
Optimised Method for Short-Chain Fatty Acid Profiling of Bovine Milk and Serum

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk and Serum Samples
2.2. Chemicals
2.3. SCFA Derivatisation
2.4. LC-MS Conditions
2.5. Method Validation
2.6. Method Application
3. Results
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.; Rochfort, S.; Cocks, B. Milk lipidomics: What we know and what we don’t. Prog. Lipid Res. 2018, 71, 70–85. [Google Scholar] [CrossRef]
- Luke, T.D.W.; Rochfort, S.; Wales, W.J.; Bonfatti, V.; Marett, L.; Pryce, J.E. Metabolic profiling of early-lactation dairy cows using milk mid-infrared spectra. J. Dairy Sci. 2019, 102, 1747–1760. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Stoop, W.M. Genetic Variation in Bovine Milk Fat Composition. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2009. [Google Scholar]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- Shen, H.; Xu, Z.; Shen, Z.; Lu, Z. The regulation of ruminal short-chain fatty acids on the functions of rumen barriers. Front. Physiol. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Van Knegsel, A.T.M.; van den Brand, H.; Dijkstra, J.; Tamminga, S.; Kemp, B. Effect of dietary energy source on energy balance, production, metabolic disorders and reproduction in lactating dairy cattle. Reprod. Nutr. Dev. 2005, 45, 665–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, G.; Li, Y.; Zhang, Y. Effects of high forage/concentrate diet on volatile fatty acid production and the microorganisms involved in VFA production in cow rumen. Animals 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoop, W.M.; Bovenhuis, H.; Heck, J.M.L.; van Arendonk, J.A.M. Effect of lactation stage and energy status on milk fat composition of Holstein-Friesian cows. J. Dairy Sci. 2009, 92, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ezernieks, V.; Wang, J.; Arachchillage, N.W.; Garner, J.B.; Wales, W.J.; Cocks, B.G.; Rochfort, S. Heat stress in dairy cattle alters lipid composition of milk. Sci. Rep. 2017, 7, 961. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wu, Y.; Shrestha, R.; Gao, Z.; Zhao, Y.; Miura, Y.; Tamakoshi, A.; Chiba, H.; Hui, S.-P. Determination of total, free and esterified short-chain fatty acid in human serum by liquid chromatography-mass spectrometry. Ann. Clin. Biochem. 2019, 56, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Dei Cas, M.; Paroni, R.; Saccardo, A.; Casagni, E.; Arnoldi, S.; Gambaro, V.; Saresella, M.; Mario, C.; La Rosa, F.; Marventano, I.; et al. A straightforward LC-MS/MS analysis to study serum profile of short and medium chain fatty acids. J. Chromatogr. B 2020, 1154, 121982. [Google Scholar] [CrossRef]
- Li, M.; Zhu, R.; Song, X.; Wang, Z.; Weng, H.; Liang, J. A sensitive method for the quantification of short-chain fatty acids by benzyl chloroformate derivatization combined with GC-MS. Analyst 2020, 145, 2692–2700. [Google Scholar] [CrossRef]
- He, L.; Prodhan, M.A.I.; Yuan, F.; Yin, X.; Lorkiewicz, P.K.; Wei, X.; Feng, W.; McClain, C.; Zhang, X. Simultaneous quantification of straight-chain and branched-chain short chain fatty acids by gas chromatography mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1092, 359–367. [Google Scholar] [CrossRef]
- Song, H.E.; Lee, H.Y.; Kim, S.J.; Back, S.H.; Yoo, H.J. A Facile Profiling Method of Short Chain Fatty Acids Using Liquid Chromatography-Mass Spectrometry. Metabolites 2019, 9, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotti, C.; Rubert, J.; Fava, F.; Tuohy, K.; Mattivi, F.; Vrhovsek, U. Development of a fast and cost-effective gas chromatography-mass spectrometry method for the quantification of short-chain and medium-chain fatty acids in human biofluids. Anal. Bioanal. Chem. 2017, 409, 5555–5567. [Google Scholar] [CrossRef] [PubMed]
- González-Córdova, A.F.; Vallejo-Cordoba, B. Quantitative Determination of Short-Chain Free Fatty Acids in Milk Using Solid-Phase Microextraction and Gas Chromatography. J. Agric. Food Chem. 2001, 49, 4603–4608. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.G. The composition of bovine milk lipids: January 1995 to December 2000. J. Dairy Sci. 2002, 85, 295–350. [Google Scholar] [CrossRef]
- Han, J.; Gagnon, S.; Eckle, T.; Borchers, C.H. Metabolomic analysis of key central carbon metabolism carboxylic acids as their 3-nitrophenylhydrazones by UPLC ESI-MS. Electrophoresis 2013, 34, 2891–2900. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Rochfort, S. A fast liquid chromatography-mass spectrometry (LC-MS) method for quantification of major polar metabolites in plants. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 912, 8–15. [Google Scholar] [CrossRef]
- Han, J.; Lin, K.; Sequeira, C.; Borchers, C.H. An isotope-labeled chemical derivatization method for the quantitation of short-chain fatty acids in human feces by liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2015, 854, 86–94. [Google Scholar] [CrossRef]
- Liebisch, G.; Ecker, J.; Roth, S.; Schweizer, S.; Öttl, V.; Schött, H.-F.; Yoon, H.; Haller, D.; Holler, E.; Burkhardt, R.; et al. Quantification of fecal short chain fatty acids by liquid chromatography tandem mass spectrometry-investigation of pre-analytic stability. Biomolecules 2019, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-Y.; Wang, C.; Guo, L.-X.; Zheng, Y.-F.; Hu, W.-H.; Dong, T.T.X.; Wang, T.-J.; Tsim, K.W.K. Simultaneous determination of short-chain fatty acids in human feces by HPLC with ultraviolet detection following chemical derivatization and solid-phase extraction segmental elution. J. Sep. Sci. 2019, 42, 2500–2509. [Google Scholar] [CrossRef]
- Månsson, H.L. Fatty acids in bovine milk fat. Food Nutr. Res. 2008, 52, 1821. [Google Scholar] [CrossRef] [Green Version]
- Walker, G.P.; Wijesundera, C.; Dunshea, F.R.; Doyle, P.T. Seasonal and stage of lactation effects on milk fat composition in northern Victoria. Anim. Prod. Sci. 2013, 53, 560–572. [Google Scholar] [CrossRef]
- Adamska, A.; Rutkowska, J.; Przybylski, W. Comparison of fatty acid composition of milk from Simmental and Polish Holstein-Friesian cows in different production seasons. Ann. Anim. Sci. 2016, 16, 1211–1225. [Google Scholar] [CrossRef] [Green Version]
- Fiore, E.; Tessari, R.; Morgante, M.; Gianesella, M.; Badon, T.; Bedin, S.; Mazzotta, E.; Berlanda, M. Identification of plasma fatty acids in four lipid classes to understand energy metabolism at different levels of ketonemia in dairy cows using thin layer chromatography and gas chromatographic techniques (TLC-GC). Animals 2020, 10, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puppel, K.; Gołębiewski, M.; Solarczyk, P.; Grodkowski, G.; Slósarz, J.; Kunowska-Slósarz, M.; Balcerak, M.; Przysucha, T.; Kalińska, A.; Kuczyńska, B. The relationship between plasma β-hydroxybutyric acid and conjugated linoleic acid in milk as a biomarker for early diagnosis of ketosis in postpartum Polish Holstein-Friesian cows. BMC Vet. Res. 2019, 15, 367. [Google Scholar] [CrossRef] [Green Version]


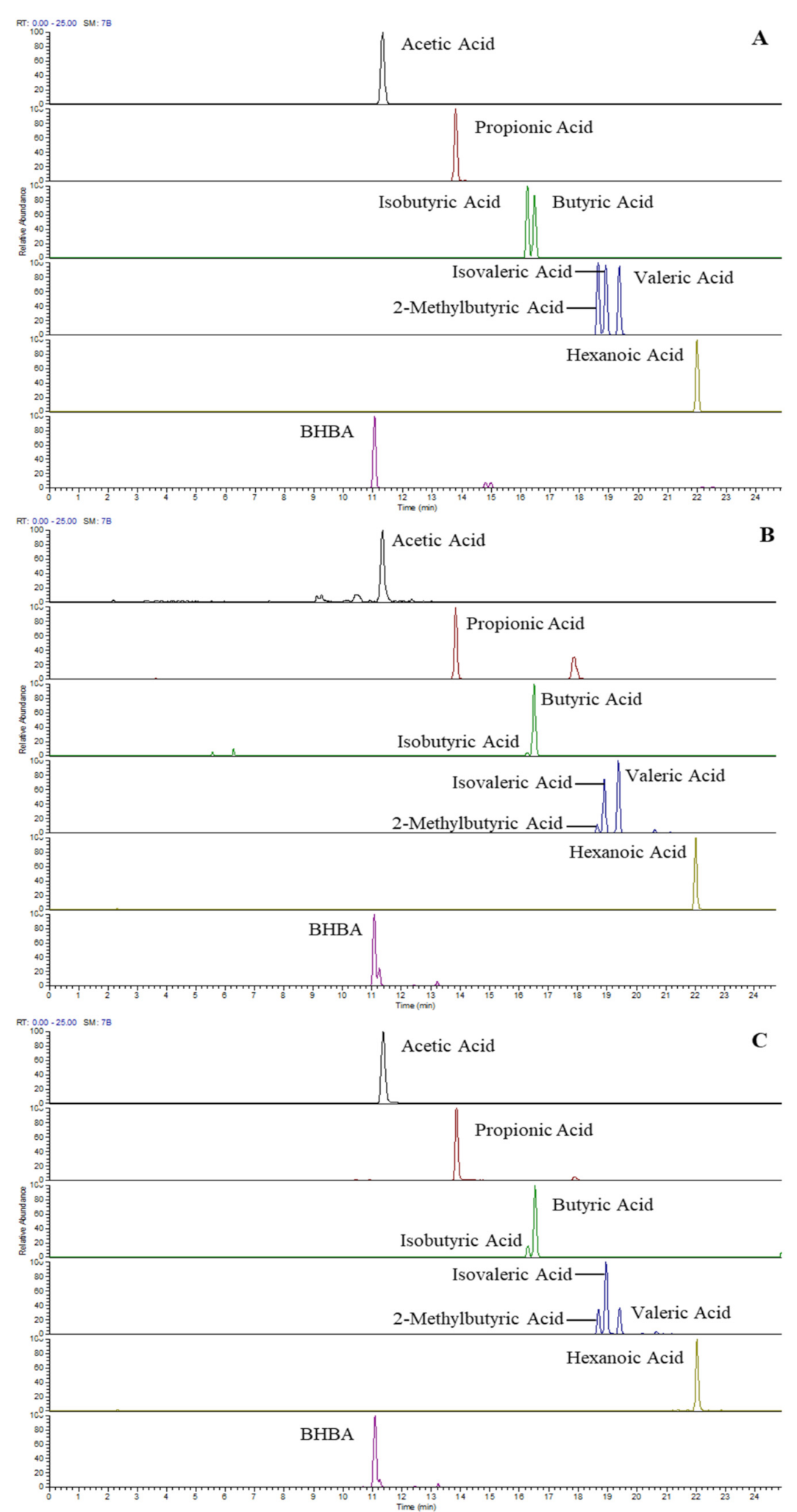{kind=link}
| Name | Accurate Mass * (m/z) | LOD (µmol/L) | LOQ (µmol/L) | Linear Range (µmol/L) | R2 |
|---|---|---|---|---|---|
| Acetic Acid | 196.0722 | 0.17 | 0.55 | 0.55–166.61 | 0.9974 |
| Propionic Acid | 210.0879 | 0.0446 | 0.14 | 0.14–44.57 | 0.9999 |
| Isobutyric Acid | 224.1035 | 0.0375 | 0.11 | 0.11–37.48 | 1.0000 |
| Butyric Acid | 224.1035 | 0.0375 | 0.11 | 0.11–37.48 | 0.9997 |
| 2-Methylbutyric Acid | 238.1192 | 0.0323 | 0.10 | 0.10–32.33 | 0.9999 |
| Isovaleric Acid | 238.1192 | 0.0323 | 0.10 | 0.10–32.33 | 0.9999 |
| Valeric Acid | 238.1192 | 0.0323 | 0.10 | 0.10–32.33 | 0.9999 |
| Hexanoic Acid | 252.1348 | 0.09 | 0.28 | 0.28–28.43 | 0.9993 |
| BHBA | 240.0984 | 0.10 | 0.32 | 0.32–96.11 | 0.9980 |
| SCFA | Milk | Serum | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample 1 | Sample 2 | Sample 3 | Bulked Sample | |||||||||
| Mean | SD | RSD | Mean | SD | RSD | Mean | SD | RSD | Mean | SD | RSD | |
| Acetic acid | 12.00 | 0.373 | 3.11 | 12.530 | 0.667 | 5.32 | 14.62 | 0.879 | 6.02 | 1379.45 | 61.76 | 4.48 |
| Propionic acid | 1.52 | 0.104 | 6.84 | 1.813 | 0.122 | 6.74 | 2.33 | 0.091 | 3.89 | 30.93 | 0.82 | 2.64 |
| Isobutyric acid | 0.23 | 0.010 | 4.45 | 0.265 | 0.008 | 3.12 | 0.30 | 0.017 | 5.63 | 3.09 | 0.07 | 2.40 |
| Butyric acid | 67.82 | 2.205 | 3.25 | 69.822 | 3.204 | 4.59 | 128.87 | 2.383 | 1.85 | 21.65 | 0.93 | 4.28 |
| 2-Methyl-butyric acid | 0.10 | 0.004 | 3.96 | 0.150 | 0.007 | 4.65 | 0.19 | 0.005 | 2.51 | 1.49 | 0.04 | 2.99 |
| Isovaleric acid | 0.13 | 0.009 | 6.83 | 0.151 | 0.010 | 6.41 | 0.16 | 0.013 | 8.24 | 1.94 | 0.06 | 3.22 |
| Valeric acid | 0.75 | 0.050 | 6.74 | 0.822 | 0.037 | 4.47 | 1.37 | 0.031 | 2.30 | 0.77 | 0.02 | 2.92 |
| Hexanoic acid | 32.30 | 1.506 | 4.66 | 35.342 | 1.922 | 5.44 | 64.07 | 1.690 | 2.64 | 1.90 | 0.08 | 4.17 |
| BHBA | 22.34 | 0.092 | 0.41 | 20.836 | 0.285 | 1.37 | 26.68 | 0.442 | 1.66 | 922.99 | 24.49 | 2.65 |
| Name | Raw Milk | Serum | ||
|---|---|---|---|---|
| Low | High | Low | High | |
| Acetic Acid | 74.3 ± 5.0 | 93.8 ± 0.7 | 79.7 ± 10.0 | 93.9 ± 4.0 |
| Propionic Acid | 97.4 ± 3.0 | 97.0 ± 0.6 | 112.9 ± 2.1 | 110.3 ± 3.1 |
| Isobutyric Acid | 100.6 ± 1.9 | 101.0 ± 1.1 | 110.3 ± 1.3 | 109.4 ± 1.5 |
| Butyric Acid | 98.1 ± 2.5 | 98.2 ± 0.5 | 110.5 ± 2.5 | 110.1 ± 1.8 |
| 2-Methylbutyric Acid | 96.9 ± 1.0 | 98.3 ± 1.5 | 113.4 ± 1.5 | 110.9 ± 0.4 |
| Isovaleric Acid | 99.5 ± 1.2 | 104.2 ± 0.6 | 111.8 ± 1.1 | 110.6 ± 1.0 |
| Valeric Acid | 98.4 ± 0.9 | 102.4 ± 0.6 | 111.1 ± 2.2 | 111.5 ± 2.8 |
| Hexanoic Acid | 96.6 ± 2.5 | 98.6 ± 0.7 | 102.4 ± 1.3 | 111.4 ± 0.8 |
| BHBA | 100.4 ± 0.1 | 96.9 ± 1.0 | 84.1 ± 8.4 | 96.4 ± 2.2 |
| Name | Raw Milk | Serum |
|---|---|---|
| Acetic Acid | 4.33–20.83 | 815.38–1922.32 |
| Propionic Acid | 0.68–2.57 | 14.86–43.76 |
| Isobutyric Acid | 0–1.02 | 1.59–18.63 |
| Butyric Acid | 11.58–175.46 | 11.81–31.00 |
| 2-Methylbutyric Acid | 0–0.39 | 0.88–11.95 |
| Isovaleric Acid | 0.20–0.39 | 1.27–3.43 |
| Valeric Acid | 0.39–1.96 | 0.10–0.78 |
| Hexanoic Acid | 4.82–58.06 | 0.09–1.72 |
| BHBA | 14.42–43.83 | 468.44–1372.64 |
| Raw Milk | Serum | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C2 | C3 | Iso-C4 | C4 | 2-MBA * | Iso-C5 | C5 | C6 | BHBA | C2 | C3 | Iso-C4 | C4 | 2-MBA | Iso-C5 | C5 | C6 | ||
| Raw Milk | C3 | 0.24 | ||||||||||||||||
| Iso-C4 | −0.09 | 0.26 | ||||||||||||||||
| C4 | −0.21 | 0.05 | 0.34 | |||||||||||||||
| 2-MBA | −0.22 | 0.25 | 0.92 | 0.31 | ||||||||||||||
| Iso-C5 | 0.10 | 0.67 | 0.26 | 0.10 | 0.34 | |||||||||||||
| C5 | −0.35 | 0.37 | 0.36 | 0.57 | 0.49 | 0.61 | ||||||||||||
| C6 | −0.17 | 0.10 | 0.28 | 0.98 | 0.28 | 0.14 | 0.55 | |||||||||||
| BHBA | 0.71 | −0.03 | 0.13 | −0.11 | −0.08 | −0.11 | −0.46 | −0.11 | ||||||||||
| Serum | C2 | 0.07 | −0.37 | 0.03 | −0.21 | −0.05 | −0.32 | −0.43 | −0.23 | 0.24 | ||||||||
| C3 | −0.21 | −0.18 | 0.13 | −0.28 | 0.23 | −0.01 | 0.02 | −0.30 | −0.25 | 0.59 | ||||||||
| Iso-C4 | −0.30 | −0.07 | 0.94 | 0.36 | 0.87 | 0.06 | 0.27 | 0.29 | 0.05 | 0.29 | 0.32 | |||||||
| C4 | −0.06 | −0.16 | 0.12 | −0.14 | 0.14 | −0.13 | −0.26 | −0.15 | 0.03 | 0.46 | 0.40 | 0.22 | ||||||
| 2-MBA | −0.37 | −0.01 | 0.91 | 0.35 | 0.95 | 0.17 | 0.41 | 0.29 | −0.13 | 0.20 | 0.38 | 0.94 | 0.27 | |||||
| Iso-C5 | −0.10 | −0.12 | 0.20 | −0.15 | 0.23 | −0.07 | −0.25 | −0.14 | 0.10 | 0.51 | 0.30 | 0.27 | 0.88 | 0.35 | ||||
| C5 | −0.10 | −0.34 | −0.23 | −0.24 | −0.04 | −0.30 | −0.14 | −0.22 | −0.18 | 0.25 | 0.54 | −0.04 | 0.35 | 0.08 | 0.27 | |||
| C6 | 0.18 | −0.24 | −0.27 | −0.23 | −0.31 | −0.25 | −0.43 | −0.20 | 0.23 | 0.49 | 0.48 | −0.05 | 0.00 | −0.19 | −0.12 | 0.30 | ||
| BHBA | 0.25 | −0.39 | −0.08 | −0.09 | −0.21 | −0.42 | −0.64 | −0.08 | 0.50 | 0.82 | 0.16 | 0.14 | 0.49 | 0.03 | 0.59 | 0.20 | 0.41 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Liu, Z.; Bath, C.; Marett, L.; Pryce, J.; Rochfort, S. Optimised Method for Short-Chain Fatty Acid Profiling of Bovine Milk and Serum. Molecules 2022, 27, 436. https://doi.org/10.3390/molecules27020436
Li C, Liu Z, Bath C, Marett L, Pryce J, Rochfort S. Optimised Method for Short-Chain Fatty Acid Profiling of Bovine Milk and Serum. Molecules. 2022; 27(2):436. https://doi.org/10.3390/molecules27020436
Chicago/Turabian StyleLi, Cheng, Zhiqian Liu, Carolyn Bath, Leah Marett, Jennie Pryce, and Simone Rochfort. 2022. "Optimised Method for Short-Chain Fatty Acid Profiling of Bovine Milk and Serum" Molecules 27, no. 2: 436. https://doi.org/10.3390/molecules27020436
APA StyleLi, C., Liu, Z., Bath, C., Marett, L., Pryce, J., & Rochfort, S. (2022). Optimised Method for Short-Chain Fatty Acid Profiling of Bovine Milk and Serum. Molecules, 27(2), 436. https://doi.org/10.3390/molecules27020436