


Effects of the Seed Oil of Carica papaya Linn on Food Consumption, Adiposity, Metabolic and Inflammatory Profile of Mice Using Hyperlipidic Diet

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Fatty Acid Composition of Papaya Seed Oil (Carica papaya Linn)
2.2. Acute Toxicity Test and “Hippocratic Screening”
2.3. Trial Period
2.3.1. Food Intake and Caloric Consumption
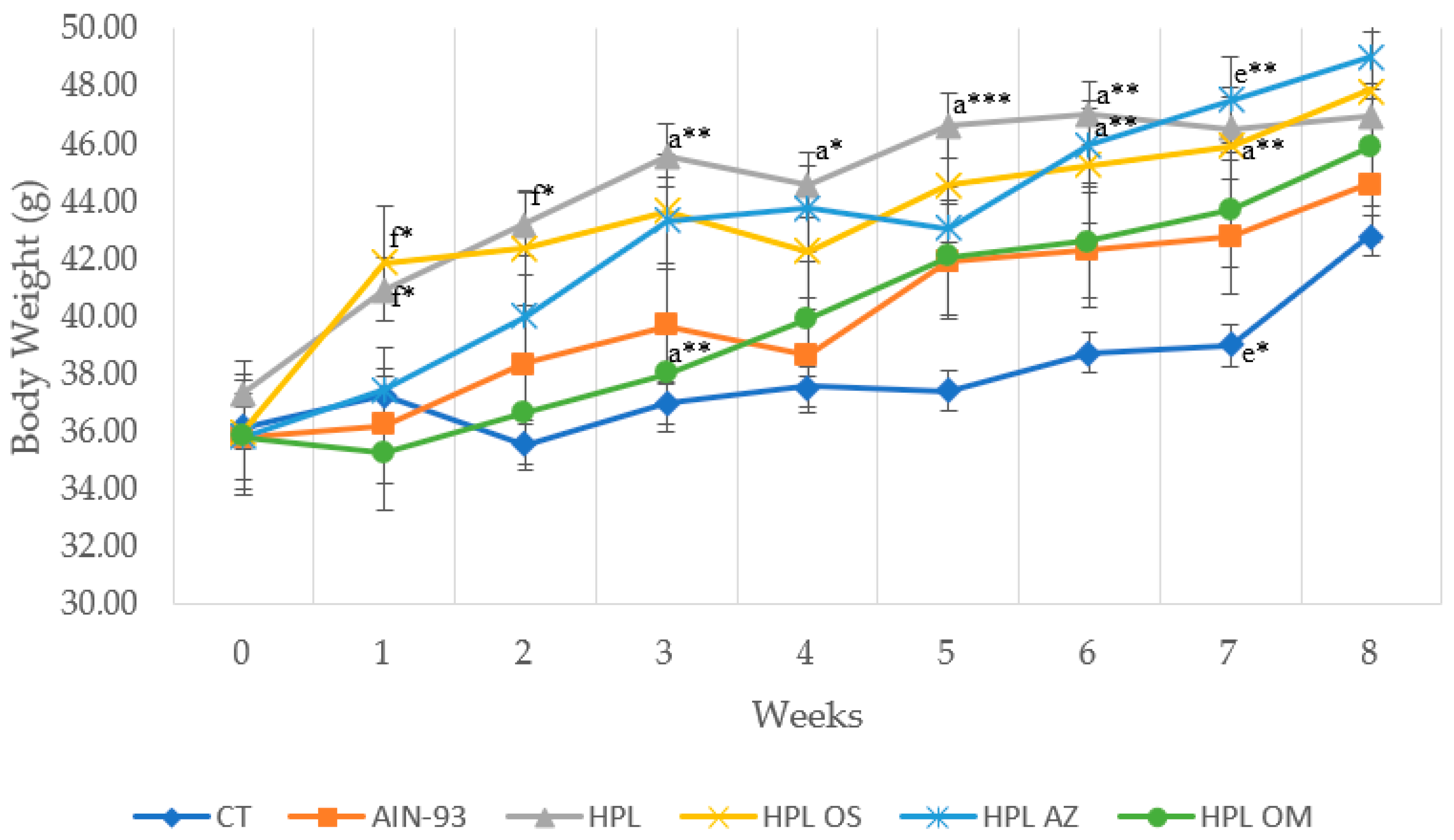
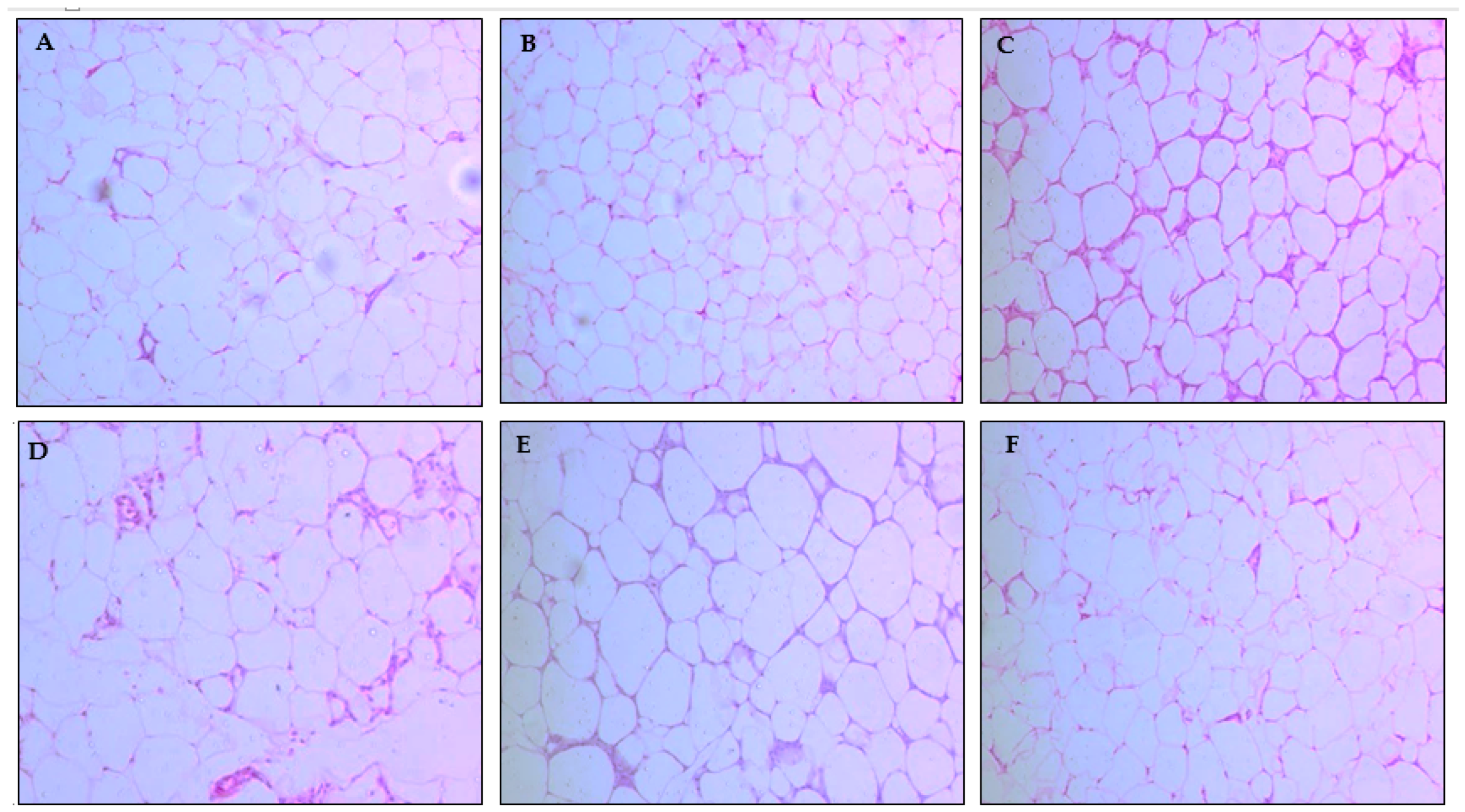
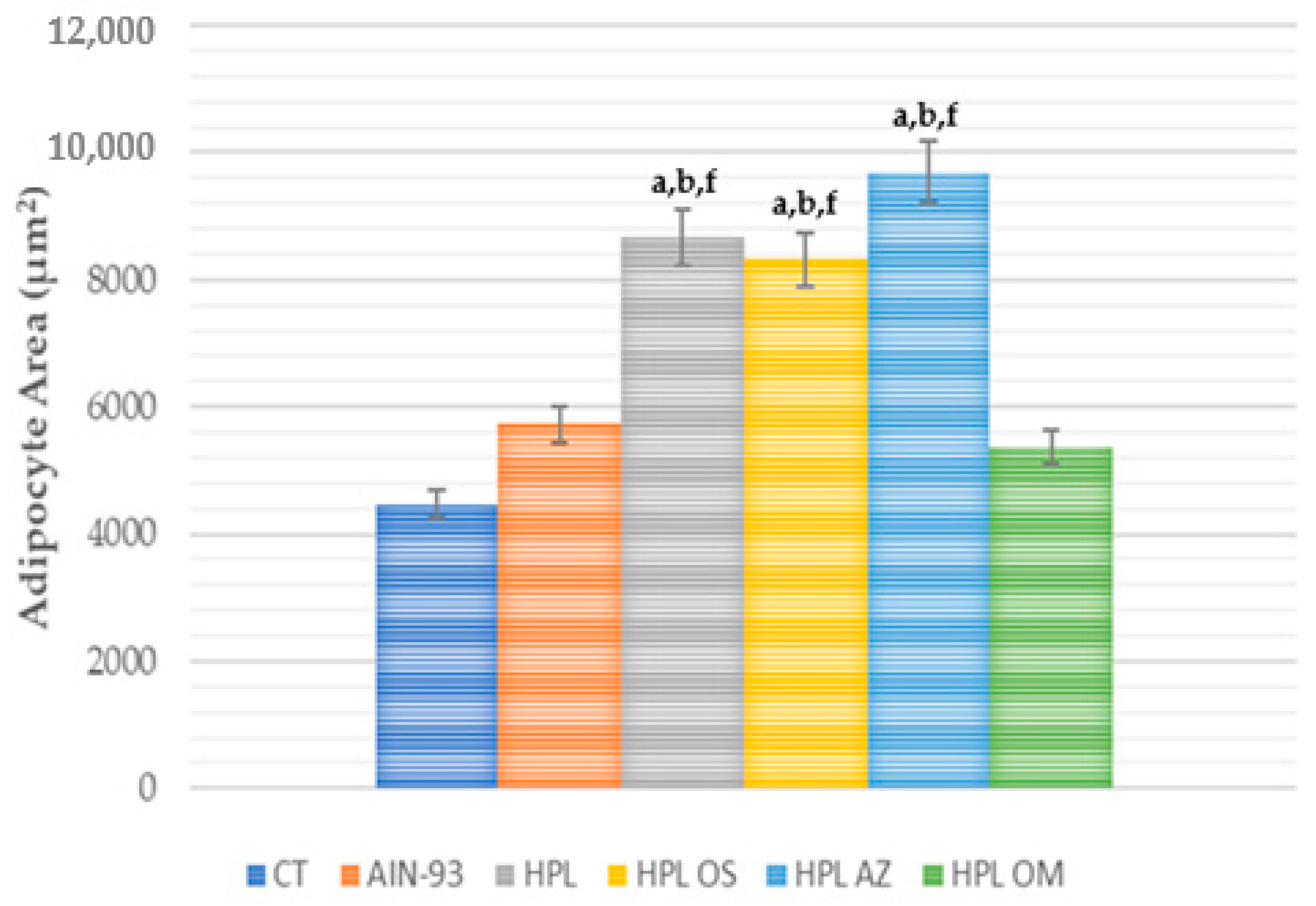
2.3.2. Assessment of Body Weight, Weight Gain, Body Fat Percentage, and Adipocyte Area
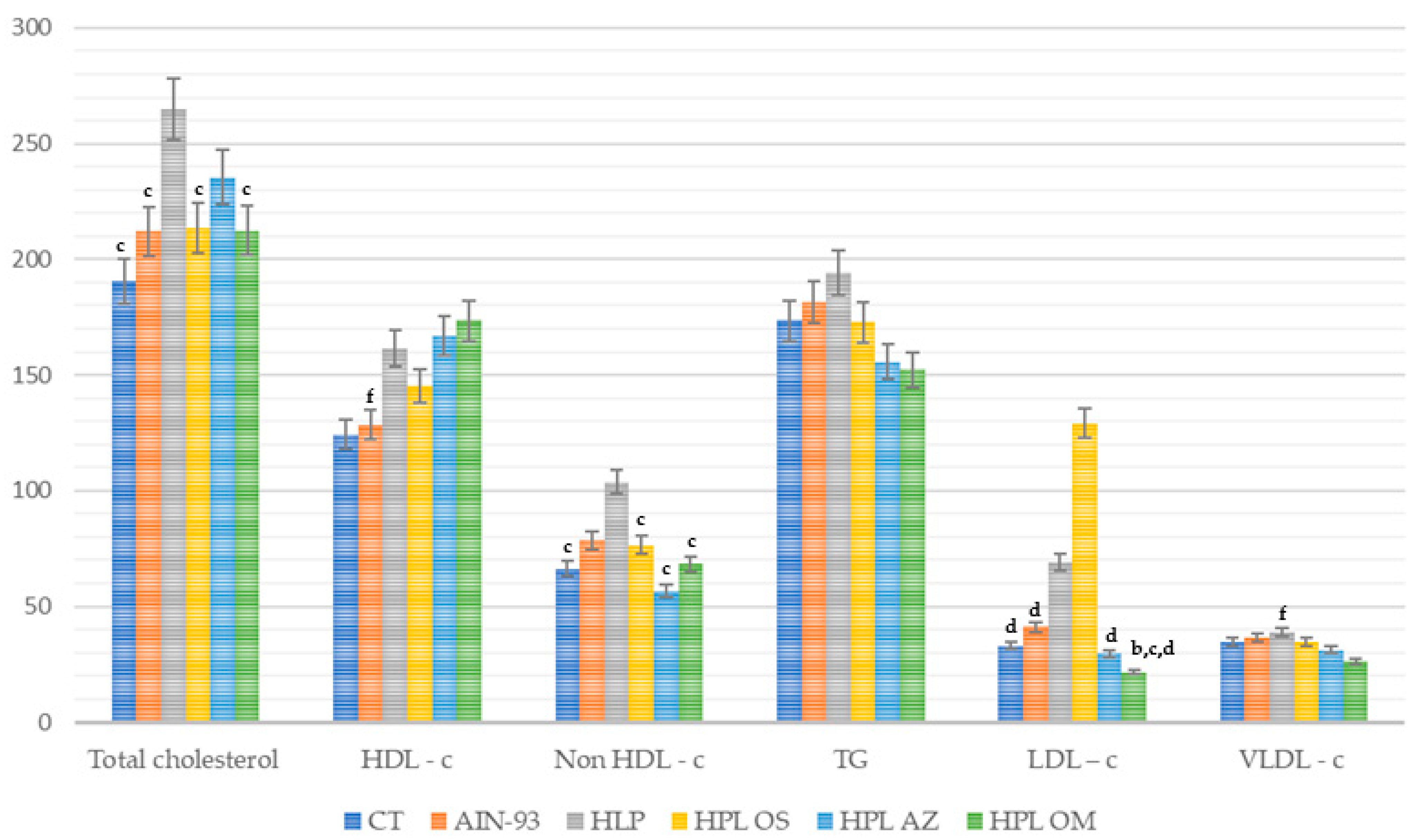
2.3.3. Serum Metabolic Changes
Triglycerides and Cholesterol (Total and Fractions) in Serum
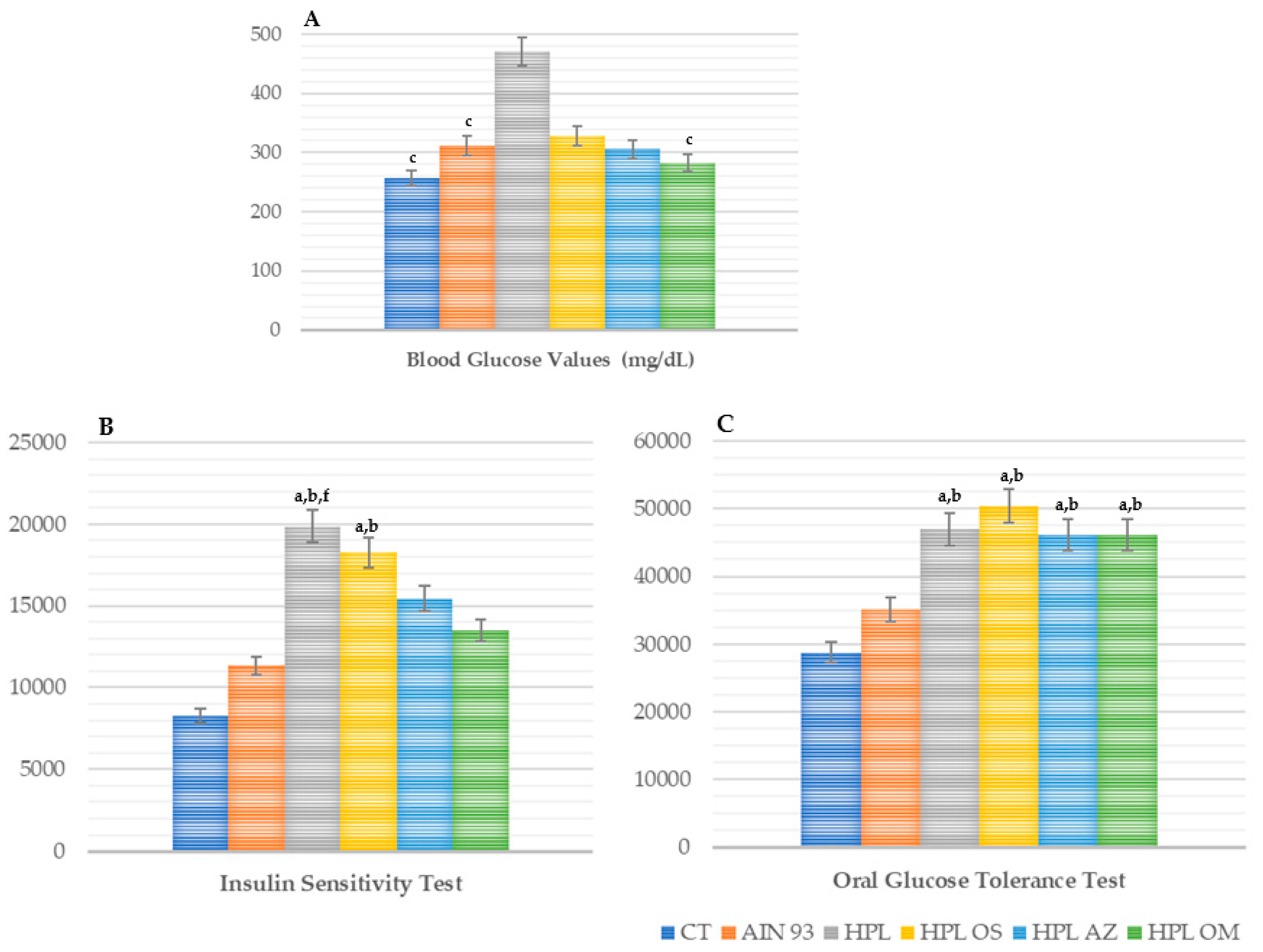
Glycemic Profile: Fasting Blood Glucose, Oral Glucose Tolerance Test, and Insulin Sensitivity
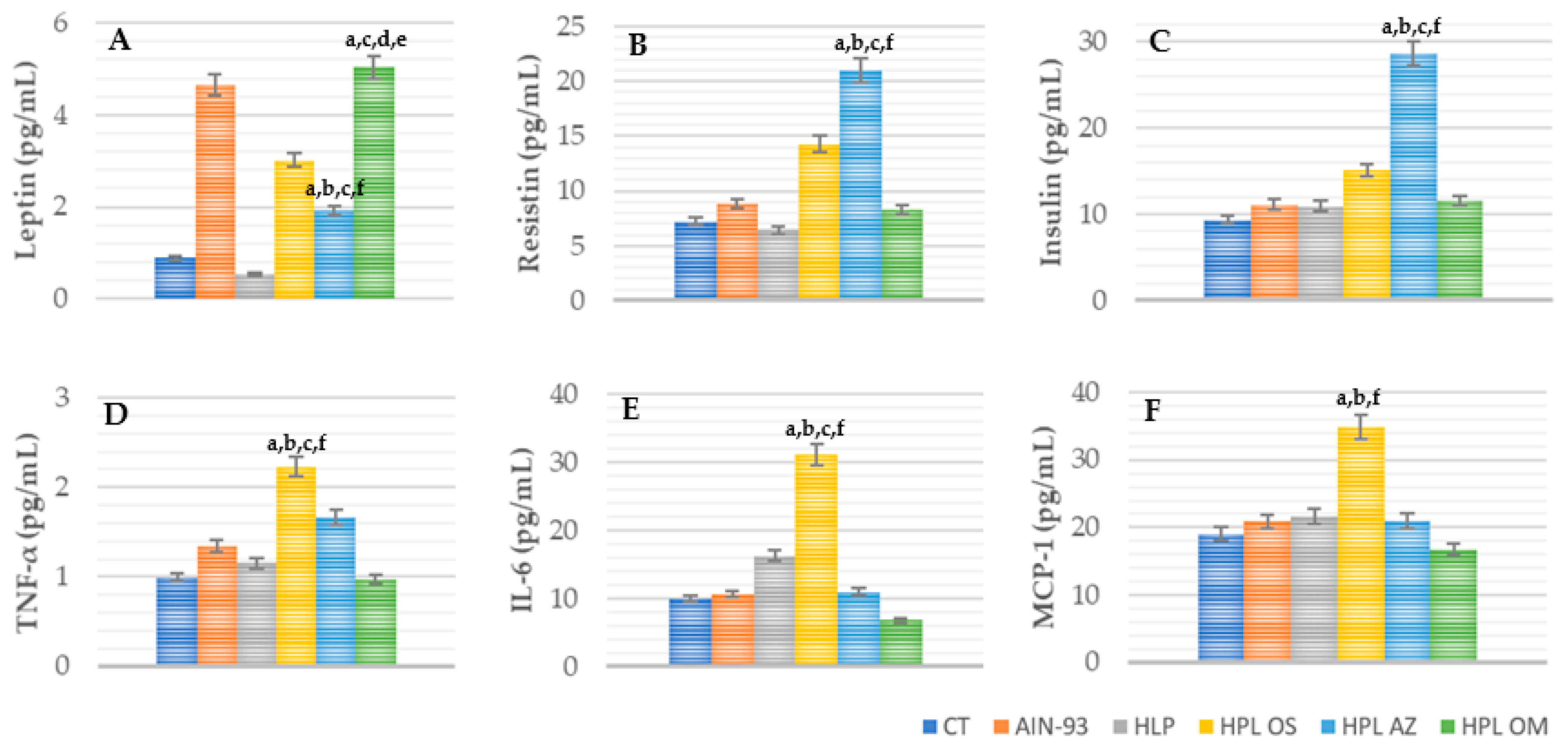
Adipokines Concentration: TNF-α, IL-6, MCP-1, Insulin, Resistin, and Leptin
2.3.4. Histological Analysis of the Liver and Pancreas
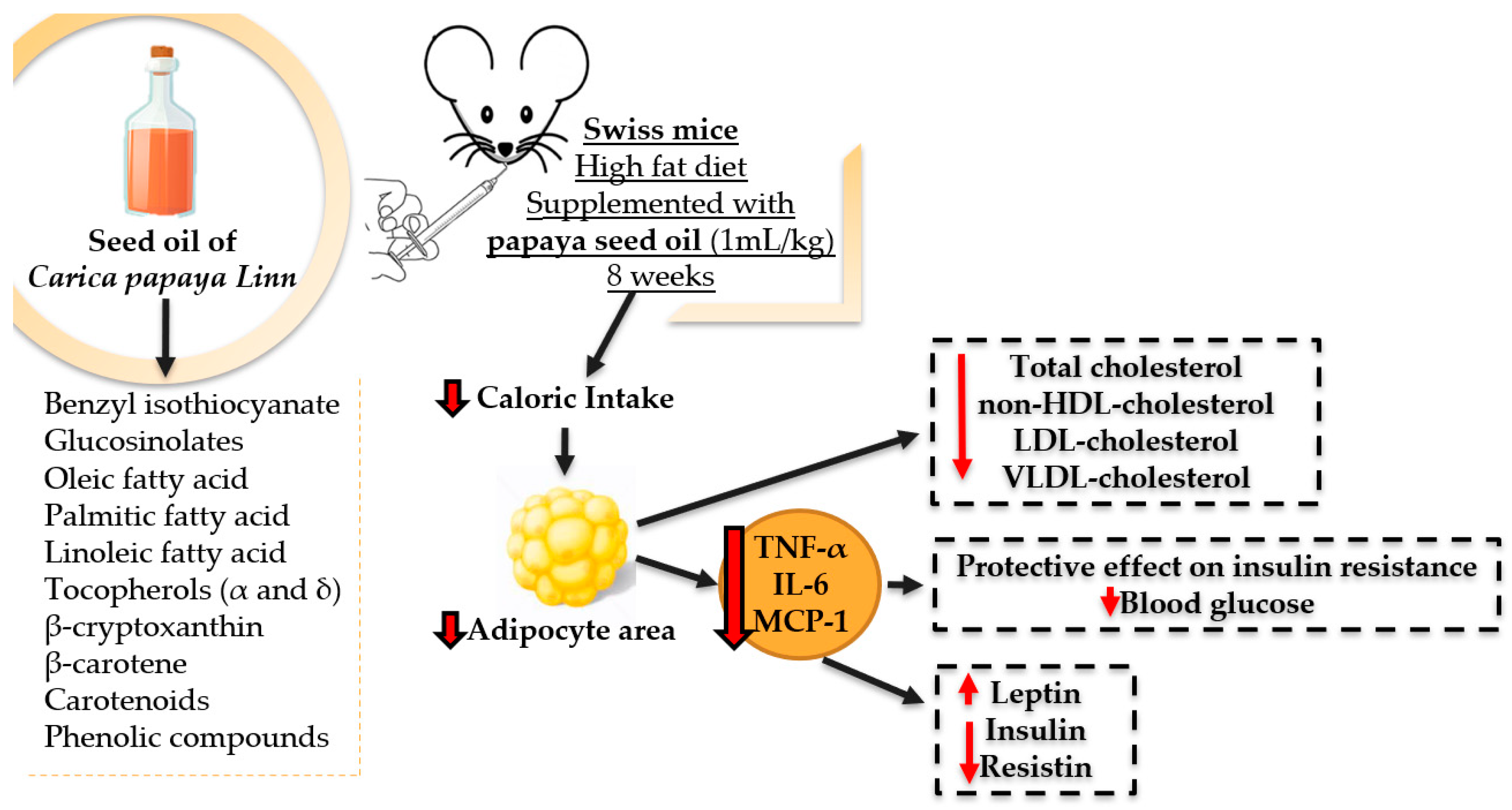
3. Discussion
4. Materials and Methods
4.1. Obtaining Oil from Papaya Seeds (Carica papaya Linn)
4.2. Fatty Acid Profile and Indices of Nutritional Quality
∑ MUFA + ∑ ω6 + ∑ ω3
0.5 × ∑ MUFA + 0.5 × ∑ ω6 + 3 × ∑ ω3
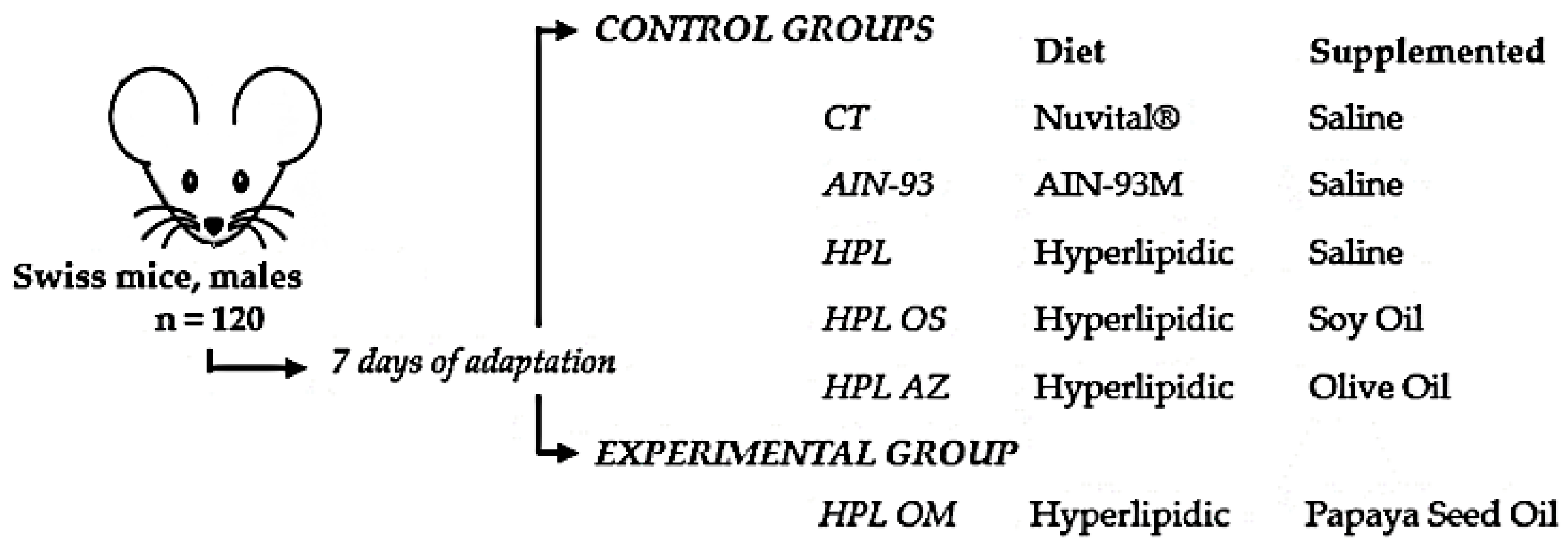
4.3. Animals
4.4. Acute Toxicity Test
4.5. Experimental Design
4.6. Metabolic Changes in Serum
4.7. Concentration of Adipokines: TNF-α, IL-6, MCP-1, Insulin, Resistin and Leptin
4.8. Histological Slides and Adipocyte Area
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Giulietti, A.M.; Harley, R.M.; De Queiroz, L.P.; Wanderley, M.D.G.L.; Van Den Berg, C. Biodiversity and Conservation of Plants in Brazil. Conserv. Biol. 2005, 19, 632–639. [Google Scholar] [CrossRef]
- Vegetabilia. McGraw-Hill Dictionary of Scientific & Technical Terms, 6E. Available online: https://encyclopedia2.thefreedictionary.com/Vegetabilia (accessed on 7 April 2022).
- Carneiro, F.M.; José, M.; Albernaz, L.C.; Darc, J.; Costa, P. Tendências dos estudos com plantas medicinais no Brasil. Rev. Sapiência 2014, 3, 44–75. [Google Scholar]
- Dutra, R.C.; Campos, M.M.; Santos, A.R.; Calixto, J.B. Medicinal plants in Brazil: Pharmacological studies, drug discovery, challenges and perspectives. Pharmacol. Res. 2016, 112, 4–29. [Google Scholar] [CrossRef] [PubMed]
- Brack, P.; Köhler, M.; Corrêa, C.A.; Ardissone, R.E.; Sobral, M.E.G.; Kinupp, V.F. Frutas nativas do Rio Grande do Sul, Brasil: Riqueza e potencial alimentício. Rodriguésia 2020, 71, 2–11. [Google Scholar] [CrossRef]
- Mello, L.D.; Pinheiro, M.F. Aspectos de azeites de oliva e de folhas de oliveira. Alim. Nutr. 2012, 23, 537–548. [Google Scholar]
- Allan, P. Effect of seeds on fruit weight in Carica papaya. Agroplantae 1969, 1, 163–170. [Google Scholar] [CrossRef]
- Rodrigues, L.G.G.; Mazzutti, S.; Vitali, L.; Micke, G.A.; Ferreira, S.R.S. Recovery of bioactive phenolic compounds from papaya seeds agroindustrial residue using subcritical water extraction. Biocatal. Agric. Biotechnol. 2019, 22, 101367. [Google Scholar] [CrossRef]
- Oloyede, O.I. Chemical profile of unripe pulp of Carica papaya. Pak. J. Nutr. 2005, 4, 379–381. [Google Scholar]
- Milind, P.; Gurditta, G. Basketful benefits of Papaya. Int. Res. J. Pharm. 2011, 1, 6–11. [Google Scholar]
- Pinto, L.A.; Cordeiro, K.W.; Carrasco, V.; Carollo, C.A.; Cardoso, C.A.L.; Argadoña, E.J.S.; Freitas, K.D.C. Antiulcerogenic activity of Caricapapaya seed in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Mariano, L.N.B.; Boeing, T.; da Silva, R.C.V.; da Silva, L.M.; Gasparotto-Júnior, A.; Cechinel-Filho, V.; de Souza, P. Exotic Medicinal Plants Used in Brazil with Diuretic Properties: A Review. Chem. Biodivers. 2022, 19, e202200258. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Kumar, S.; Mathan, S.V.; Tomar, M.S.; Singh, R.K.; Verma, P.K.; Kumar, A.; Kumar, S.; Singh, R.P.; Acharya, A. Therapeutic application of Carica papaya leaf extract in the management of human diseases. DARU J. Pharm. Sci. 2020, 28, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Vij, T.; Prashar, Y. A review on medicinal properties of Carica papaya Linn. Asian Pac. J. Trop. Dis. 2015, 5, 1–6. [Google Scholar] [CrossRef]
- Eno, A.E.; Owo, O.I.; Itam, E.H.; Konya, R.S. Blood pressure depression by the fruit juice of Carica papaya (L.) in renal and DOCA-induced hypertension in the rat. Phytother. Res. 2000, 14, 235–239. [Google Scholar] [CrossRef]
- Sadeque, M.Z.; Begum, Z.A. Protective effect of dried fruits of Carica papaya on hepatotoxicity in rat. Bangladesh J. Pharm. 2010, 5, 48–50. [Google Scholar] [CrossRef] [Green Version]
- Noor, K.A.H.; Peace, O.S.; Khadijah, A.M.H.; Nurul, A.O.; Zulhisyam, A.K.; Lee, S.W.; Mahmoud, A.O.D. Effect of papaya (Carica papaya) leaf extract as dietary growth promoter supplement in red hybrid tilapia (Oreochromis mossambicus × Oreochromis niloticus) diet. Saudi J. Biol. Sci. 2022, 29, 3911–3917. [Google Scholar] [CrossRef]
- Cortese, S.; Angriman, M.; Comencini, E.; Vincenzi, B.; Maffeis, C. Association between inflammatory cytokines and ADHD symptoms in children and adolescents with obesity: A pilot study. Psychiatry Res. 2019, 278, 7–11. [Google Scholar] [CrossRef]
- Patra, A.; Abdullah, S.; Pradhan, R.C. Review on the extraction of bioactive compounds and characterization of fruit industry by-products. Bioresour. Bioprocess. 2022, 9, 14. [Google Scholar] [CrossRef]
- Santana, L.F.; Inada, A.C.; Espirito Santo, B.L.S.D.; Filiú, W.F.; Pott, A.; Alves, F.M.; Guimarães, R.C.A.; Freitas, K.C.; Hiane, P.A. Nutraceutical potential of Carica papaya in metabolic syndrome. Nutrients 2019, 11, 1608. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.; Santos, D.; Alexandrino, S.; Oliveira, E.; Castellón, R. Obtenção e caracterização físico-química do extrato oleoso de alho roxo (Allium sativum). Tecnol. Ciência Agropecuária 2010, 4, 01–04. [Google Scholar]
- Zago, L.; Squeo, G.; Bertoncini, E.I.; Difonzo, G.; Caponio, F. Chemical and sensory characterization of Brazilian virgin olive oils. Food Res. Intern. 2019, 126, 108588. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Jacobsen, C. Oxidative Stability and Shelf Life of Foods Containing Oils and Fats, 1st ed.; Elsevier: London, UK; AOCS Press: London, UK, 2016; pp. 461–519. [Google Scholar]
- Hohmann, C.D.; Cramer, H.; Michalsen, A.; Kessler, C.; Steckhan, N.; Choi, K.; Dobos, G. Effects of high phenolic olive oil on cardiovascular risk factors: A systematic review and meta-analysis. Phytomedicine 2015, 22, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Adili, R.; Hawley, M.; Holinstat, M. Regulation of platelet function and thrombosis by omega-3 and omega-6 polyunsaturated fatty acids. Prostaglandins Other Lipid Mediat. 2019, 139, 10–18. [Google Scholar] [CrossRef]
- Bušová, M.; Kouřimská, L.; Tuček, M. Fatty acids profile, atherogenic and thrombogenic indices in freshwater fish common carp (Cyprinus carpio) and rainbow trout (Oncorhynchus mykiss) from market chain. Cent. Eur. J. Public Health 2020, 28, 313–319. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists International, 16th ed.; Patrícia Cunniff: Washington, DC, USA, 2008. [Google Scholar]
- Awodele, O.; Yemitan, O.; Ise, P.U.; Ikumawoyi, V.O. Modulatory potentials of aqueous leaf and unripe fruit extracts of Carica papaya Linn. (Caricaceae) against carbon tetrachloride and acetaminophen-induced hepatotoxicity in rats. J. Intercult. Ethnopharm. 2016, 5, 27–35. [Google Scholar] [CrossRef]
- Olagunju, J.A.; Adeneye, A.A.; Fagbohunka, B.S.; Bisuga, N.A.; Ketiku, A.O.; Benebo, A.S.; Olufowobi, O.M.; Adeoye, A.G.; Alimi, M.A.; Adeleke, A.G. Nephroprotective activities of the aqueous seed extract of Carica papaya Linn. in carbon tetrachloride induced renal injured Wistar rats: A dose-and time-dependent study. Biol. Med. 2009, 1, 11–19. [Google Scholar]
- Phillips, C.M. Metabolically healthy obesity across the life course: Epidemiology, determinants, and implications. Ann. N. Y. Acad. Sci. 2017, 1391, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.; Jeong, H.S.; Lee, J. Effect of the capsicoside G-rich fraction from pepper (Capsicum annuum L.) seeds on high-fat diet-induced obesity in mice. Phytother. Res. 2016, 30, 1848–1855. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Sakar, Y.; Covasa, M. The modulatory role of high fat feeding on gastrointestinal signals in obesity. J. Nutr. Biochem. 2013, 24, 1663–1677. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.V.; Gallão, M.I.; De Brito, E.S. Obesidade e fibra dietética: Destaque para a fibra de caju. Braz. J. Dev. 2020, 6, 43474–43488. [Google Scholar] [CrossRef]
- Campuzano-Bublitz, M.A.; Rolón, L.E.; Vera, L.M.; Kennedy, M.L. Efectodel consumo de pulpa de Carica papaya sobre la glicemia y peso de ratones normo e hiperglicémicos por aloxano. Arch. Latinoam. Nutr. 2018, 68, 132–140. [Google Scholar]
- Od-Ek, P.; Deenin, W.; Malakul, W.; Phoungpetchara, I.; Tusophon, S. Anti-obesity effect of Carica papaya in high-fat diet fed rats. Biomed. Rep. 2020, 13, 30. [Google Scholar] [CrossRef] [PubMed]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Keirsey, K.I.; Kirkland, R.; Grunewald, Z.I.; Fischer, J.G.; De La Serre, C.B. Blueberry supplementation influences the gut microbiota, inflammation, and insulin resistance in high-fat-diet–fed rats. J. Nutr. 2018, 148, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, K.S. Antioxidant and immunostimulant effect of Carica papaya Linn. aqueous extract in acrylamide intoxicated rats. Acta Inform. Med. 2012, 20, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Aschar, N.; Naqvi, S.A.; Hussain, Z.; Rasool, N.; Khan, Z.A.; Shahzad, S.A.; Sherazi, T.A.; Janjua, M.R.; Nagra, S.A.; Zia-Ul-Haq, M.; et al. Compositional difference in antioxidant and antibacterial activity of all parts of the Carica papaya using different solvents. Chem. Cent. J. 2016, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Zetina-Esquivel, A.M.; Tovilla-Zárate, C.A.; Guzman-Garcia, C. Effect of Carica papaya leaf extract on serum lipids and liver metabolic parameters of rats fed a high cholesterol diet. Health 2015, 7, 1196–1205. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, oxidative stress, adipose tissue dysfunction, and the associated health risks: Causes and therapeutic strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.P.; Lage, M.J.; Mo, D.; Nelson, D.R.; Hoogwerf, B.J. Obesity and glycemic control in patients with diabetes mellitus: Analysis of physician electronic health records in the US from 2009–2011. J. Diabetes Complicat. 2016, 30, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.M.; Islam, A.; Bari, M.W.; Hossain, M.I.; Abdul, M. Hypoglycemic and hypolipidemic activities of Carica papaya fruits in streptozotocin induced diabetic mice. WJPPS 2019, 8, 332–343. [Google Scholar] [CrossRef]
- Ezekwe, S.A.; Chikezie, P.C. GC-MS analysis, hypoglycemic activity of aqueous root extract of Carica papaya and its effects on blood lipid profile and hepatorenal tissues biomarkers of diabetic rats. J. Diabetes Metab. 2017, 8, 2234–2248. [Google Scholar] [CrossRef]
- Agada, R.; Usman, W.A.; Shehu, S.; Thagariki, D.; Agada, R. In vitro and in vivo inhibitory effects of Carica papaya seed on α-amylase and α-glucosidase enzymes. Heliyon 2020, 6, e03618. [Google Scholar] [CrossRef]
- McGregor, G.; Harvey, J. Regulation of hippocampal synaptic function by the metabolic hormone, leptin: Implications for health and neurodegenerative disease. Front. Cell. Neurosci. 2018, 12, 340. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Odle, A.K.; Haney, A.; Allensworth-James, M.; Akhter, N.; Childs, G.V. Adipocyte versus pituitary leptin in the regulation of pituitary hormones: Somatotropes develop normally in the absence of circulating leptin. Endocrinology 2014, 155, 4316–4328. [Google Scholar] [CrossRef] [Green Version]
- Moehlecke, M.; Canani, L.H.; Trindade, M.R.M.; Friedman, R.; Leitão, C.B. Determinants of body weight regulation in humans. Arch. Endocrinol. Metab. 2016, 60, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.W.; Cannon, B.; Nedergaard, J. Leptin: Is It Thermogenic? Endocr. Ver. 2020, 41, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, N.; Zhu, Y.; Straub, L.; Zhang, Z.; Wang, M.Y.; Scherer, P.E. Partial leptin deficiency confers resistance to diet-induced obesity in mice. Mol. Metab. 2020, 37, 100995. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Kant, S.; Pandey, S.; Ehtesham, N.Z. Resistin in metabolism, inflammation, and disease. FEBS J. 2020, 287, 3141–3149. [Google Scholar] [CrossRef] [Green Version]
- Challet, E. The circadian regulation of food intake. Nat. Rev. Endocrinol. 2019, 15, 393–405. [Google Scholar] [CrossRef]
- Vugic, L.; Colson, N.; Nikbakht, E.; Gaiz, A.; Holland, O.J.; Kundur, A.R.; Singh, I. Anthocyanin supplementation inhibits secretion of pro-inflammatory cytokines in overweight and obese individuals. J. Funct. Foods 2020, 64, 103596. [Google Scholar] [CrossRef]
- Parnell, J.A.; Klancic, T.; Reimer, R.A. Oligofructose decreases serum lipopolysaccharide and plasminogen activator inhibitor-1 in adults with overweight/obesity. Obesity 2017, 25, 510–513. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, K.F.; Pietrani, N.T.; Bosco, A.A.; Campos, F.M.F.; Sandrim, V.C.; Gomes, K.B. IL-6, TNF-α, and IL-10 levels/polymorphisms and their association with type 2 diabetes mellitus and obesity in Brazilian individuals. Arch. Endocrinol. Metab. 2017, 61, 438–446. [Google Scholar] [CrossRef] [Green Version]
- Somanah, J.; Bourdon, E.; Bahorun, T. Extracts of Mauritian Carica papaya (var. solo) protect SW872 and HepG2 cells against hydrogen peroxide induced oxidative stress. J. Food Sci. Technol. 2017, 54, 1917–1927. [Google Scholar] [CrossRef]
- Aravind, G.; Bhowmik, D.; Duraivel, S.; Harish, G. Traditional and medicinal uses of Carica papaya. J. Med. Plants Stud. 2013, 1, 7–15. [Google Scholar]
- Drehmer, E.; Navarro-Moreno, M.Á.; Carrera, S.; Villar, V.M.; Moreno, M.L. Oxygenic metabolism in nutritional obesity induced by olive oil. The influence of vitamin C. Food Funct. 2019, 10, 3567–3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Wang, L.; Yang, D.; Li, L.; Togo, J.; Wu, Y.; Speakman, J.R. Dietary fat, but not protein or carbohydrate, regulates energy intake and causes adiposity in mice. Cell Metab. 2018, 28, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Manzano, A.B. Distribuição, Taxa de Entrada, Composição Química e Identificação de Fontes de Grânulos Plásticos na Enseada de Santos, SP, Brasil. Tese de Mestrado, Instituto Oceanográfico, Universidade de São Paulo, São Paulo, Brazil, 2009. [Google Scholar]
- Maia, E.L.; Rodriguez-Amaya, D.B. Avaliação de um método simples e econômico para a metilação de ácidos graxos com lipídios de diversas espécies de peixes. Rev. Inst. Adolfo Lutz. 1993, 53, 27–35. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Silva, N.R. Enriquecimento da Carne Suína Com Blends de Óleos: Estudo dos Parâmetros Sanguíneos, Perfil dos Ácidos Graxos e Índices Trombogênicos e Aterogênicos. Tese de Mestrado, Universidade Federal de Lavras, Lavras, Brazil, 2018. [Google Scholar]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, A.R.S. Manual de Ensaios Toxicológicos In Vivo; Ciências Médicas; Unicamp: Campinas, Brazil, 1994; pp. 15–22. [Google Scholar]
- Lenquiste, S.A.; da Silva Marineli, R.; Moraes, É.A.; Dionísio, A.P.; de Brito, E.S.; Junior, M.R.M. Jaboticaba peel and jaboticaba peel aqueous extract shows in vitro and in vivo antioxidant properties in obesity model. Food Res. Int. 2015, 77, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Qu, L.; Liu, Q.; Zhang, Q.; Tuo, X.; Fan, D.; Deng, J.; Yang, H. Kiwifruit seed oil prevents obesity by regulating inflammation, thermogenesis, and gut microbiota in high-fat diet-induced obese C57BL/6 mice. Food Chem. Toxicol. 2019, 125, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Santana, L.F.; Santo, B.L.S.E.; Sasso, S.; Aquino, D.F.S.; Cardoso, C.A.L.; Pott, A.; Soares, F.L.P.; Freitas, K.C. Efeitos do extrato etanólico das folhas de cagaiteira (Eugenia dysenterica DC.) em camundongos diabéticos induzidos por estreptozotocina. Int. J. Dev. Res. 2018, 8, 1355–1362. [Google Scholar]
- Taylor, B.A.; Phillips, S.J. Detection of obesity QTLs on mouse chromosomes 1 and 7 by selective DNA pooling. Genomics 1996, 34, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Mello, E.S.; Alves, V.A. Atlas de Patologia Hepática; Manole: São Paulo, Brazil, 2010. [Google Scholar]
- Tai, M.M. A mathematical model for the determination of total area under glucose tolerance and other metabolic curves. Diabetes Care 1994, 17, 152–154. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W. Nonalcoholic Steatohepatitis Clinical Research Network. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Pereira, F.L.; Vasques, F.T.; Moricz, A.D.; Campos, T.D.; Pacheco, A.M., Jr.; Silva, R.A. Correlation analysis between post-pancreatoduodenectomy pancreatic fistula and pancreatic histology. Rev. Col. Bras. Cir. 2012, 39, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.S.; Teixeira, L.G.; Aguilar, E.C.; Matoso, R.O.; Soares, F.L.P.; Ferreira, A.V.M.; Alvarez-Leite, J.I. Differences in adipose tissue inflammation and oxidative status in C57BL/6 andApoE-/- mie fed high fat diet. Anim. Sci. J. 2012, 83, 549–555. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Carica papaya Linn Seed Oil (%) * |
|---|---|
| Palmitic acid (C16:0) | 16.95 ± 0.09 |
| Stearic acid (C18:0) | 3.62 ± 0.01 |
| Σ SATURATED (SFA) | 20.57 |
| Palmitoleic acid (C16:1ω7) | 0.34 ± 0.01 |
| Vacenic acid (C18:1ω7) | 0.87 ± 0.01 |
| Oleic acid (C18:1ω9) | 72.93 ± 0.01 |
| Gadoleic acid (C20:1ω9) | 0.33 ± 0.01 |
| Erucic acid (C22:1ω9) | 0.25 ± 0.01 |
| Σ MONOUNSATURATED (MUFA) | 74.75 |
| Linoleic acid (C18:2ω6) | 3.64 ± 0.02 |
| Alpha-linolenic acid (C18:3ω3) | 0.31 ± 0.01 |
| Eicosadienoic acid (C20:2ω6) | 0.21 ± 0.01 |
| Σ POLY-UNSATURATED (PUFA) | 4.16 |
| Atherogenicity index | 0.78 |
| Thrombogenicity index | 0.01 |
| ω6:ω3 | 12.42 |
| Groups | Food Intake (g) | Caloric Consumption (cal) | ||||
|---|---|---|---|---|---|---|
| First Month | Second Month | Total | First Month | Second Month | Total | |
| CT | 170.59 ± 1.80 | 153.71 ± 6.19 | 324.30 ± 24.25 | 743.80 ± 10.50 | 670.20 ± 35.50 | 1414.00 ± 37.30 |
| AIN-93 | 128.24 ± 3.34 | 119.48 ± 3.98 | 247.72 ± 18.06 a,* | 487.40 ± 14.90 a | 454.10 ± 19.10 a,c | 941.50 ± 20.20 a |
| HPL | 104.99 ± 1.81 | 107.37 ± 2.08 a,* | 212.36 ± 14.61 a,* | 556.50 ± 12.40 a | 569.10 ± 15.70 a | 1125.60 ± 14.80 a |
| HPL OS | 92.53 ± 0.66 | 95.87 ± 1.76 a,* | 188.40 ± 12.87 a,b,** | 490.50 ± 5.10 a | 508.20 ± 14.4 a,c | 998.70 ± 13.70 a |
| HPL AZ | 84.17 ± 2.03 a,* | 85.50 ± 2.14 a,* | 169.67 ± 11.73 a,b,** | 446.20 ± 15.10 a,c | 453.30 ± 18.40 a,c | 899.50 ± 18.00 a,c |
| HPL OM | 87.07 ± 2.28 a,* | 81.91 ± 1.47 a,b,c,* | 168.98 ± 12.24 a,b,** | 461.50 ± 15.20 a,c | 434.20 ± 14.60 a,c | 895.70 ± 11.90 a,c |
| Groups | Adipose Tissue Sites (mg) | % Body Fat | ||||
|---|---|---|---|---|---|---|
| Epididymal | Perirenal | Mesenteric | Retroperitoneal | Omental | ||
| CT | 1,275.42 ± 814.65 | 158.72 ± 127.02 | 537.25 ± 342.19 e | 410.95 ± 246.33 | 33.92 ± 15.85 e | 5.63 d,e |
| AIN-93 | 1584.08 ± 646.90 | 252.40 ± 184.67 | 748.53 ± 375.13 | 607.79 ± 270.90 | 31.00 ± 15.61 e | 7.52 |
| HPL | 1748.35 ± 742.81 | 322.37 ± 156.38 | 1,002.62 ± 613.01 | 612.06 ± 337.03 | 36.41 ± 20.84 e | 8.56 |
| HPL OS | 2319.62 ± 831.44 a | 284.56 ± 103.50 a | 1,114.27 ± 430.42 | 799.10 ± 251.65 a | 46.96 ± 14.97 | 10.65 |
| HPL AZ | 2122.54 ± 708.52 a | 323.46 ± 152.15 a | 1,124.91 ± 490.63 | 788.41 ± 255.67 a | 62.10 ± 27.74 | 10.32 |
| HPL OM | 2305.33 ± 722.87 a | 270.70 ± 96.15 a | 862.75 ± 334.07 | 719.45 ± 215.80 a | 41.25 ± 13.43 e | 7.37 |
| Evaluated Parameters | CT | AIN-93 | HPL | HPL OS | HPL AZ | HPL OM | p | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | n | % | n | % | n | % | n | % | |||
| Steatosis | Up to 5% | 12 | 100.0 | 9 | 69.2 | 1 | 8.3 | 2 | 14.3 | 2 | 14.3 | 1 | 7.1 | <0.001 |
| 5 to 33% | - | - | 4 | 30.8 | 2 | 16.7 | 4 | 28.6 | 2 | 14.3 | 2 | 14.3 | ||
| 33 to 66% | - | - | - | - | 3 | 25.0 | 4 | 28.6 | 5 | 35.7 | 7 | 50.0 | ||
| More than 66% | - | - | - | - | 6 | 50.0 a,b | 4 | 28.6 a,b | 5 | 35.7 a,b | 4 | 28.6 a,b | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Microvesicular Steatosis | Absent | 12 | 100.0 | 11 | 84.6 | 6 | 50.0 | 7 | 50.0 | 6 | 42.9 | 7 | 50.0 | 0.005 |
| Presence | - | - | 2 | 15.4 | 6 | 50.0 a,b | 7 | 50.0 a,b | 8 | 57.1 a,b | 7 | 50.0 a,b | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Lobular Inflammation | Absent | - | - | - | - | 1 | 8.3 | 2 | 14.3 | - | - | - | - | 0.443 |
| <2 focuses per 200x field | 12 | 100.0 | 12 | 92.3 | 9 | 75.0 | 11 | 78.6 | 13 | 92.9 | 11 | 78.6 | ||
| 2–4 focuses per 200x field | - | - | 1 | 12.5 | 2 | 16.7 | 1 | 7.1 | 1 | 7.1 | 3 | 21.4 | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Ballooning | Absent | 2 | 16.7 | 1 | 7.7 | 1 | 8.3 | - | - | 1 | 7.1 | - | - | 0.015 |
| Few cells | 10 | 83.3 | 8 | 61.5 | 8 | 66.7 | 8 | 57.1 | 13 | 92.9 | 13 | 92.9 | ||
| Many cells | - | - | 4 | 30.8 a,e | 3 | 25.0 a,e | 6 | 42.9 a,e | - | - | 1 | 7.1 a,e | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Mallory’s Hyaline | Absent | 12 | 100.0 | 9 | 69.2 | 10 | 83.3 | 8 | 57.1 | 13 | 92.9 | 13 | 92.9 | 0.030 |
| Presence | - | - | 4 | 30.8 a,d | 2 | 16.7 a,d | 6 | 42.9 | 1 | 7.1 a,d | 1 | 7.1 a,d | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Apoptosis | Absent | 12 | 100.0 | 13 | 100.0 | 10 | 83.3 | 13 | 92.9 | 11 | 78.6 | 13 | 92.9 | 0.305 |
| Presence | - | - | - | - | 2 | 16.7 | 1 | 7.1 | 3 | 21.4 | 1 | 7.1 | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Glycogenated core | None/rare | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | -- |
| Some | - | - | - | - | - | - | - | - | - | - | - | - | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Islet of Langerhans | Atrophy /hypotrophy | - | - | - | - | - | - | - | - | - | - | - | - | -- |
| Hypertrophy | - | - | - | - | - | - | - | - | - | - | - | - | ||
| Normal | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Pancreatic Acini | Dilated | - | - | - | - | - | - | - | - | - | - | - | - | -- |
| Necrosis | - | - | - | - | - | - | - | - | - | - | - | - | ||
| Normal | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Inflammatory Cells | Insulitis | - | - | - | - | - | - | - | - | - | - | - | - | -- |
| Perinsulitis | - | - | - | - | - | - | - | - | - | - | - | - | ||
| Absent | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Total | 12 | 100.0 | 13 | 100.0 | 12 | 100.0 | 14 | 100.0 | 14 | 100.0 | 14 | 100.0 | ||
| Ingredients (g/kg) | AIN-93M | Nuvital® | Hypercaloric |
|---|---|---|---|
| Starch | 620.692 | 725.67 | 320.692 |
| Casein (≥82% protein) | 140.00 | 40.00 | 140.00 |
| DL-methionine | - | 100.00 | -- |
| Lard | - | - | 320.00 |
| Sugar | 100.00 | - | 100.00 |
| Soy oil | 40.00 | 40.00 | 20.00 |
| Cellulose | 50.00 | 100.00 | 50.00 |
| Mineral mix * | 35.00 | 35.00 | 35.00 |
| Vitamins mix ** | 10.00 | 10.00 | 10.00 |
| L-cystine | 1.80 | 1.80 | 1.80 |
| Choline bitartrate | 2.50 | 2.50 | 2.50 |
| Tertbutyl hydroquinone | 0.008 | 0.008 | 0.008 |
| Energy (cal/kg) | 3802.80 | 3422.68 | 5302.80 |
| Carbohydrates (%) | 75.81% | 75.75% | 31.73% |
| Proteins (%) | 14.73% | 16.00% | 10.56% |
| Lipids (%) | 9.47% | 8.25% | 57.71% |
| Calories/g diet | 3.80 | 3.42 | 5.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, L.F.; do Espirito Santo, B.L.S.; Tatara, M.B.; Negrão, F.J.; Croda, J.; Alves, F.M.; de Oliveira Filiú, W.F.; Cavalheiro, L.F.; Nazário, C.E.D.; Asato, M.A.; et al. Effects of the Seed Oil of Carica papaya Linn on Food Consumption, Adiposity, Metabolic and Inflammatory Profile of Mice Using Hyperlipidic Diet. Molecules 2022, 27, 6705. https://doi.org/10.3390/molecules27196705
Santana LF, do Espirito Santo BLS, Tatara MB, Negrão FJ, Croda J, Alves FM, de Oliveira Filiú WF, Cavalheiro LF, Nazário CED, Asato MA, et al. Effects of the Seed Oil of Carica papaya Linn on Food Consumption, Adiposity, Metabolic and Inflammatory Profile of Mice Using Hyperlipidic Diet. Molecules. 2022; 27(19):6705. https://doi.org/10.3390/molecules27196705
Chicago/Turabian StyleSantana, Lidiani Figueiredo, Bruna Larissa Spontoni do Espirito Santo, Mariana Bento Tatara, Fábio Juliano Negrão, Júlio Croda, Flávio Macedo Alves, Wander Fernando de Oliveira Filiú, Leandro Fontoura Cavalheiro, Carlos Eduardo Domingues Nazário, Marcel Arakaki Asato, and et al. 2022. "Effects of the Seed Oil of Carica papaya Linn on Food Consumption, Adiposity, Metabolic and Inflammatory Profile of Mice Using Hyperlipidic Diet" Molecules 27, no. 19: 6705. https://doi.org/10.3390/molecules27196705
APA StyleSantana, L. F., do Espirito Santo, B. L. S., Tatara, M. B., Negrão, F. J., Croda, J., Alves, F. M., de Oliveira Filiú, W. F., Cavalheiro, L. F., Nazário, C. E. D., Asato, M. A., de Faria, B. B., do Nascimento, V. A., de Cássia Avellaneda Guimarães, R., de Cássia Freitas, K., & Hiane, P. A. (2022). Effects of the Seed Oil of Carica papaya Linn on Food Consumption, Adiposity, Metabolic and Inflammatory Profile of Mice Using Hyperlipidic Diet. Molecules, 27(19), 6705. https://doi.org/10.3390/molecules27196705