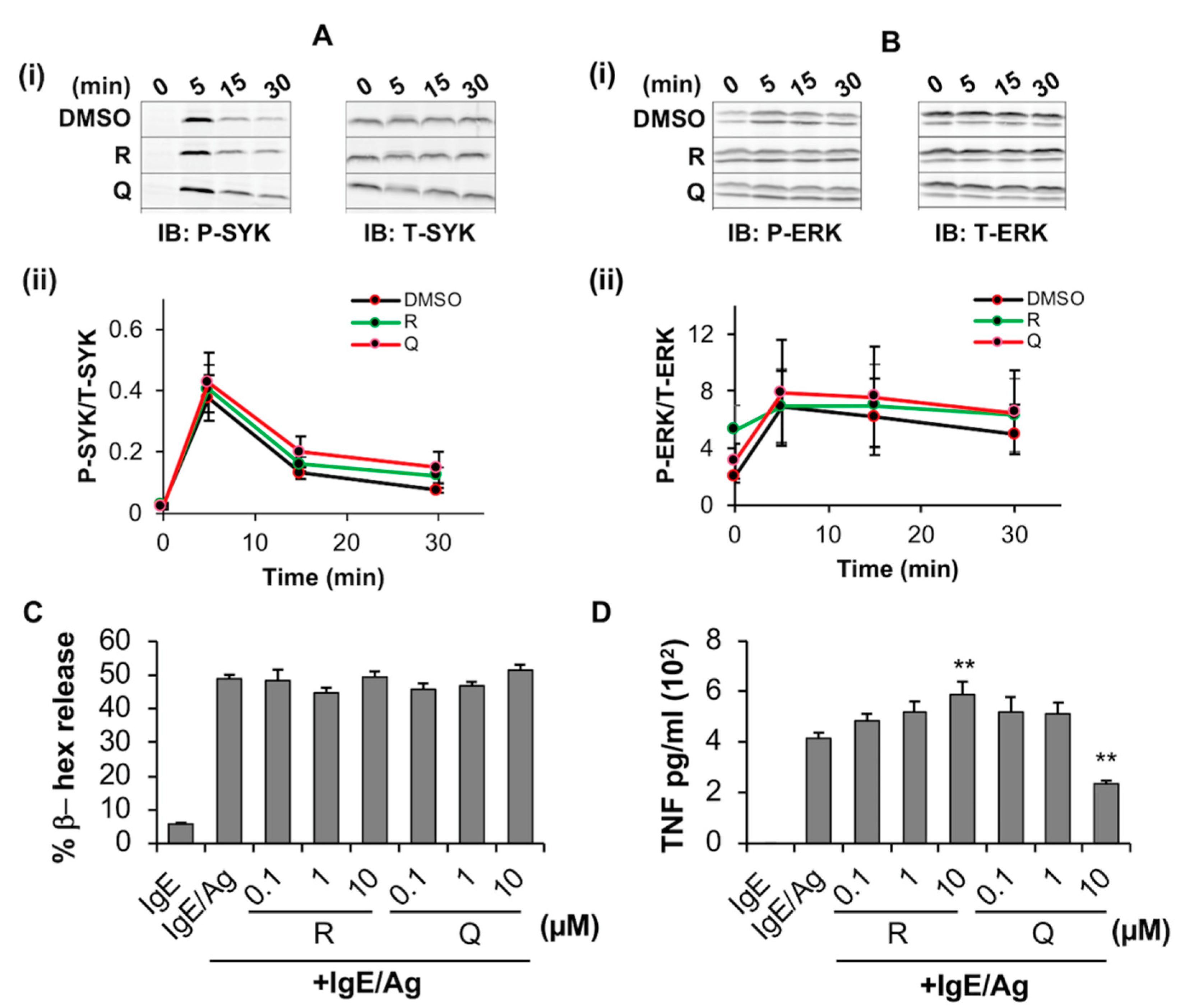
2.2.2. Quercetin Inhibits the Release of TNF from BMMC but Has No Measurable Effect on Phosphorylation of SYK and ERK or Degranulation
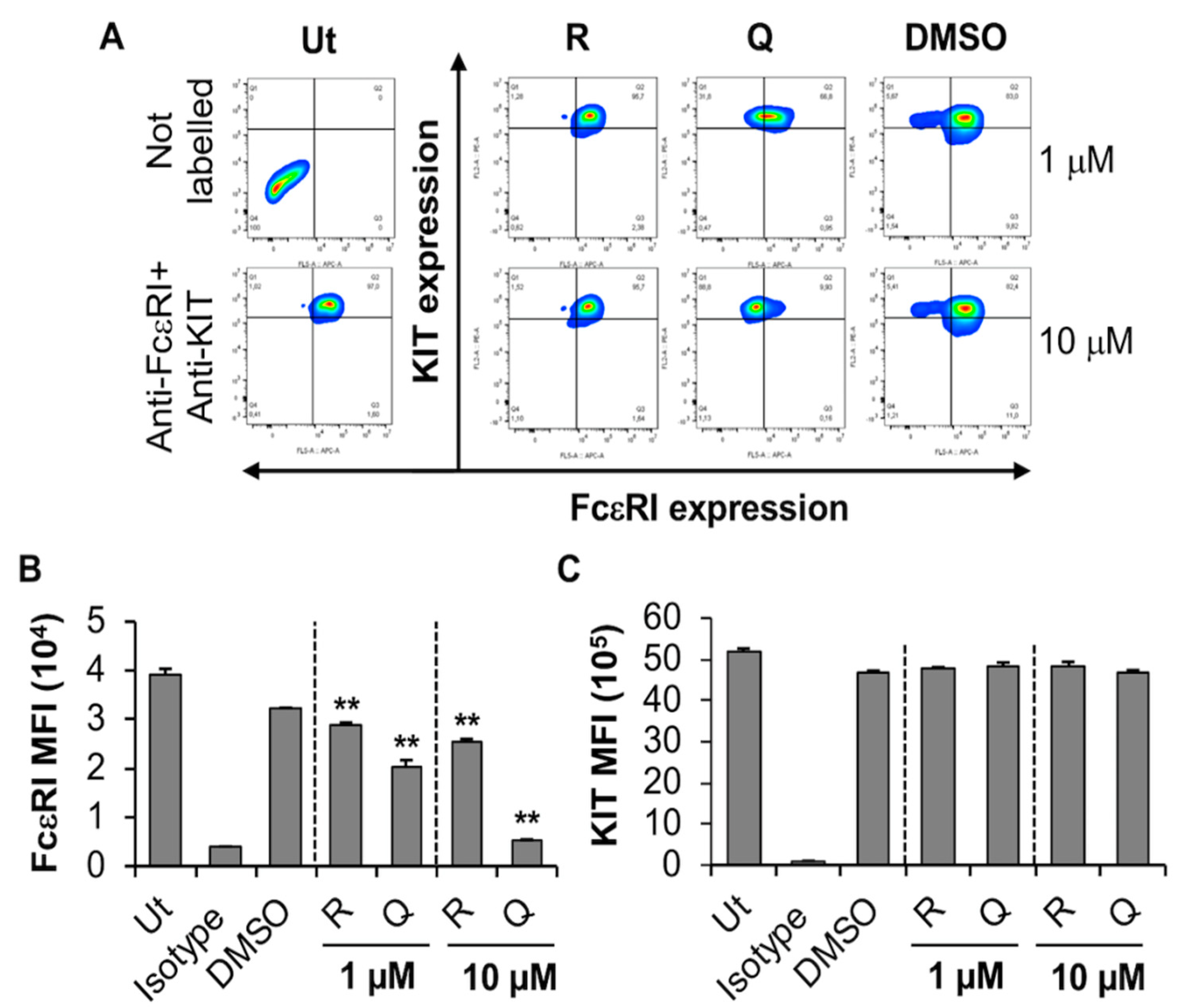
2.3. Effect of Quercetin and Resveratrol on FcεRI Expression, FcεRI-Mediated Signaling and Degranulation by LAD2
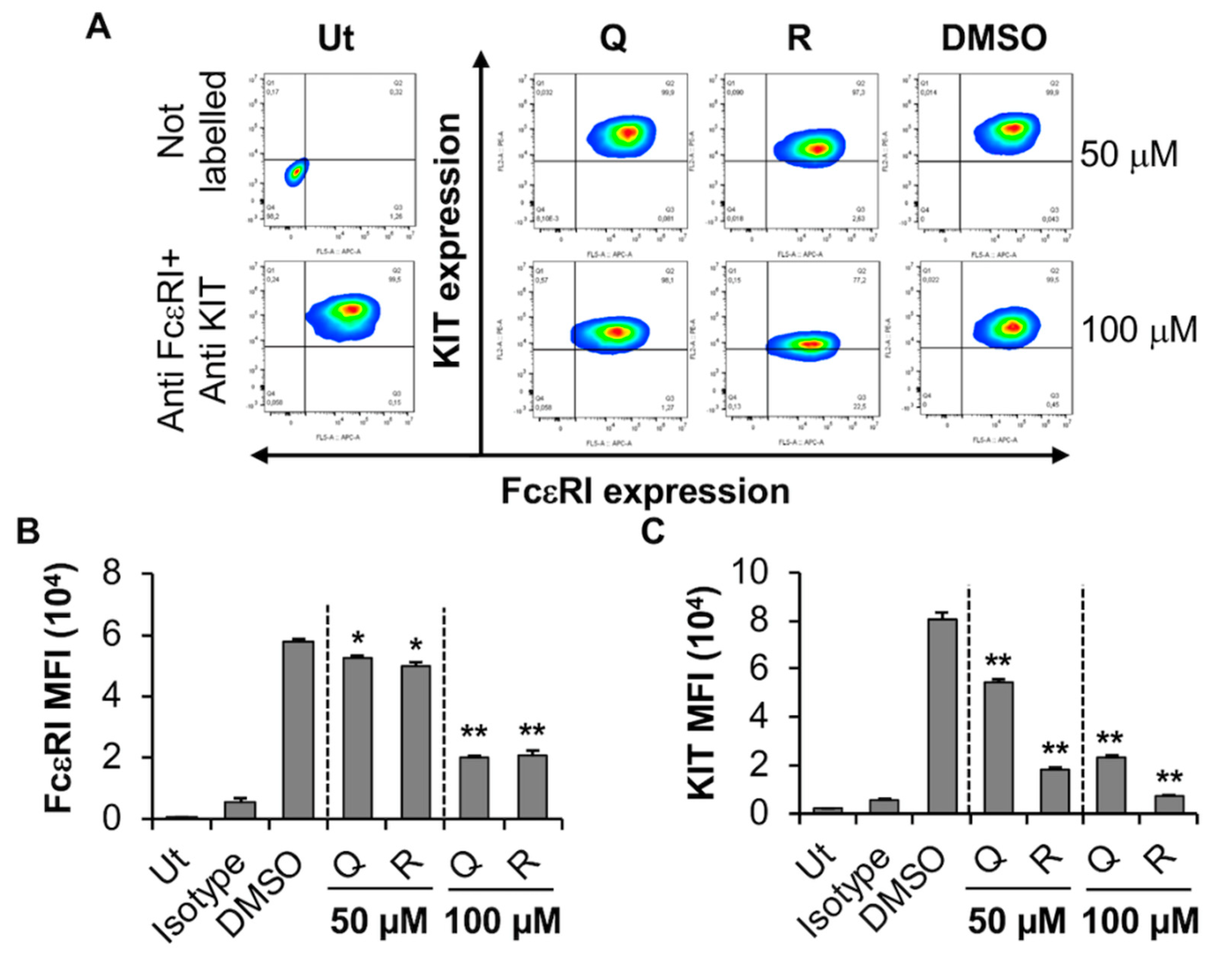
2.3.1. Quercetin and Resveratrol Inhibit the Expression of KIT While Moderately Affecting FcεRI Expression by the Human MC Line, LAD2
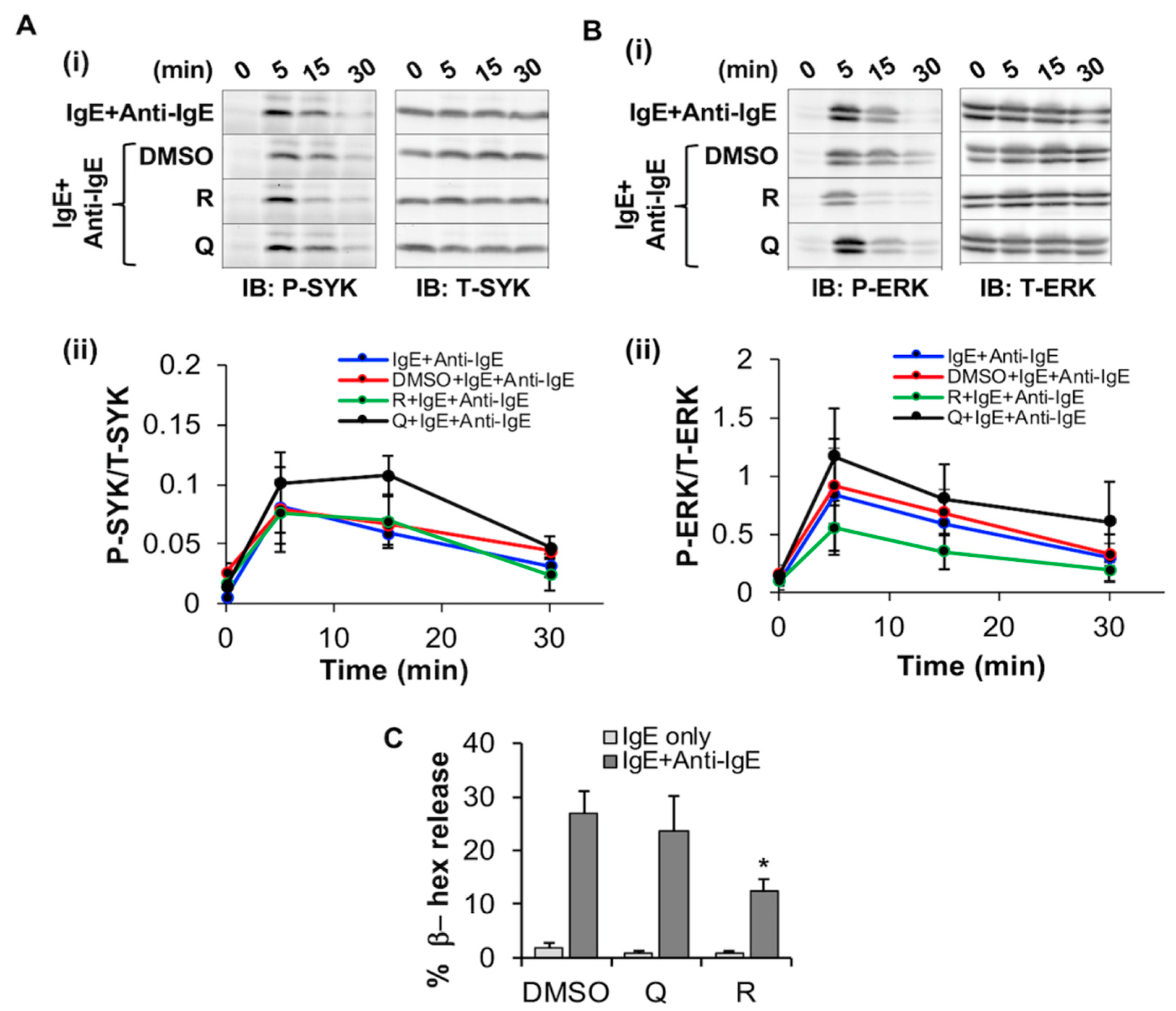
2.3.2. Resveratrol Inhibits FcεRI-Dependent Signaling and Degranulation in LAD2
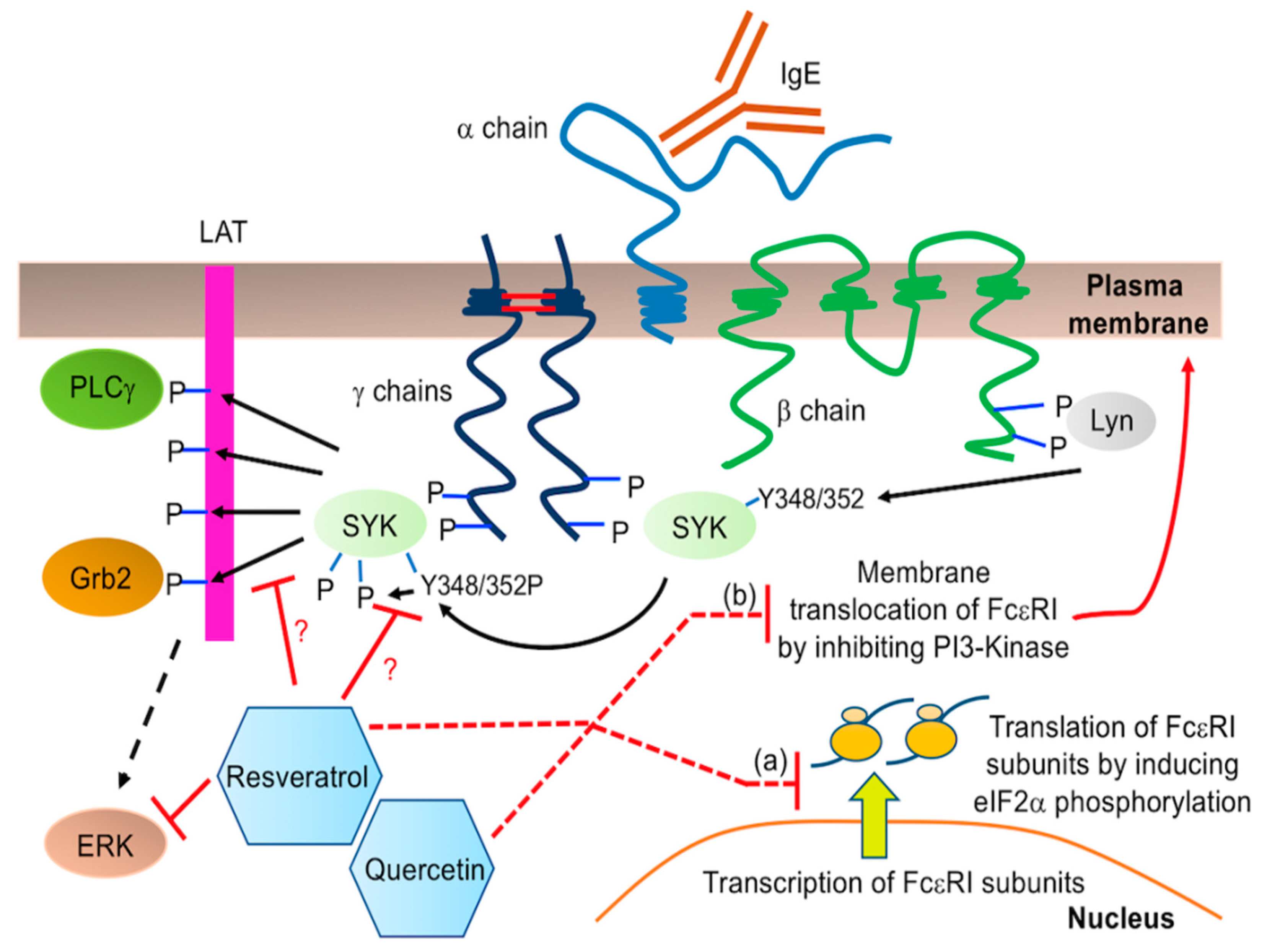
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Trypan Blue Exclusion Assay
4.3. Degranulation Assay
4.4. ELISA
4.5. Flow Cytometric Analysis
4.6. Western Blot
4.7. RNA Extraction and cDNA Synthesis
4.8. qPCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duthie, S.J.; Dobson, V.L. Dietary flavonoids protect human colonocyte DNA from oxidative attack in vitro. Eur. J. Nutr. 1999, 38, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.A.; Takaishi, Y.; Shirotori, M.; Kawaguchi, Y.; Tsuchiya, K.; Shibata, H.; Higuti, T.; Tadokoro, T.; Takeuchi, M. Antibacterial and antioxidant activities of quercetin oxidation products from yellow onion (Allium cepa) skin. J. Agric. Food Chem. 2006, 54, 3551–3557. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Teoh, B.T.; Sam, S.S.; Wong, P.F.; Mustafa, M.R.; Abubakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 2011, 8, 560. [Google Scholar] [CrossRef] [Green Version]
- Boots, A.W.; Wilms, L.C.; Swennen, E.L.R.; Kleinjans, J.C.S.; Bast, A.; Haenen, G.R.M.M. In vitro and ex vivo anti-inflammatory activity of quercetin in healthy volunteers. Nutrition 2008, 24, 703–710. [Google Scholar] [CrossRef]
- Alam, S.B.; Rochon, D. Cucumber Necrosis Virus Recruits Cellular Heat Shock Protein 70 Homologs at Several Stages of Infection. J. Virol. 2015, 90, 3302–3317. [Google Scholar] [CrossRef] [Green Version]
- Agullo, G.; Gamet-Payrastre, L.; Manenti, S.; Viala, C.; Rémésy, C.; Chap, H.; Payrastre, B. Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: A comparison with tyrosine kinase and protein kinase C inhibition. Biochem. Pharmacol. 1997, 53, 1649–1657. [Google Scholar] [CrossRef]
- Gábor, M. Anti-inflammatory and anti-allergic properties of flavonoids. Prog. Clin. Biol. Res. 1986, 213, 471–480. [Google Scholar]
- Boesch-Saadatmandi, C.; Wagner, A.E.; Wolffram, S.; Rimbach, G. Effect of quercetin on inflammatory gene expression in mice liver in vivo—Role of redox factor 1, miRNA-122 and miRNA-125b. Pharmacol. Res. 2012, 65, 523–530. [Google Scholar] [CrossRef]
- Weng, Z.; Zhang, B.; Asadi, S.; Sismanopoulos, N.; Butcher, A.; Fu, X.; Katsarou-Katsari, A.; Antoniou, C.; Theoharides, T.C. Quercetin is more effective than cromolyn in blocking human mast cell cytokine release and inhibits contact dermatitis and photosensitivity in humans. PLoS ONE 2012, 7, e33805. [Google Scholar] [CrossRef] [PubMed]
- Jafarinia, M.; Sadat Hosseini, M.; Kasiri, N.; Fazel, N.; Fathi, F.; Ganjalikhani Hakemi, M.; Eskandari, N. Quercetin with the potential effect on allergic diseases. Allergy Asthma Clin. Immunol. 2020, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Shishehbor, F.; Behroo, L.; Ghafouriyan Broujerdnia, M.; Namjoyan, F.; Latifi, S.M. Quercetin effectively quells peanut-induced anaphylactic reactions in the peanut sensitized rats. Iran. J. Allergy Asthma Immunol. 2010, 9, 27–34. [Google Scholar] [PubMed]
- Moon, H.; Choi, H.H.; Lee, J.Y.; Moon, H.J.; Sim, S.S.; Kim, C.J. Quercetin inhalation inhibits the asthmatic responses by exposure to aerosolized-ovalbumin in conscious guinea-pigs. Arch. Pharm. Res. 2008, 31, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.; Ha, J.H.; Lee, M.G.; Kim, J.E.; et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef]
- Foreman, J.C. Mast cells and the actions of flavonoids. J. Allergy Clin. Immunol. 1984, 73, 769–774. [Google Scholar] [CrossRef]
- Pearce, F.L.; Befus, A.D.; Bienenstock, J. Mucosal mast cells. III. Effect of quercetin and other flavonoids on antigen-induced histamine secretion from rat intestinal mast cells. J. Allergy Clin. Immunol. 1984, 73, 819–823. [Google Scholar] [CrossRef]
- Shichijo, M.; Yamamoto, N.; Tsujishita, H.; Kimata, M.; Nagai, H.; Kokubo, T. Inhibition of syk activity and degranulation of human mast cells by flavonoids. Biol. Pharm. Bull. 2003, 26, 1685–1690. [Google Scholar] [CrossRef] [Green Version]
- Theoharides, T.C.; Alysandratos, K.D.; Angelidou, A.; Delivanis, D.A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast cells and inflammation. Biochim. Biophys. Acta 2012, 1822, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [Green Version]
- Theoharides, T.C.; Kalogeromitros, D. The critical role of mast cells in allergy and inflammation. Ann. N. Y. Acad. Sci. 2006, 1088, 78–99. [Google Scholar] [CrossRef] [PubMed]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Kraft, S.; Kinet, J.P. New developments in FcepsilonRI regulation, function and inhibition. Nat. Rev. Immunol. 2007, 7, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.M.; Abraham, S.N. New roles for mast cells in modulating allergic reactions and immunity against pathogens. Curr. Opin. Immunol. 2009, 21, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Rudich, N.; Ravid, K.; Sagi-Eisenberg, R. Mast Cell Adenosine Receptors Function: A Focus on the A3 Adenosine Receptor and Inflammation. Front. Immunol. 2012, 3, 134. [Google Scholar] [CrossRef] [Green Version]
- Arizmendi, N.; Kulka, M. Adenosine activates Gα(s) proteins and inhibits C3a-induced activation of human mast cells. Biochem. Pharmacol. 2018, 156, 157–167. [Google Scholar] [CrossRef]
- Hosokawa, J.; Suzuki, K.; Nakagomi, D.; Tamachi, T.; Takatori, H.; Suto, A.; Nakajima, H. Role of calcium ionophore A23187-induced activation of IkappaB kinase 2 in mast cells. Int. Arch. Allergy Immunol. 2013, 161 (Suppl. S2), 37–43. [Google Scholar] [CrossRef]
- Beaven, M.A.; Ozawa, K. Role of calcium, protein kinase C and MAP kinase in the activation of mast cells. Allergol. Int. 1996, 45, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, H.; Inaba, M.; Fujikura, T.; Kunitomo, M. Histochemical and functional characteristics of metachromatic cells in the nasal epithelium in allergic rhinitis: Studies of nasal scrapings and their dispersed cells. J. Allergy Clin. Immunol. 1995, 96, 528–536. [Google Scholar] [CrossRef]
- Haggag, E.G.; Abou-Moustafa, M.A.; Boucher, W.; Theoharides, T.C. The effect of a herbal water-extract on histamine release from mast cells and on allergic asthma. J. Herb. Pharmacother. 2003, 3, 41–54. [Google Scholar] [CrossRef]
- Catalli, A.; MacDonald, C.; Pundir, P.; Kulka, M. Inhibitory effects of resveratrol on human mast cell degranulation, cytokine, chemokine and leukotriene release. Open J. Immunol. 2012, 2, 8. [Google Scholar] [CrossRef]
- Bilotta, S.; Arbogast, J.; Schart, N.; Frei, M.; Lorentz, A. Resveratrol Treatment Prevents Increase of Mast Cells in Both Murine OVA Enteritis and IL-10(−/−) Colitis. Int. J. Mol. Sci. 2022, 23, 1213. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Bae, J.Y.; Park, S.H.; Kim, Y.H.; Park, J.H.; Kang, Y.H. Resveratrol inhibits IgE-mediated basophilic mast cell degranulation and passive cutaneous anaphylaxis in mice. J. Nutr. 2013, 143, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilotta, S.; Paruchuru, L.B.; Feilhauer, K.; Köninger, J.; Lorentz, A. Resveratrol Is a Natural Inhibitor of Human Intestinal Mast Cell Activation and Phosphorylation of Mitochondrial ERK1/2 and STAT3. Int. J. Mol. Sci. 2021, 22, 7640. [Google Scholar] [CrossRef] [PubMed]
- Civelek, M.; Bilotta, S.; Lorentz, A. Resveratrol Attenuates Mast Cell Mediated Allergic Reactions: Potential for Use as a Nutraceutical in Allergic Diseases? Mol. Nutr. Food Res. 2022, 66, e2200170. [Google Scholar] [CrossRef] [PubMed]
- Shaik, Y.; Caraffa, A.; Ronconi, G.; Lessiani, G.; Conti, P. Impact of polyphenols on mast cells with special emphasis on the effect of quercetin and luteolin. Cent.-Eur. J. Immunol. 2018, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Fürst, R.; Zündorf, I. Plant-derived anti-inflammatory compounds: Hopes and disappointments regarding the translation of preclinical knowledge into clinical progress. Mediat. Inflamm. 2014, 2014, 146832. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T.; Gander, I.; Wirthmueller, U.; Ravetch, J.V. The beta subunit of the Fc epsilon RI is associated with the Fc gamma RIII on mast cells. J. Exp. Med. 1992, 175, 447–451. [Google Scholar] [CrossRef]
- Turner, H.; Kinet, J.-P. Signalling through the high-affinity IgE receptor FcεRI. Nature 1999, 402, 24–30. [Google Scholar] [CrossRef]
- Kambayashi, T.; Koretzky, G.A. Proximal signaling events in Fc epsilon RI-mediated mast cell activation. J. Allergy Clin. Immunol. 2007, 119, 544–552; quiz 553–544. [Google Scholar] [CrossRef]
- Kulka, M.; Befus, A.D. The dynamic and complex role of mast cells in allergic disease. Arch. Immunol. Ther. Exp. 2003, 51, 111–120. [Google Scholar]
- Wang, X.; Ma, D.W.L.; Kang, J.X.; Kulka, M. n-3 Polyunsaturated fatty acids inhibit Fc ε receptor I-mediated mast cell activation. J. Nutr. Biochem. 2015, 26, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Krajewski, D.; Polukort, S.H.; Gelzinis, J.; Rovatti, J.; Kaczenski, E.; Galinski, C.; Pantos, M.; Shah, N.N.; Schneider, S.S.; Kennedy, D.R.; et al. Protein Disulfide Isomerases Regulate IgE-Mediated Mast Cell Responses and Their Inhibition Confers Protective Effects During Food Allergy. Front. Immunol. 2020, 11, 606837. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef]
- Chachay, V.S.; Kirkpatrick, C.M.; Hickman, I.J.; Ferguson, M.; Prins, J.B.; Martin, J.H. Resveratrol--pills to replace a healthy diet? Br. J. Clin. Pharmacol. 2011, 72, 27–38. [Google Scholar] [CrossRef]
- Nam, N.H. Naturally occurring NF-kappaB inhibitors. Mini Rev. Med. Chem. 2006, 6, 945–951. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, C.M.; Jung, I.D.; Lee, J.S.; Jeong, Y.I.; Chang, J.H.; Chun, S.H.; Kim, M.J.; Choi, I.W.; Ahn, S.C.; et al. Quercetin regulates Th1/Th2 balance in a murine model of asthma. Int. Immunopharmacol. 2009, 9, 261–267. [Google Scholar] [CrossRef]
- Tanaka, Y.; Furuta, A.; Asano, K.; Kobayashi, H. Modulation of Th1/Th2 Cytokine Balance by Quercetin In Vitro. Medicines 2020, 7, 46. [Google Scholar] [CrossRef]
- Nakajima, S.; Ishimaru, K.; Kobayashi, A.; Yu, G.; Nakamura, Y.; Oh-Oka, K.; Suzuki-Inoue, K.; Kono, K.; Nakao, A. Resveratrol inhibits IL-33-mediated mast cell activation by targeting the MK2/3-PI3K/Akt axis. Sci. Rep. 2019, 9, 18423. [Google Scholar] [CrossRef] [Green Version]
- Porebski, G.; Kwiecien, K.; Pawica, M.; Kwitniewski, M. Mas-Related G Protein-Coupled Receptor-X2 (MRGPRX2) in Drug Hypersensitivity Reactions. Front. Immunol. 2018, 9, 3027. [Google Scholar] [CrossRef] [Green Version]
- Castellani, M.L.; Kempuraj, D.; Frydas, S.; Theoharides, T.C.; Simeonidou, I.; Conti, P.; Vecchiet, J. Inhibitory Effect of Quercetin on Tryptase and MCP-1 Chemokine Release, and Histidine Decarboxylase mRNA Transcription by Human Mast Cell-1 Cell Line. Neuroimmunomodulation 2006, 13, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Kandere-Grzybowska, K.; Kempuraj, D.; Cao, J.; Cetrulo, C.L.; Theoharides, T.C. Regulation of IL-1-induced selective IL-6 release from human mast cells and inhibition by quercetin. Br. J. Pharmacol. 2006, 148, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Kyaw, M.; Yoshizumi, M.; Tsuchiya, K.; Izawa, Y.; Kanematsu, Y.; Tamaki, T. Atheroprotective effects of antioxidants through inhibition of mitogen-activated protein kinases. Acta Pharmacol. Sin. 2004, 25, 977–985. [Google Scholar] [PubMed]
- Kim, M.; Lim, S.J.; Kang, S.W.; Um, B.H.; Nho, C.W. Aceriphyllum rossii extract and its active compounds, quercetin and kaempferol inhibit IgE-mediated mast cell activation and passive cutaneous anaphylaxis. J. Agric. Food Chem. 2014, 62, 3750–3758. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Yoou, M.S.; Han, N.R.; Hwang, S.Y.; Yoon, K.W.; Kim, H.M. The new therapeutic herbal drug HM0601 and its bioactive compound rutin exert potent antiproliferative activities in mast cells. Fundam. Clin. Pharmacol. 2018, 32, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Nanua, S.; Zick, S.M.; Andrade, J.E.; Sajjan, U.S.; Burgess, J.R.; Lukacs, N.W.; Hershenson, M.B. Quercetin blocks airway epithelial cell chemokine expression. Am. J. Respir. Cell Mol. Biol. 2006, 35, 602–610. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Faris, A.N.; Comstock, A.T.; Wang, Q.; Nanua, S.; Hershenson, M.B.; Sajjan, U.S. Quercetin inhibits rhinovirus replication in vitro and in vivo. Antivir. Res. 2012, 94, 258–271. [Google Scholar] [CrossRef]
- Russo, M.; Milito, A.; Spagnuolo, C.; Carbone, V.; Rosén, A.; Minasi, P.; Lauria, F.; Russo, G.L. CK2 and PI3K are direct molecular targets of quercetin in chronic lymphocytic leukaemia. Oncotarget 2017, 8, 42571–42587. [Google Scholar] [CrossRef] [Green Version]
- Gulati, N.; Laudet, B.; Zohrabian, V.M.; Murali, R.; Jhanwar-Uniyal, M. The antiproliferative effect of Quercetin in cancer cells is mediated via inhibition of the PI3K-Akt/PKB pathway. Anticancer Res. 2006, 26, 1177–1181. [Google Scholar]
- Bhattacharya, S.; McElhanon, K.E.; Gushchina, L.V.; Weisleder, N. Role of phosphatidylinositol-4,5-bisphosphate 3-kinase signaling in vesicular trafficking. Life Sci. 2016, 167, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Nomura, M.; Takahashi, T.; Nagata, N.; Tsutsumi, K.; Kobayashi, S.; Akiba, T.; Yokogawa, K.; Moritani, S.; Miyamoto, K. Inhibitory mechanisms of flavonoids on insulin-stimulated glucose uptake in MC3T3-G2/PA6 adipose cells. Biol. Pharm. Bull. 2008, 31, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, Y.; Suzuki, R. FcεRI: A Master Regulator of Mast Cell Functions. Cells 2022, 11, 622. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ilarraza, R.; Tancowny, B.P.; Alam, S.B.; Kulka, M. Disrupted Lipid Raft Shuttling of FcεRI by n-3 Polyunsaturated Fatty Acid Is Associated With Ligation of G Protein-Coupled Receptor 120 (GPR120) in Human Mast Cell Line LAD2. Front. Nutr. 2020, 7, 597809. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Khanbabapour Sasi, A.; Taheri, M.; Ayatollahi, S.A. The impact of the phytotherapeutic agent quercetin on expression of genes and activity of signaling pathways. Biomed. Pharmacother. 2021, 141, 111847. [Google Scholar] [CrossRef] [PubMed]
- Keshvara, L.M.; Isaacson, C.C.; Yankee, T.M.; Sarac, R.; Harrison, M.L.; Geahlen, R.L. Syk- and Lyn-dependent phosphorylation of Syk on multiple tyrosines following B cell activation includes a site that negatively regulates signaling. J. Immunol. 1998, 161, 5276–5283. [Google Scholar]
- Mansueto, M.S.; Reens, A.; Rakhilina, L.; Chi, A.; Pan, B.S.; Miller, J.R. A reevaluation of the spleen tyrosine kinase (SYK) activation mechanism. J. Biol. Chem. 2019, 294, 7658–7668. [Google Scholar] [CrossRef]
- Kirshenbaum, A.S.; Akin, C.; Wu, Y.; Rottem, M.; Goff, J.P.; Beaven, M.A.; Rao, V.K.; Metcalfe, D.D. Characterization of novel stem cell factor responsive human mast cell lines LAD 1 and 2 established from a patient with mast cell sarcoma/leukemia; activation following aggregation of FcepsilonRI or FcgammaRI. Leuk.Res. 2003, 27, 677–682. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| FcεRI α subunit | gttagcagtccctcagaaacc | ctgccattgtggaaccatttg |
| FcεRI β subunit FcεRI γ subunit Glyceraldegyde-3-phosphate dehydrogenase (GapDH) | aaatcttgctctcccacagg actgaagatccaagtgcgaaag agccacatcgctcagacac | ggatgaggccgacttcaatag agtctcgtaagtctcctggttc gcccaatacgaccaaatcc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, S.B.; Wagner, A.; Willows, S.; Kulka, M. Quercetin and Resveratrol Differentially Decrease Expression of the High-Affinity IgE Receptor (FcεRI) by Human and Mouse Mast Cells. Molecules 2022, 27, 6704. https://doi.org/10.3390/molecules27196704
Alam SB, Wagner A, Willows S, Kulka M. Quercetin and Resveratrol Differentially Decrease Expression of the High-Affinity IgE Receptor (FcεRI) by Human and Mouse Mast Cells. Molecules. 2022; 27(19):6704. https://doi.org/10.3390/molecules27196704
Chicago/Turabian StyleAlam, Syed Benazir, Ashley Wagner, Steven Willows, and Marianna Kulka. 2022. "Quercetin and Resveratrol Differentially Decrease Expression of the High-Affinity IgE Receptor (FcεRI) by Human and Mouse Mast Cells" Molecules 27, no. 19: 6704. https://doi.org/10.3390/molecules27196704
APA StyleAlam, S. B., Wagner, A., Willows, S., & Kulka, M. (2022). Quercetin and Resveratrol Differentially Decrease Expression of the High-Affinity IgE Receptor (FcεRI) by Human and Mouse Mast Cells. Molecules, 27(19), 6704. https://doi.org/10.3390/molecules27196704