
The Influence of Polysaccharides/TiO2 on the Model Membranes of Dipalmitoylphosphatidylglycerol and Bacterial Lipids



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Samples and Subphases Preparation
2.2. Isolation of Lipids from Bacteria
2.3. Model Bacterial Membrane Preparation (Langmuir Films)
2.4. π-A Isotherms and Stability of the Lipid Monolayers Measurements
2.5. Measurements of the Monolayer’s Morphology
2.6. Microbiological Tests
3. Results
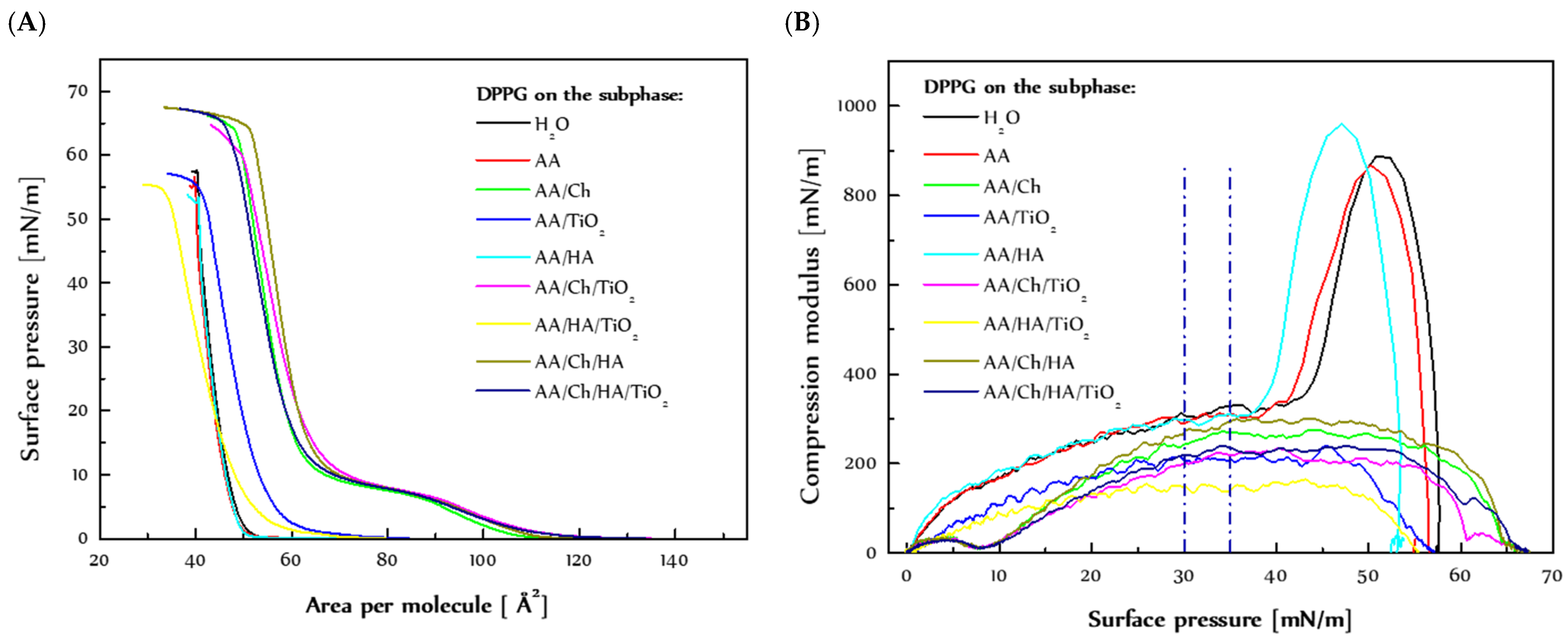
3.1. Effect of Ch, HA, TiO2 on the DPPG Model Membranes
3.1.1. Behavior of the DPPG Monolayer during Compression
3.1.2. Stability of the DPPG Monolayer
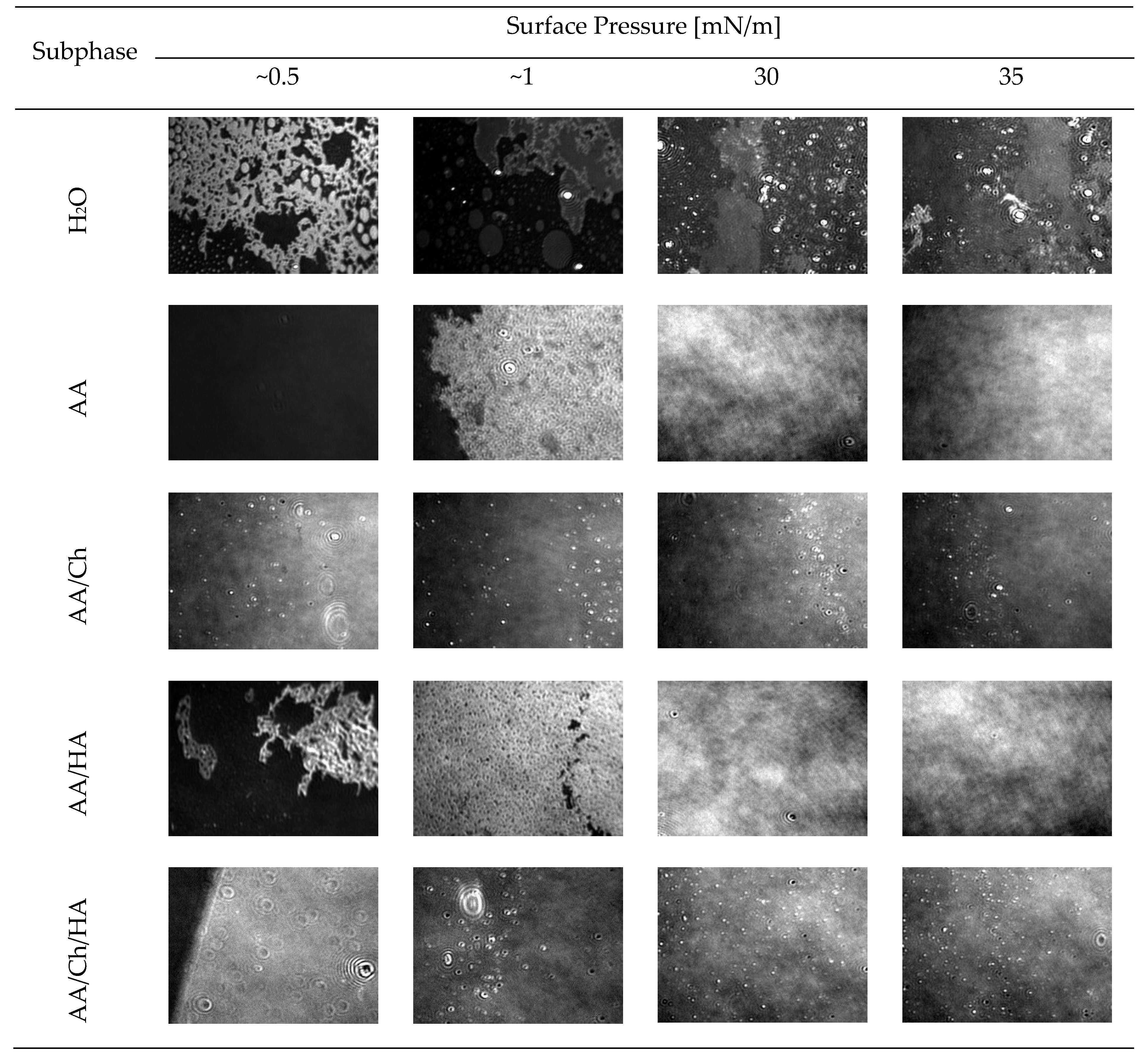
3.1.3. Thickness and Structure of the DPPG Monolayers
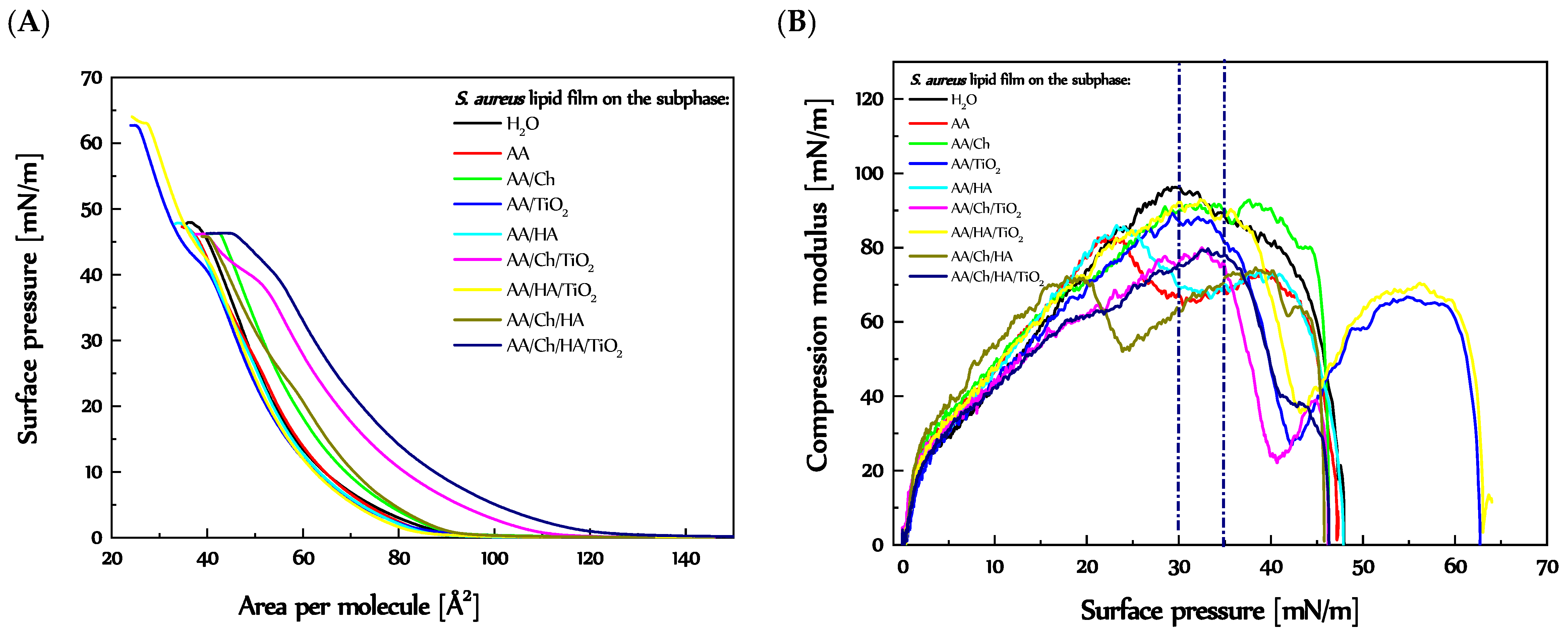
3.2. The Influence of Ch, HA and TiO2 on the S. aureus Lipids Monolayer
3.2.1. Behavior and Packing of S. aureus Lipid Monolayers during Compression
3.2.2. Stability of the S. aureus Lipid Monolayers
3.2.3. Thickness and Structure of the S. aureus Lipid Monolayers
3.3. The Influence of the Ch, HA and TiO2 on the E. coli Lipids Monolayers
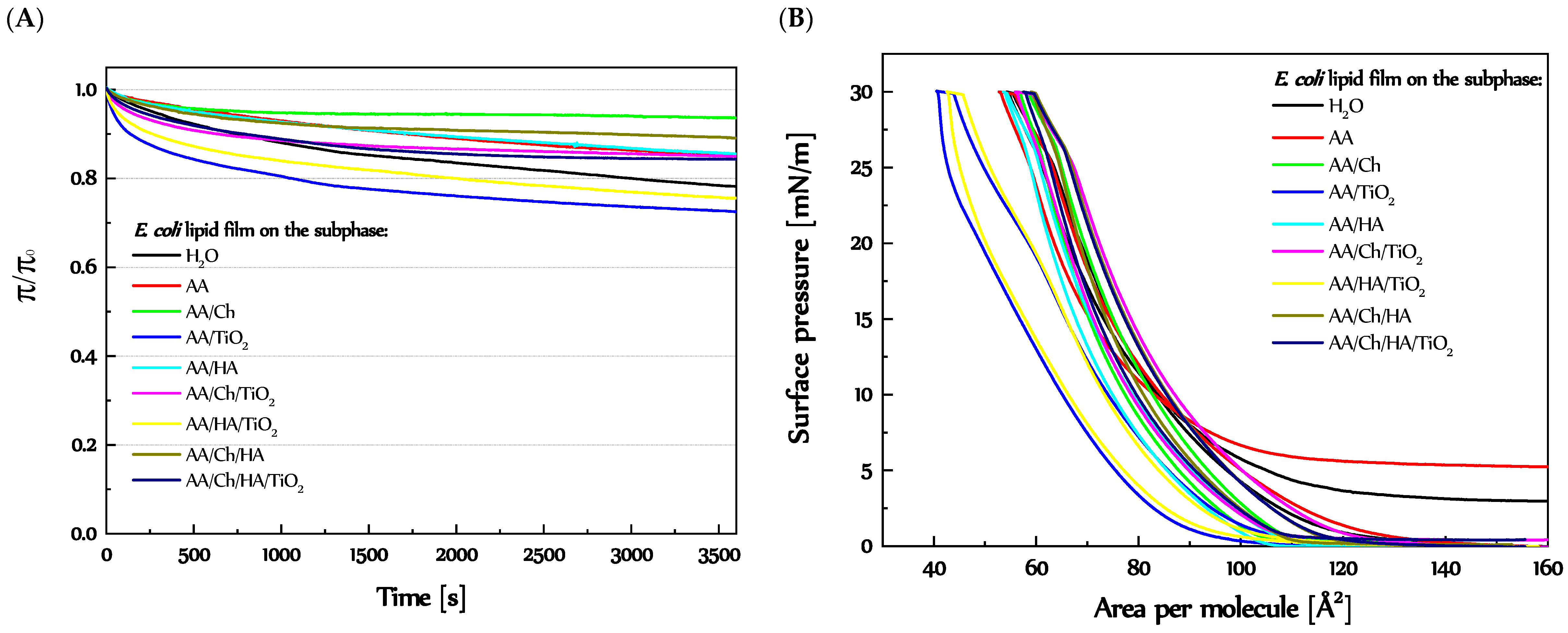
3.3.1. Behavior and Packing of E. coli Lipid Monolayers during Compression
3.3.2. Stability of the E. coli Lipid Monolayers
3.3.3. Thickness and Structure of the E. coli Lipid Monolayers
3.4. Microbiological Tests
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eeman, M.; Deleu, M. From Biological membranes to biomimetic model membranes. Biotechnol. Agron. Soc. 2010, 14, 719–736. [Google Scholar]
- Guzmán, E.; Santini, E.; Ferrari, M.; Liggieri, L.; Ravera, F. Interaction of particles with Langmuir monolayers of 1,2-dipalmitoyl-sn-glycero-3-phosphocholine: A matter of chemistry? Coatings 2020, 10, 469. [Google Scholar] [CrossRef]
- Brockman, H. Lipid monolayers: Why use half a membrane to characterize protein-membrane interactions? Curr. Opin. Struct. Biol. 1999, 9, 438–443. [Google Scholar] [CrossRef]
- Maget-Dana, R. The monolayer technique: A potent tool for studying the interfacial properties of antimicrobial and membrane-lytic peptides and their interactions with lipid membranes. Biochim. Biophys. Acta 1999, 1462, 109–140. [Google Scholar] [CrossRef] [Green Version]
- Marsh, D. Lateral pressure in membranes. Biochim. Biophys. Acta 1996, 1286, 183–223. [Google Scholar] [CrossRef]
- Chapman, M.N.; Micelles, D. Monolayers and Biomembranes; Wiley-Liss: New York, NY, USA, 1995; Volume XII, ISBN 0-471-56139-8. [Google Scholar]
- Stefaniu, C.; Brezesinski, G.; Möhwald, H. Langmuir monolayers as models to study processes at membrane surfaces. Adv. Colloid Interface Sci. 2014, 208, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, E.; Santini, E. Lung surfactant-particles at fluid interfaces for toxicity assessments. Curr. Opin. Colloid Interface Sci. 2019, 39, 24–39. [Google Scholar] [CrossRef]
- Garcia-Mouton, C.; Hidalgo, A.; Cruz, A.; Pérez-Gil, J. The lord of the lungs: The essential role of pulmonary surfactant upon inhalation of nanoparticles. Eur. J. Pharm. Biopharm. 2019, 144, 230–243. [Google Scholar] [CrossRef]
- Hoenig, D.; Moebius, D. Direct Visualization of monolayers at the air-water interface by Brewster angle microscopy. J. Phys. Chem. 1991, 95, 4590–4592. [Google Scholar] [CrossRef]
- Vollhardt, D. Brewster angle microscopy: A preferential method for mesoscopic characterization of monolayers at the air/water interface. Curr. Opin. Colloid Interface. Sci. 2014, 19, 183–197. [Google Scholar] [CrossRef]
- Quemeneur, F.; Rammal, A.; Rinaudo, M.; Pépin-Donat, B. Large and giant vesicles “decorated” with chitosan: Effects of pH, salt or glucose stress, and surface adhesion. Biomacromolecules 2007, 8, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Quemeneur, F.; Rinaudo, M.; Pépin-Donat, B. Influence of molecular weight and pH on adsorption of chitosan at the surface of large and giant vesicles. Biomacromolecules 2008, 9, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Mero, A.; Campisi, M. Hyaluronic Acid bioconjugates for the delivery of bioactive molecules. Polymers 2014, 6, 346–369. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-L.; Du, L.-C.; Weng, Y.-X.; Wang, L.; Chen, H.-Y.; Li, J.-Q. Particle-size-dependent distribution of carboxylate adsorption sites on TiO2 nanoparticle surfaces: Insights into the surface modification of nanostructured TiO2 electrodes. J. Phys. Chem. B 2004, 108, 15077–15083. [Google Scholar] [CrossRef]
- Wang, M.; Guo, Y.; Wang, Q.; Zhang, X.; Huang, J.; Lu, X.; Wang, K.; Zhang, H.; Leng, Y. Density functional theory study of interactions between glycine and TiO2/graphene nanocomposites. Chem. Phys. Lett. 2014, 599, 86–91. [Google Scholar] [CrossRef]
- Wiącek, A.E.; Gozdecka, A.; Jurak, M. Physicochemical characteristics of chitosan–TiO2 biomaterial. 1. Stability and swelling properties. Ind. Eng. Chem. Res. 2018, 57, 1859–1870. [Google Scholar] [CrossRef]
- Gozdecka, A.; Wiącek, A.E. Effect of UV radiation and chitosan coating on the adsorption-photocatalytic activity of TiO2 particles. Mater. Sci. Eng. C 2018, 93, 582–594. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, J.W. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.T. Interfacial Phenomena, 2nd ed.; Academic Press: London, UK, 1963; ISBN 978-0-323-16166-4. [Google Scholar]
- Krajewska, B.; Wydro, P.; Jańczyk, A. Probing the modes of antibacterial activity of chitosan. Effects of pH and molecular weight on chitosan interactions with membrane lipids in Langmuir films. Biomacromolecules 2011, 12, 4144–4152. [Google Scholar] [CrossRef]
- Mildner, J.; Wnętrzak, A.; Dynarowicz-Latka, P. Cholesterol and cardiolipin importance in local anesthetics–membrane interactions: The Langmuir monolayer study. J. Membr. Biol. 2019, 252, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez Patino, J.M.; Sánchez, C.C.; Rodríguez Niño, M.R. Morphological and structural characteristics of monoglyceride monolayers at the air−water interface observed by Brewster angle microscopy. Langmuir 1999, 15, 2484–2492. [Google Scholar] [CrossRef]
- Winsel, K.; Hönig, D.; Lunkenheimer, K.; Geggel, K.; Witt, C. Quantitative Brewster angle microscopy of the surface film of human broncho-alveolar lavage fluid. Eur. Biophys. J. 2003, 32, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Vollhardt, D.; Fainerman, V.B.; Siegel, S. Thermodynamic and textural characterization of DPPG phospholipid monolayers. J. Phys. Chem. B 2000, 104, 4115–4121. [Google Scholar] [CrossRef]
- Piosik, E.; Ziegler-Borowska, M.; Chełminiak-Dudkiewicz, D.; Martyński, T. Effect of aminated chitosan-coated Fe3O4 nanoparticles with applicational potential in nanomedicine on DPPG, DSPC, and POPC Langmuir monolayers as cell membrane models. Int. J. Mol. Sci. 2021, 22, 2467. [Google Scholar] [CrossRef]
- Ładniak, A.; Jurak, M.; Wiącek, A.E. Langmuir monolayer study of phospholipid DPPC on the titanium dioxide–chitosan–hyaluronic acid subphases. Adsorption 2019, 25, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Pasqui, D.; Golini, L.; Giovampaola, C.D.; Atrei, A.; Barbucci, R. Chemical and biological properties of polysaccharide-coated titania nanoparticles: The key role of proteins. Biomacromolecules 2011, 12, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Neouze, M.-A.; Schubert, U. Surface modification and functionalization of metal and metal oxide nanoparticles by organic ligands. Monatsh. Chem. 2008, 139, 183–195. [Google Scholar] [CrossRef]
- Guerrero, G.; Mutin, P.H.; Vioux, A. Anchoring of phosphonate and phosphinate coupling molecules on titania particles. Chem. Mater. 2001, 13, 4367–4373. [Google Scholar] [CrossRef]
- Michelmore, A.; Gong, W.; Jenkins, P.; Ralston, J. The interaction of linear polyphosphates with titanium dioxide surfaces. Phys. Chem. Chem. Phys. 2000, 2, 2985–2992. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Z.; Hua, W.; Castada, H.; Allen, H.C. Reorganization and caging of DPPC, DPPE, DPPG, and DPPS monolayers caused by dimethylsulfoxide observed using Brewster angle microscopy. Langmuir 2010, 26, 18902–18908. [Google Scholar] [CrossRef]
- Wydro, P.; Flasiński, M.; Broniatowski, M. Molecular organization of bacterial membrane lipids in mixed systems-A comprehensive monolayer study combined with grazing incidence X-Ray diffraction and Brewster angle microscopy experiments. Biochim. Biophys. Acta 2012, 1818, 1745–1754. [Google Scholar] [CrossRef] [Green Version]
- Torrano, A.A.; Pereira, Â.S.; Oliveira, O.N.; Barros-Timmons, A. Probing the interaction of oppositely charged gold nanoparticles with DPPG and DPPC Langmuir monolayers as cell membrane models. Colloids Surf. B Biointerfaces 2013, 108, 120–126. [Google Scholar] [CrossRef]
- Wydro, P.; Krajewska, B.; Hąc-Wydro, K. Chitosan as a lipid binder: A Langmuir monolayer study of chitosan−lipid interactions. Biomacromolecules 2007, 8, 2611–2617. [Google Scholar] [CrossRef]
- Epand, R.F.; Savage, P.B.; Epand, R.M. Bacterial lipid composition and the antimicrobial efficacy of cationic steroid compounds (ceragenins). Biochim. Biophys. Acta 2007, 1768, 2500–2509. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.M.; Epand, R.F. Domains in bacterial membranes and the action of antimicrobial agents. Mol. BioSyst. 2009, 5, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.D.; Shin, K. Effects of cardiolipin on membrane morphology: A Langmuir monolayer study. Biophys. J. 2015, 108, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Jurak, M.; Szafran, K.; Cea, P.; Martín, S. Analysis of molecular interactions between components in phospholipid-immunosuppressant-antioxidant mixed Langmuir films. Langmuir 2021, 37, 5601–5616. [Google Scholar] [CrossRef] [PubMed]
- Unsay, J.D.; Cosentino, K.; Subburaj, Y.; García-Sáez, A.J. Cardiolipin effects on membrane structure and dynamics. Langmuir 2013, 29, 15878–15887. [Google Scholar] [CrossRef]
- Wydro, P. The Influence of cardiolipin on phosphatidylglycerol/phosphatidylethanolamine monolayers—Studies on ternary films imitating bacterial membranes. Colloids Surf. B Biointerfaces 2013, 106, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Domènech, Ò.; Sanz, F.; Montero, M.T.; Hernández-Borrell, J. Thermodynamic and structural study of the main phospholipid components comprising the mitochondrial inner membrane. Biochim. Biophys. Acta 2006, 1758, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ładniak, A.; Jurak, M.; Wiącek, A. Surface characteristics of DPPC monolayers deposited from titanium dioxide-chitosan-hyaluronic acid subphases on a glass support. Prog. Chem. Appl. Chitin Deriv. 2019, 24, 106–118. [Google Scholar] [CrossRef]
- Ładniak, A.; Jurak, M.; Wiącek, A.E. Effect of chitosan, hyaluronic acid and/or titanium dioxide on the physicochemical characteristic of phospholipid film/glass surface. Physicochem. Probl. Miner. Process. 2019, 55, 1535–1548. [Google Scholar] [CrossRef]
- Ładniak, A.; Jurak, M.; Wiącek, A.E. Wettability of DPPC monolayers deposited from the titanium dioxide–chitosan–hyaluronic acid subphases on glass. Colloids Interfaces 2019, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Ładniak, A.; Jurak, M.; Wiącek, A.E. Physicochemical characteristics of chitosan-TiO2 biomaterial. 2. Wettability and biocompatibility. Colloid Surf. A-Physicochem. Eng. Asp. 2021, 630, 127546. [Google Scholar] [CrossRef]
- Sennato, S.; Bordi, F.; Cametti, C.; Coluzza, C.; Desideri, A.; Rufini, S. Evidence of domain formation in cardiolipin—Glycerophospholipid mixed monolayers. A thermodynamic and AFM Study. J. Phys. Chem. B 2005, 109, 15950–15957. [Google Scholar] [CrossRef]
- Simonson, A.W.; Lawanprasert, A.; Goralski, T.D.P.; Keiler, K.C.; Medina, S.H. Bioresponsive peptide-polysaccharide nanogels—A versatile delivery system to augment the utility of bioactive cargo. Nanomedicine 2019, 17, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Makvandi, P.; Ali, G.W.; Della Sala, F.; Abdel-Fattah, W.F.; Borzacchiello, A. Biosynthesis and characterization of antibacterial thermosensitive hydrogels based on corn silk extract, hyaluronic acid and nanosilver for potential wound healing. Carbohydr. Polym. 2019, 223, 115023. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, R.M.; Abdel-Mohsen, A.M.; Zboncak, M.; Frankova, J.; Lepcio, P.; Kobera, L.; Steinhart, M.; Pavlinak, D.; Spotaz, Z.; Sklenářévád, R.; et al. Hyaluronan biofilms reinforced with partially deacetylated chitin nanowhiskers: Extraction, fabrication, In-Vitro and antibacterial properties of advanced nanocomposites. Carbohydr. Polym. 2020, 235, 115951. [Google Scholar] [CrossRef]
- Chen, W.; Zhu, Y.; Zhang, Z.; Gao, Y.; Liu, W.; Borjihan, Q.; Qu, H.; Zhang, Y.; Zhang, Y.; Wang, Y.-J.; et al. Engineering a multifunctional N-halamine-based antibacterial hydrogel using a super-convenient strategy for infected skin defect therapy. Chem. Eng. J. 2020, 379, 122238. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subphase | DPPG Monolayer on Different Subphases | |
|---|---|---|
| A0 [Å2] | Alim [Å2] | |
| H2O | 52.5 | 48.3 |
| AA | 51.9 | 46.5 |
| AA/Ch | 107.2 | 63.5 |
| AA/TiO2 | 72.8 | 54.3 |
| AA/HA | 51.0 | 47.9 |
| AA/Ch/TiO2 | 117.0 | 67.1 |
| AA/HA/TiO2 | 68.2 | 50.9 |
| AA/Ch/HA | 110.5 | 65.3 |
| AA/Ch/HA/TiO2 | 116.4 | 64.5 |
| Surface Pressure [mN/m] | Thickness of DPPG Monolayer Obtained on Different Subphases [nm] | ||||
|---|---|---|---|---|---|
| H2O | AA | AA/Ch | AA/HA | AA/Ch/HA | |
| 0.5 | 0.3 | 2.1 | 0.9 | - | 1.1 |
| 5 | 2.5 | 2.7 | 1.5 | 2.6 | 1.6 |
| 10 | 2.6 | 2.7 | 1.4 | 2.7 | 1.9 |
| 15 | 2.7 | 2.8 | 2.7 | 2.8 | 0.9 |
| 20 | 2.7 | 2.8 | 3.5 | 2.8 | 1.8 |
| 25 | 2.8 | 2.8 | 3.7 | 2.8 | 3.6 |
| 30 | 2.8 | 2.8 | 3.8 | 2.8 | 3.9 |
| 35 | 2.8 | 2.8 | 3.8 | 2.9 | 4.0 |
| 40 | 2.8 | 2.9 | 3.9 | 2.8 | 4.0 |
| 45 | 2.8 | 2.9 | 3.9 | 2.9 | 4.1 |
| 50 | 2.8 | 2.9 | 3.9 | 2.9 | 4.1 |
| 55 | 2.8 | 2.9 | 3.9 | 2.9 | 4.1 |
| 60 | 3.8 | 4.0 | |||
| 65 | 3.5 | 3.6 | |||
| Surface Pressure [mN/m] | Thickness of S. aureus Lipid Monolayer Obtained on Different Subphases [nm] | ||||
|---|---|---|---|---|---|
| H2O | AA | AA/Ch | AA/HA | AA/Ch/HA | |
| 0.5 | 1.4 | 1.7 | 1.7 | 1.5 | 1.6 |
| 5 | 1.8 | 1.9 | 2.0 | 1.9 | 1.8 |
| 10 | 2.0 | 2.2 | 2.0 | 2.1 | 2.0 |
| 15 | 2.1 | 2.3 | 2.5 | 2.3 | 2.2 |
| 20 | 2.2 | 2.4 | 2.6 | 2.4 | 2.3 |
| 25 | 2.3 | 2.6 | 3.1 | 2.5 | 2.5 |
| 30 | 2.4 | 2.7 | 2.8 | 2.6 | 2.5 |
| 35 | 2.5 | 2.8 | 3.0 | 2.8 | 2.7 |
| 40 | 2.7 | 2.8 | 3.0 | 2.8 | 2.8 |
| 45 | 2.7 | 2.7 | 3.1 | 2.7 | 2.8 |
| Surface Pressure [mN/m] | Thickness of E. coli Monolayer Obtained on Different Subphases [nm] | ||||
|---|---|---|---|---|---|
| H2O | AA | AA/Ch | AA/HA | AA/Ch/HA | |
| 0.5 | 1.5 | 1.4 | 1.4 | 0.7 | 1.6 |
| 5 | 1.7 | 1.7 | 1.7 | 1.2 | 1.7 |
| 10 | 1.9 | 1.8 | 1.9 | 1.4 | 1.7 |
| 15 | 2.0 | 2.0 | 2.0 | 1.6 | 2.4 |
| 20 | 2.1 | 2.1 | 2.2 | 1.7 | 2.5 |
| 25 | 2.2 | 2.2 | 2.3 | 1.8 | 2.5 |
| 30 | 2.3 | 2.3 | 2.3 | 1.8 | 2.7 |
| 35 | 2.4 | 2.3 | 2.4 | 1.9 | 2.8 |
| 40 | 2.5 | 2.4 | 2.5 | 2.1 | 2.9 |
| 45 | 2.5 | 2.5 | 2.6 | 2.1 | 2.9 |
| Sample | Mortality (%) | |||
|---|---|---|---|---|
| E. coli | S. aureus | |||
| A | B | A | B | |
| AA/Ch | 96 ± 2 | 74 ± 6 | 96 ± 1 | 72 ± 7 |
| AA/TiO2 | 78 ± 4 | 58 ± 7 | 70 ± 5 | 65 ± 3 |
| AA/HA | 73 ± 5 | 49 ± 4 | 25± 4 | 35±5 |
| AA/Ch/TiO2 | 92 ± 2 | 73 ± 8 | 43 ± 1 | 41 ± 2 |
| AA/HA/TiO2 | 94 ± 2 | 74 ± 6 | 0 | 52 ± 2 |
| AA/Ch/HA | 92 ± 1 | 82 ± 5 | 62 ± 1 | 60 ± 4 |
| AA/Ch/HA/TiO2 | 93 ± 2 | 91 ± 1 | 42 ± 4 | 41 ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ładniak, A.; Jurak, M.; Palusińska-Szysz, M.; Wiącek, A.E. The Influence of Polysaccharides/TiO2 on the Model Membranes of Dipalmitoylphosphatidylglycerol and Bacterial Lipids. Molecules 2022, 27, 343. https://doi.org/10.3390/molecules27020343
Ładniak A, Jurak M, Palusińska-Szysz M, Wiącek AE. The Influence of Polysaccharides/TiO2 on the Model Membranes of Dipalmitoylphosphatidylglycerol and Bacterial Lipids. Molecules. 2022; 27(2):343. https://doi.org/10.3390/molecules27020343
Chicago/Turabian StyleŁadniak, Agata, Małgorzata Jurak, Marta Palusińska-Szysz, and Agnieszka Ewa Wiącek. 2022. "The Influence of Polysaccharides/TiO2 on the Model Membranes of Dipalmitoylphosphatidylglycerol and Bacterial Lipids" Molecules 27, no. 2: 343. https://doi.org/10.3390/molecules27020343
APA StyleŁadniak, A., Jurak, M., Palusińska-Szysz, M., & Wiącek, A. E. (2022). The Influence of Polysaccharides/TiO2 on the Model Membranes of Dipalmitoylphosphatidylglycerol and Bacterial Lipids. Molecules, 27(2), 343. https://doi.org/10.3390/molecules27020343