


Effect of Culture Conditions on Fatty Acid Profiles of Bacteria and Lipopolysaccharides of the Genus Pseudomonas—GC-MS Analysis on Ionic Liquid-Based Column
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
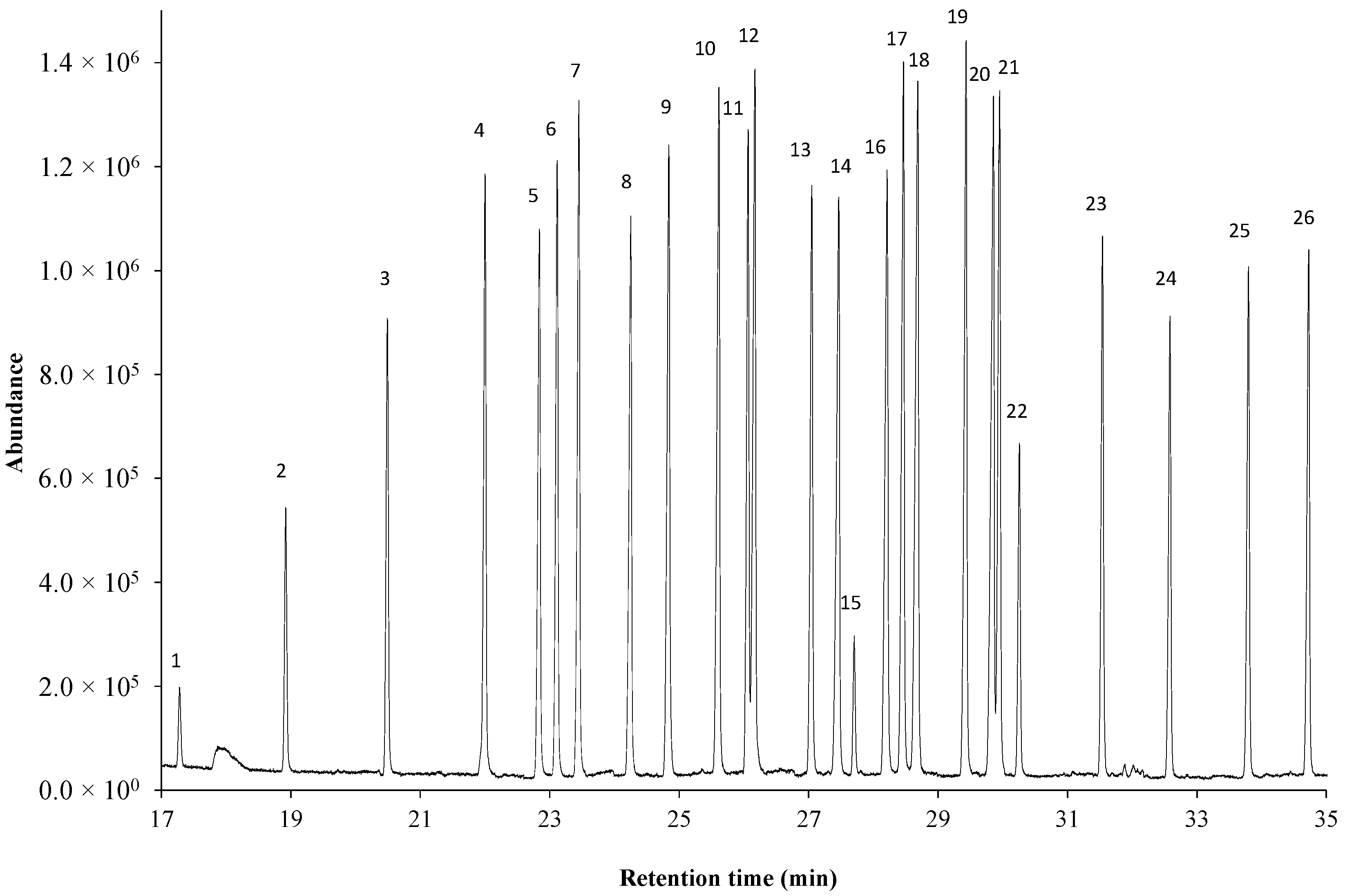
2.1. Separation on Ionic Liquid-Based Column
2.2. Evaluation of the Chromatographic Method
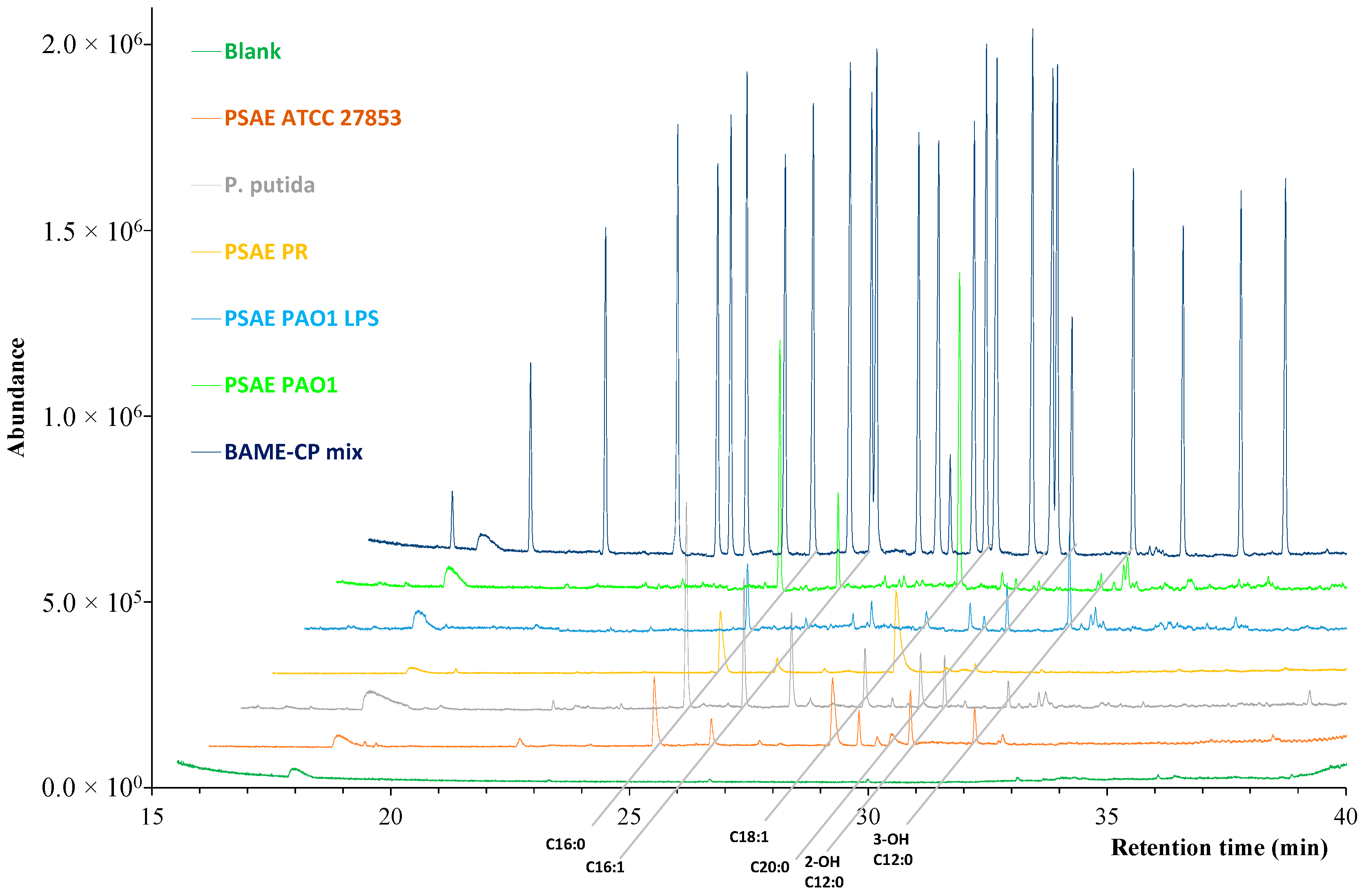
2.3. Fatty Acid Profiles of Bacteria
2.3.1. Representation of Optimal Conditions
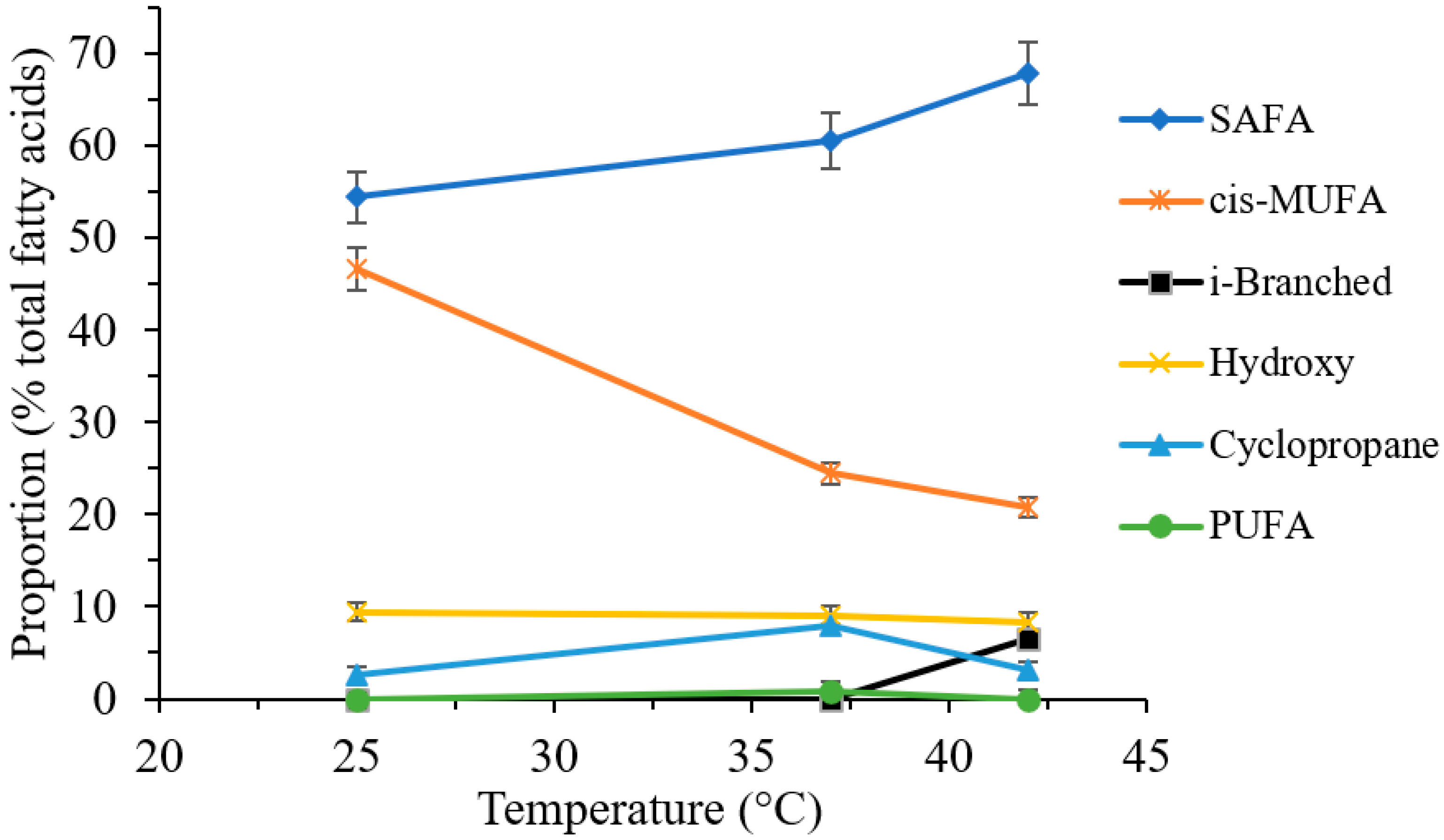
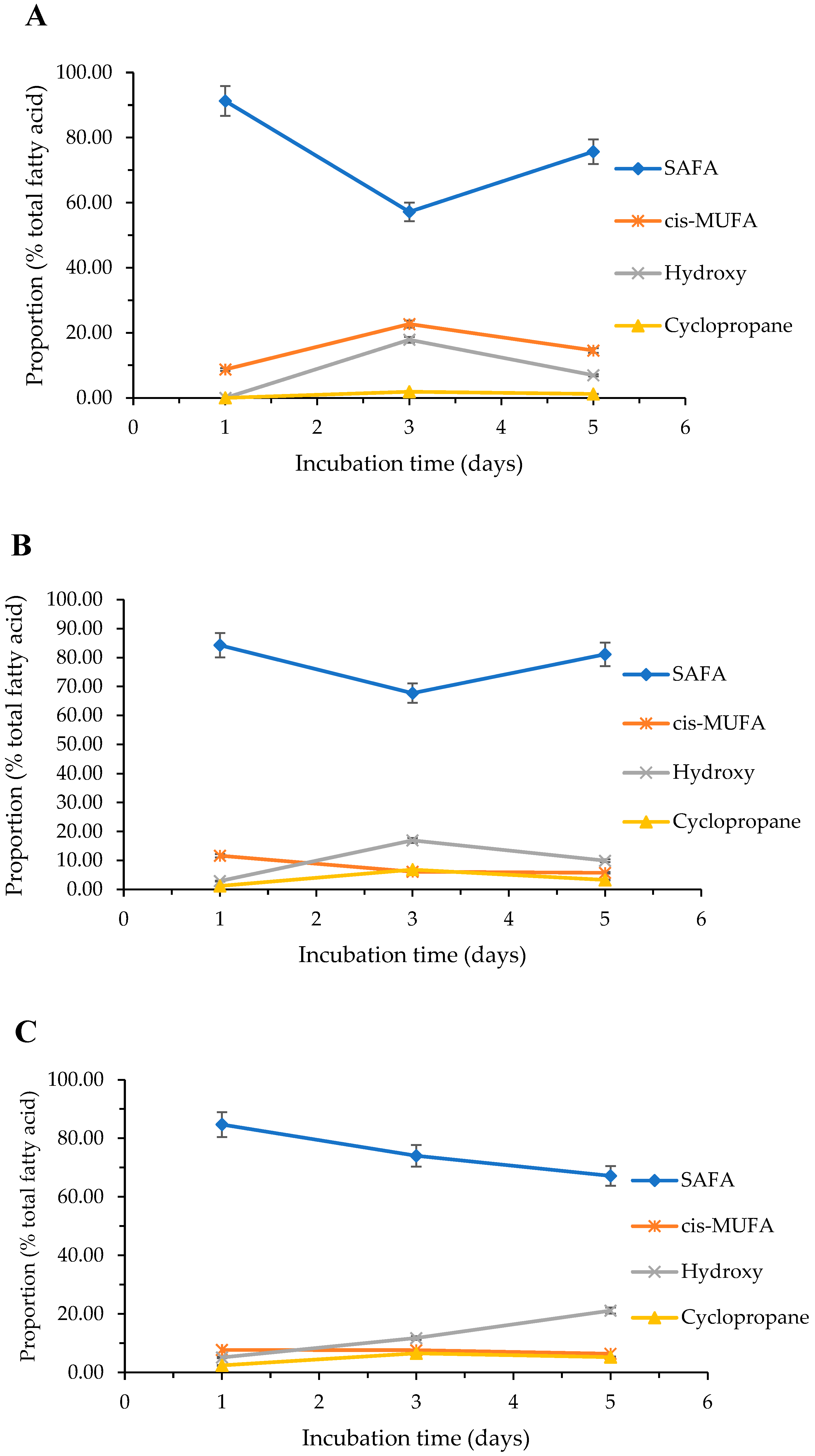
2.3.2. Effect of Cultivation Temperature on the Fatty Acid Composition
2.3.3. Effect of Cultivation Media on the Fatty Acid Composition
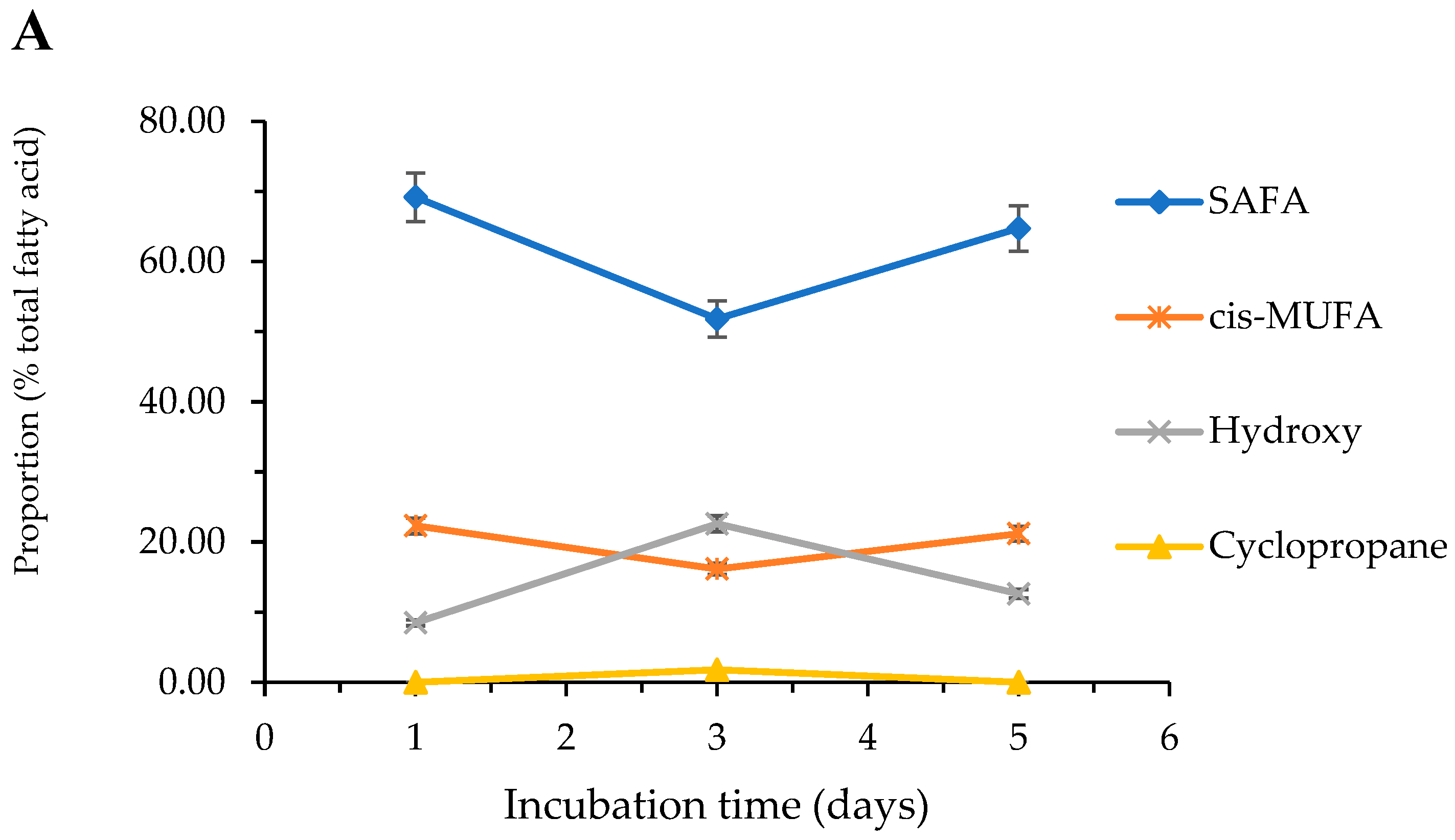
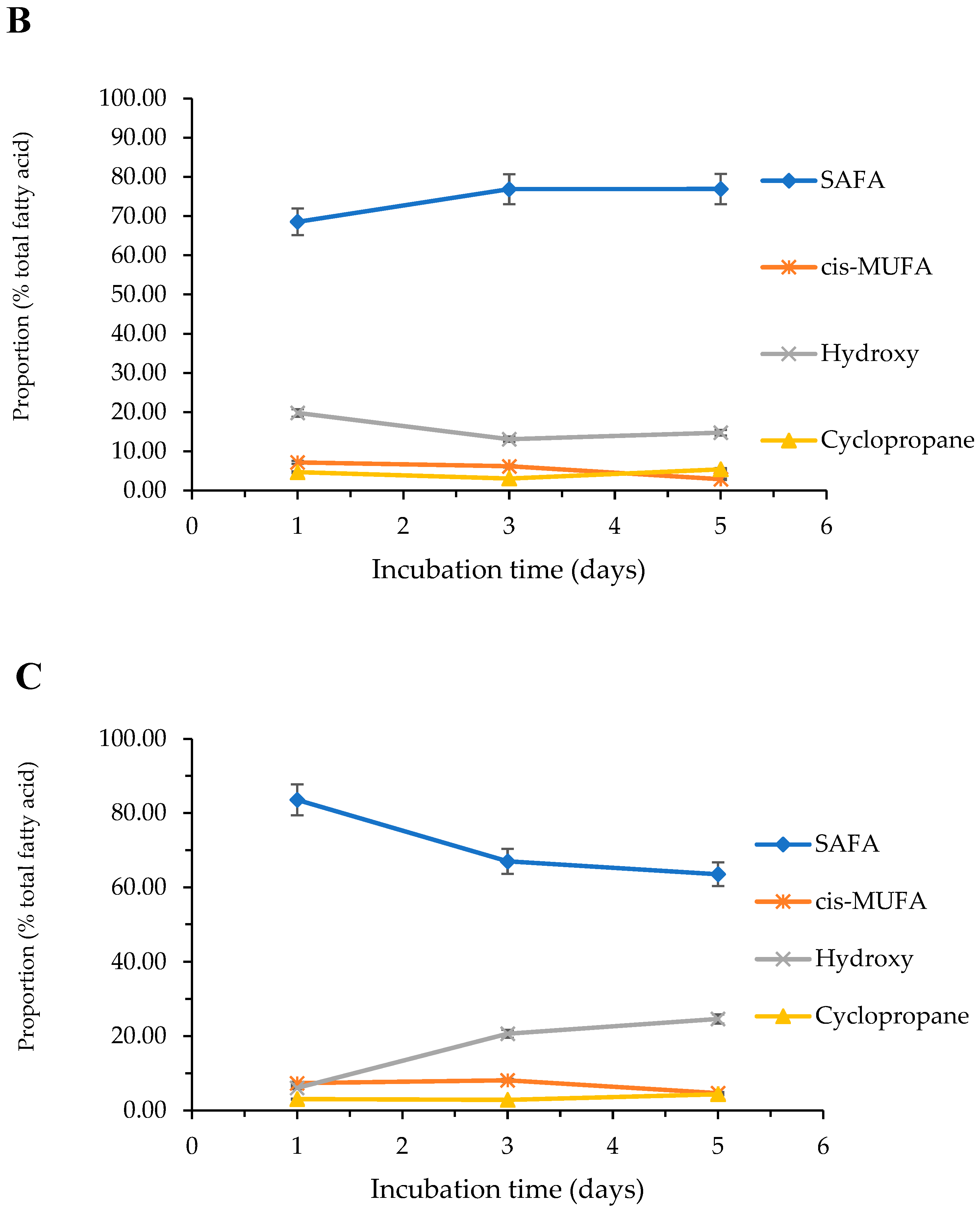
2.3.4. Effect of Cultivation Media and Time on the Fatty Acid Composition
2.3.5. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Samples and Derivative Preparation
4.2.1. Bacterial Strains
4.2.2. Preparation of Lipopolysaccharides
4.2.3. Derivatization Procedure
4.3. Gas Chromatography—Mass Spectrometry Analysis
4.4. Validation Procedure
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lukácová, M.; Barák, I.; Kazár, J. Role of structural variations of polysaccharide antigens in the pathogenicity of Gram-negative bacteria. Clin. Microbiol. Infect. 2008, 14, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Abel, K.; Deschmertzing, H.; Peterson, J.I. Classification of microorganisms by analysis of chemical composition I. Feasibility of utilizing gas chromatography. J. Bacteriol. 1963, 85, 1039–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himelbloom, B.H.; Oliveira, A.C.M.; Shetty, T.S. Rapid Methods for the Identification of Seafood Micro-Organisms; Wiley-Blackwell: Chichester, UK, 2010; pp. 226–236. [Google Scholar]
- Kropinski, A.M.; Lewis, V.; Berry, D. Effect of growth temperature on the lipids, outer membrane proteins, and lipopolysaccharides of Pseudomonas aeruginosa PAO. J. Bacteriol. 1987, 169, 1960–1966. [Google Scholar] [CrossRef] [Green Version]
- Russell, N.J.; Evans, R.I.; ter Steeg, P.F.; Hellemons, J.; Verheul, A.; Abee, T. Membranes as a target for stress adaptation. Int. J. Food Microbiol. 1995, 28, 255–261. [Google Scholar] [CrossRef]
- Mrozik, A.; Labuzek, S.; Piotrowska-Seget, Z. Changes in fatty acid composition in Pseudomonas putida and Pseudomonas stutzeri during naphthalene degradation. Microbiol. Res. 2005, 160, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Mrozik, A.; Piotrowska-Seget, Z.; Łabuzek, S. Changes in whole cell-derived fatty acids induced by naphthalene in bacteria from genus Pseudomonas. Microbiol. Res. 2004, 159, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Cronan, J.E.; Luk, T. Advances in the Structural Biology, Mechanism, and Physiology of Cyclopropane Fatty Acid Modifications of Bacterial Membranes. Microbiol. Mol. Biol. Rev. 2022, 86, e0001322. [Google Scholar] [CrossRef]
- Chapter 32-Nutritional Strategies in Gastrointestinal Disease. In Canine and Feline Gastroenterology; Washabau, R.J.; Day, M.J. (Eds.) W.B. Saunders: Saint Louis, MI, USA, 2013; pp. 409–428. [Google Scholar]
- Segura, A.; Duque, E.; Mosqueda, G.; Ramos, J.L.; Junker, F. Multiple responses of Gram-negative bacteria to organic solvents. Env. Microbiol. 1999, 1, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberlein, C.; Baumgarten, T.; Starke, S.; Heipieper, H.J. Immediate response mechanisms of Gram-negative solvent-tolerant bacteria to cope with environmental stress: Cistrans isomerization of unsaturated fatty acids and outer membrane vesicle secretion. Appl. Microbiol. Biotechnol. 2018, 102, 2583–2593. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Wolfaardt, G.M.; Arts, M.T. Characterization of Pseudomonas aeruginosa fatty acid profiles in biofilms and batch planktonic cultures. Can. J. Microbiol. 2010, 56, 1028–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mező, E.; Bufa, A.; Páger, C.; Poór, V.; Marosvölgyi, T.; Kilár, F.; Makszin, L. The Role of Ionic Liquid Interaction in the Separation of Fatty Acid Methyl Esters—Polyunsaturated Geometric Isomers in GC–MS. Separations 2021, 8, 38. [Google Scholar] [CrossRef]
- LaBauve, A.E.; Wargo, M.J. Growth and laboratory maintenance of Pseudomonas aeruginosa. Curr. Protoc. Microbiol. 2012, 25. Chapter 6, Unit 6E.1. Available online: https://currentprotocols.onlinelibrary.wiley.co (accessed on 3 September 2022). [CrossRef] [PubMed] [Green Version]
- Lingfa, L. Study of membrane fatty acids of Gram-negative bacteria and its influence towards the terrestrial ecosystem. Res. J. Pharm. Biol. Chem. Sci. 2015, 4, 1–19. [Google Scholar]
- Kim, B.H.; Kim, S.; Kim, H.G.; Lee, J.; Lee, I.S.; Park, Y.K. The formation of cyclopropane fatty acids in Salmonella enterica serovar Typhimurium. Microbiology 2005, 151, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodowska, J.; Wolny, D.; Weglarz, L.; Dzierzewicz, Z. The structural diversity of lipid A from Gram-negative bacteria. Postepy Hig Med. Dosw 2007, 61, 106–121. [Google Scholar]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Junker, F.; Ramos, J.L. Involvement of the cis/trans isomerase Cti in solvent resistance of Pseudomonas putida DOT-T1E. J. Bacteriol. 1999, 181, 5693–5700. [Google Scholar] [CrossRef] [Green Version]
- Wayne Moss, C.; Dees, S.B. Identification of microorganisms by gas chromatographic-mass spectrometric analysis of cellular fatty acids. J. Chromatogr. A 1975, 112, 595–604. [Google Scholar] [CrossRef]
- Buchanan, P.J.; Ernst, R.K.; Elborn, J.S.; Schock, B. Role of CFTR, Pseudomonas aeruginosa and Toll-like receptors in cystic fibrosis lung inflammation. Biochem. Soc. Trans. 2009, 37, 863–867. [Google Scholar] [CrossRef] [Green Version]
- Walczak-Skierska, J.; Złoch, M.; Pauter, K.; Pomastowski, P.; Buszewski, B. Lipidomic analysis of lactic acid bacteria strains by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. J. Dairy Sci. 2020, 103, 11062–11078. [Google Scholar] [CrossRef] [PubMed]
- Scherer, C.; Müller, K.D.; Rath, P.M.; Ansorg, R.A. Influence of culture conditions on the fatty acid profiles of laboratory-adapted and freshly isolated strains of Helicobacter pylori. J. Clin. Microbiol. 2003, 41, 1114–1117. [Google Scholar] [CrossRef] [PubMed]
- Paulton, R.J.L. The bacterial growth curve. J. Biol. Educ. 1991, 25, 92–94. [Google Scholar] [CrossRef]
- Khelissa, S.O.; Abdallah, M.; Jama, C.; Chihib, N.E. Actively detached Pseudomonas aeruginosa biofilm cell susceptibility to benzalkonium chloride and associated resistance mechanism. Arch. Microbiol. 2019, 201, 747–755. [Google Scholar] [CrossRef]
- Luo, J.; Kong, J.L.; Dong, B.Y.; Huang, H.; Wang, K.; Wu, L.H.; Hou, C.C.; Liang, Y.; Li, B.; Chen, Y.Q. Baicalein attenuates the quorum sensing-controlled virulence factors of Pseudomonas aeruginosa and relieves the inflammatory response in P. aeruginosa-infected macrophages by downregulating the MAPK and NFκB signal-transduction pathways. Drug Des. Dev. 2016, 10, 183–203. [Google Scholar] [CrossRef] [Green Version]
- Al-kafaween, M.A.; Mohd Hilmi, A.B.; Jaffar, N.; Al-Jamal, H.A.N.; Zahri, M.K. Determination of optimum incubation time for formation of Pseudomonas aeruginosa and Streptococcus pyogenes biofilms in microtiter plate. Bull. Natl. Res. Cent. 2019, 43, 100. [Google Scholar] [CrossRef] [Green Version]
- Madison, L.L.; Huisman, G.W. Metabolic engineering of poly(3-hydroxyalkanoates): From DNA to plastic. Microbiol. Mol. Biol. Rev. 1999, 63, 21–53. [Google Scholar] [CrossRef] [Green Version]
- Ayub, N.D.; Julia Pettinari, M.; Méndez, B.S.; López, N.I. Impaired polyhydroxybutyrate biosynthesis from glucose in Pseudomonas sp. 14-3 is due to a defective beta-ketothiolase gene. FEMS Microbiol. Lett. 2006, 264, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Signature Lipid Biomarker (SLB). Analysis in Determining Changes in Community Structure of Soil Microorganisms. Pol. J. Environ. Stud. 2003, 12, 669–675. [Google Scholar]
- de Santana-Filho, A.P.; Noleto, G.R.; Gorin, P.A.J.; de Souza, L.M.; Iacomini, M.; Sassaki, G.L. GC–MS detection and quantification of lipopolysaccharides in polysaccharides through 3-O-acetyl fatty acid methyl esters. Carbohydr. Polym. 2012, 87, 2730–2734. [Google Scholar] [CrossRef] [Green Version]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides. Extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- As, S.; Ramasamy, P. Fatty Acid Methyl Ester (FAME) Analysis of Moderately Thermophilic Bacteria Isolated from the Coramandal Coast, Chennai, Tamilnadu. Eur. J. Exp. Biol. 2016, 6, 1–7. [Google Scholar]
- Taverniers, I.; De Loose, M.; Van Bockstaele, E. Trends in quality in the analytical laboratory. II. Analytical method validation and quality assurance. TrAC Trends Anal. Chem. 2004, 23, 535–552. [Google Scholar] [CrossRef]
- FDA Laboratory Manual Volume II: Methods, Method Verification and Validation. Available online: https://www.fda.gov/media/73920/download (accessed on 3 September 2022).
- Karnes, H.T.; March, C. Precision, Accuracy, and Data Acceptance Criteria in Biopharmaceutical Analysis. Pharm. Res. 1993, 10, 1420–1426. [Google Scholar] [CrossRef]
- Dodds, E.D.; McCoy, M.R.; Rea, L.D.; Kennish, J.M. Gas chromatographic quantification of fatty acid methyl esters: Flame ionization detection vs. electron impact mass spectrometry. Lipids 2005, 40, 419–428. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Label | Compound ID | Name | Retention Time (Min) * |
|---|---|---|---|
| 1 | C11:0 | Methyl undecanoate | 17.28 |
| 2 | C12:0 | Methyl dodecanoate | 18.92 |
| 3 | C13:0 | Methyl tridecanoate | 20.49 |
| 4 | C14:0 | Methyl tetradecanoate | 22.01 |
| 5 | i C15:0 | Methyl 13-methyltetradecanoate | 22.84 |
| 6 | ai C15:0 | Methyl 12-methyltetradecanoate | 23.12 |
| 7 | C15:0 | Methyl pentadecanoate | 23.45 |
| 8 | i C16:0 | Methyl 14-methylpentadecanoate | 24.26 |
| 9 | C16:0 | Methyl hexadecanoate | 24.85 |
| 10 | i C17:0 | Methyl 15-methylhexadecanoate | 25.62 |
| 11 | C16:1c | Methyl hexadecenoate (cis-9) | 26.07 |
| 12 | C17:0 | Methyl heptadecanoate | 26.18 |
| 13 | C17:0Δ | Methyl cis-9,10-methylenehexadecanoate | 27.05 |
| 14 | C18:0 | Methyl octadecanoate | 27.48 |
| 15 | 2-OH C10:0 | Methyl 2-hydroxydecanoate | 27.70 |
| 16 | C18:1t | Methyl octadecenoate (trans-9) | 28.22 |
| 17 | C18:1c | Methyl octadecenoate (cis-9) | 28.47 |
| 18 | C19:0 | Methyl nonadecanoate | 28.70 |
| 19 | C19:0Δ | Methyl cis-9,10-methyleneoctadecanoate | 29.44 |
| 20 | C20:0 | Methyl eicosanoate | 29.86 |
| 21 | C18:2cc | Methyl octadecadienote (all cis 9,12) | 29.96 |
| 22 | 2-OH C12:0 | Methyl 2-hydroxydodecanoate | 30.26 |
| 23 | 3-OH C12:0 | Methyl 3-hydroxydodecanoate | 31.55 |
| 24 | 2-OH C14:0 | Methyl 2-hydroxytetradecanoate | 32.59 |
| 25 | 3-OH C14:0 | Methyl 3-hydroxytetradecanoate | 33.80 |
| 26 | 2-OH C16:0 | Methyl 2-hydroxyhexadecanoate | 34.73 |
| Fatty Acids | PSAE PAO1 | PSAE ATCC 27853 | P. putida | PSAE PR | Mean |
|---|---|---|---|---|---|
| % of total fatty acids | |||||
| Ʃ SAFA | 36.1 | 68.6 | 46.1 | 91.5 | 60.6 |
| Ʃ cis MUFA | 55.9 | 7.2 | 28.2 | 6.4 | 24.4 |
| Ʃ Hydroxy FA | 4.3 | 19.8 | 10.6 | 2.1 | 9.0 |
| Ʃ Cyclopropane FA | 3.8 | 4.7 | 15.6 | - | 8.0 |
| Ʃ PUFA | - | 0.8 | - | - | 0.8 |
| Ʃ MCFA | 4.3 | 20.9 | 10.1 | 2.1 | 9.3 |
| Ʃ LCFA | 95.8 | 77.2 | 90.0 | 97.9 | 90.2 |
| Ʃ VLCFA | - | 3.0 | - | - | 0.7 |
| Fatty Acids | PSAE PAO1 | PSAE PAO1 LPS | P. putida | PSAE PR | Retention Time (Min) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | ||
| Saturated fatty acids (SAFAs) | |||||||||||||
| C11:0 | - | - | - | 33.6 | - | 46.0 | - | - | - | - | - | - | 17.28 |
| C14:0 | - | - | - | - | - | - | - | 1.3 | - | - | - | - | 22.01 |
| C15:0 | - | - | - | - | - | - | - | 0.7 | - | - | - | - | 23.45 |
| C16:0 | 18.8 | 32.6 | 39.5 | 10.1 | 20.6 | 5.3 | 17.9 | 33.1 | 11.6 | 12.6 | 30.9 | 37.3 | 24.85 |
| C17:0 | - | - | - | - | - | - | - | - | 3.8 | 10.1 | - | - | 26.18 |
| C18:0 | - | 1.9 | 1.7 | 7.6 | 8.3 | 7.3 | 1.1 | 1.9 | 16.1 | 42.1 | - | - | 27.48 |
| C19:0 | - | - | - | - | - | - | - | - | - | 19.0 | 57.0 | 46.0 | 28.70 |
| C20:0 | - | 1.6 | - | - | 5.5 | 3.8 | 18.3 | 9.1 | 31.5 | 8.4 | 3.5 | - | 29.86 |
| cis Monounsaturated fatty acids (MUFAs) | |||||||||||||
| C16:1c | 26.7 | 11.1 | 8.3 | 4.9 | 2.8 | 3.7 | 35.1 | 18.0 | - | - | 6.4 | 10.6 | 26.07 |
| C18:1c | 47.4 | 44.7 | 39.9 | 7.4 | 6.3 | 7.5 | 8.3 | 10.2 | 16.6 | - | - | - | 28.47 |
| Branched fatty acids | |||||||||||||
| iso | |||||||||||||
| i C15:0 | - | - | - | - | - | - | - | - | 3.2 | - | - | - | 22.84 |
| i C16:0 | - | - | - | - | - | - | - | - | 2.6 | - | - | - | 24.26 |
| i C17:0 | - | - | - | - | - | - | - | - | 0.7 | - | - | - | 25.62 |
| Hydroxy fatty acids | |||||||||||||
| 2-OH C10:0 | - | 1.2 | 0.8 | - | - | - | - | - | - | - | - | - | 27.70 |
| 2-OH C12:0 | 1.0 | 1.0 | 2.9 | 16.3 | 13.5 | 2.5 | 11.4 | 6.4 | - | - | 2.1 | 4.3 | 30.26 |
| 3-OH C12:0 | 3.2 | 2.0 | 3.9 | 20.0 | 29.6 | 4.1 | 5.7 | 3.7 | 4.6 | - | - | 1.8 | 31.55 |
| 2-OH C14:0 | - | - | - | - | - | - | - | - | 4.7 | 7.8 | - | - | 32.59 |
| 3-OH C14:0 | - | - | - | - | - | - | - | - | 4.3 | - | - | - | 33.80 |
| Cyclopropane fatty acids | |||||||||||||
| C17:0Δ | - | 1.3 | 1.3 | - | 4.7 | - | 2.3 | 14.9 | - | - | - | - | 27.05 |
| C19:0Δ | 2.8 | 2.5 | 1.7 | - | 8.7 | - | - | 0.6 | - | - | - | - | 29.44 |
| Polyunsaturated fatty acids (PUFAs) | |||||||||||||
| C20:2cc | - | - | - | - | - | 19.8 | - | - | - | - | - | - | 32.06 |
| Ʃ SAFA | 18.8 | 36.1 | 41.1 | 51.3 | 34.3 | 62.4 | 37.3 | 46.1 | 63.1 | 92.2 | 91.5 | 83.3 | |
| Ʃ cis MUFA | 74.1 | 55.9 | 48.3 | 12.3 | 9.1 | 11.2 | 43.3 | 28.2 | 16.6 | - | 6.4 | 10.6 | |
| Ʃ Hydroxy FA | 4.2 | 4.2 | 7.5 | 36.4 | 43.1 | 6.6 | 17.1 | 10.1 | 13.7 | 7.8 | 2.1 | 6.0 | |
| Ʃ Cyclopropane FA | 2.8 | 3.8 | 3.0 | - | 13.4 | - | 2.3 | 15.6 | - | - | - | - | |
| Ʃ PUFA | - | - | - | - | - | 19.8 | - | - | - | - | - | - | |
| Ʃ MCFA | 4.2 | 4.2 | 7.5 | 70.0 | 43.1 | 52.6 | 17.1 | 10.1 | 4.6 | - | 2.1 | 6.0 | |
| Ʃ LCFA | 95.8 | 95.8 | 92.5 | 30.0 | 56.9 | 47.4 | 82.9 | 89.9 | 95.4 | 100.0 | 97.9 | 94.0 | |
| Fatty Acids | PSAE ATCC 27853 | ||
|---|---|---|---|
| Blood Agar | |||
| 25 °C | 37 °C | 42 °C | |
| Saturated fatty acids (SAFAs) | |||
| C16:0 | 12.7 | 27.7 | 26.8 |
| C19:0 | 74.0 | 36.3 | 36.1 |
| C20:0 | 4.6 | 20.2 | 21.8 |
| cis Monounsaturated fatty acids (MUFAs) | |||
| C16:1c | 6.0 | 11.6 | 7.7 |
| C18:1c | 2.8 | - | - |
| Hydroxy fatty acids | |||
| 2-OH C10:0 | - | - | - |
| 2-OH C12:0 | - | 1.9 | 3.2 |
| 3-OH C12:0 | - | 1.0 | 1.9 |
| 2-OH C14:0 | - | - | - |
| 3-OH C14:0 | - | - | - |
| 3-OH C16:0 | - | - | - |
| Cyclopropane fatty acids | |||
| C17:0Δ | - | - | 1.0 |
| C19:0Δ | - | 1.2 | 1.5 |
| Ʃ SAFA | 91.3 | 84.3 | 84.7 |
| Ʃ MUFA | |||
| Ʃ cis | 8.7 | 11.6 | 7.7 |
| Ʃ Hydroxy FA | - | 2.9 | 5.1 |
| Ʃ Cyclic FA | - | 1.2 | 2.4 |
| Ʃ MCFA | - | 2.9 | 5.1 |
| Ʃ LCFA | 100.0 | 97.1 | 94.9 |
| Fatty Acids | 1 Day | 3 Days | 5 Days | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Agar | Blood Agar | Agar | Blood Agar | Agar | Blood Agar | |||||||||||||
| 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | 25 °C | 37 °C | 42 °C | |
| (n = 3) | ||||||||||||||||||
| Saturated fatty acids (SAFAs) | ||||||||||||||||||
| C12:0 | - | 1.1 | - | - | - | - | 2.8 | - | - | 4.7 | ||||||||
| C14:0 | - | 3.6 | - | - | - | - | 1.3 | 1.1 | - | 1.8 | - | - | 1.2 | 3.2 | - | - | - | 3.1 |
| C15:0 | - | 0.8 | - | - | - | - | 0.4 | 1.1 | - | 1.0 | - | - | - | - | - | - | - | - |
| C16:0 | 14.1 | 23.2 | 27.0 | 12.7 | 27.7 | 26.8 | 14.6 | 23.2 | 27.0 | 20.3 | 27.5 | 27.6 | 21.4 | 26.2 | 25.2 | 18.3 | 19.0 | 24.8 |
| C17:0 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1.7 | - | - | - | - |
| C18:0 | - | 1.2 | - | - | - | - | - | 1.8 | 1.0 | - | 1.4 | - | - | 1.5 | - | - | - | 3.4 |
| C19:0 | 39.7 | 28.3 | 36.7 | 74.0 | 36.3 | 36.1 | 25.8 | 30.9 | 26.0 | 24.8 | 28.1 | 29.8 | 42.2 | 26.6 | 24.1 | 54.6 | 27.5 | 22.5 |
| C20:0 | 14.1 | 7.4 | 19.9 | 4.6 | 20.2 | 21.8 | 5.5 | 18.7 | 13.0 | 3.3 | 10.7 | 16.7 | - | 17.8 | 12.3 | 2.7 | 34.6 | 13.4 |
| C22:0 | 1.3 | 3.0 | - | - | - | - | 1.5 | - | - | 1.3 | - | - | - | - | 2.7 | - | - | - |
| Monounsaturated fatty acids (MUFAs) | ||||||||||||||||||
| cis | ||||||||||||||||||
| C16:1c | 22.3 | 7.2 | 7.3 | 6.0 | 11.6 | 7.7 | 16.2 | 6.2 | 8.1 | 21.6 | 5.3 | 6.7 | 21.2 | 2.9 | 4.7 | 14.6 | 5.7 | 6.4 |
| C18:1c | - | - | - | 2.8 | - | - | - | - | - | 1.1 | 0.8 | 1.0 | - | - | - | - | - | - |
| trans | ||||||||||||||||||
| C16:1t | - | - | - | - | - | - | 5.1 | - | - | 0.4 | 2.4 | - | - | - | - | 1.7 | - | - |
| Branched fatty acids | ||||||||||||||||||
| iso | ||||||||||||||||||
| i C17:0 | - | - | - | - | - | - | 2.5 | - | - | - | - | - | 2.6 | - | - | - | - | - |
| Hydroxy fatty acids | ||||||||||||||||||
| 2-OH C10:0 | - | - | - | - | - | - | 9.7 | - | - | - | 3.7 | - | 1.3 | 2.3 | 6.3 | - | - | - |
| 2-OH C12:0 | 6.3 | 11.6 | 4.4 | - | 1.9 | 3.2 | 9.3 | 6.3 | 12.1 | 13.8 | 8.8 | 9.1 | 6.3 | 7.9 | 3.4 | 5.2 | 13.7 | |
| 3-OH C12:0 | 2.2 | 8.2 | 1.7 | - | 1.0 | 1.9 | 3.6 | 6.8 | 8.5 | 4.1 | 4.4 | 2.7 | 11.4 | 6.2 | 10.8 | 3.6 | 4.7 | 7.4 |
| Cyclopropane fatty acids | ||||||||||||||||||
| C17:0Δ | - | 1.6 | 1.2 | - | - | 1.0 | 0.9 | 0.8 | 1.4 | 1.1 | 2.4 | 3.3 | - | 2.0 | 1.4 | - | - | 2.1 |
| C19:0Δ | - | 3.1 | 1.8 | - | 1.2 | 1.5 | 0.9 | 2.2 | 1.4 | 0.8 | 4.5 | 3.2 | - | 3.4 | 3.0 | 1.2 | 3.2 | 3.1 |
| Polyunsaturated fatty acids (PUFAs) | ||||||||||||||||||
| C20:2cc | - | 0.8 | - | - | - | - | - | 0.8 | 1.4 | - | - | - | - | - | 1.7 | - | - | - |
| Ʃ SAFA | 69.2 | 68.6 | 83.6 | 91.3 | 84.3 | 84.7 | 51.8 | 76.9 | 67.0 | 57.2 | 67.7 | 74.1 | 64.7 | 76.9 | 64.3 | 75.7 | 81.1 | 67.2 |
| Ʃ cis | 22.3 | 7.2 | 7.3 | 8.7 | 11.6 | 7.7 | 16.2 | 6.2 | 8.1 | 22.7 | 6.1 | 7.6 | 21.2 | 2.9 | 4.7 | 14.6 | 5.7 | 6.4 |
| Ʃ Branched FA | ||||||||||||||||||
| Ʃ iso | - | - | - | - | - | - | 2.5 | - | - | - | - | - | 2.6 | - | - | - | - | - |
| Ʃ Hydroxy FA | 8.5 | 19.8 | 6.1 | - | 2.9 | 5.1 | 22.6 | 13.1 | 20.6 | 17.9 | 17.0 | 11.8 | 12.6 | 14.8 | 24.9 | 6.9 | 9.9 | 21.2 |
| Ʃ Cyclopropane FA | - | 4.7 | 3.0 | - | 1.2 | 2.4 | 1.8 | 3.1 | 2.9 | 1.9 | 6.8 | 6.5 | - | 5.4 | 4.4 | 1.2 | 3.2 | 5.2 |
| Ʃ PUFA | - | 0.8 | - | - | - | - | - | 0.8 | 1.4 | - | - | - | - | - | 1.7 | - | - | - |
| Ʃ MCFA | 8.5 | 20.9 | 6.1 | - | 2.9 | 5.1 | 25.4 | 13.1 | 20.6 | 22.6 | 17.0 | 11.8 | 12.6 | 14.8 | 24.6 | 6.9 | 9.9 | 21.2 |
| Ʃ LCFA | 90.2 | 77.2 | 93.9 | 100.0 | 97.1 | 94.9 | 73.1 | 86.9 | 79.4 | 76.1 | 83.0 | 88.2 | 88.5 | 85.2 | 72.4 | 93.1 | 90.1 | 78.8 |
| Ʃ VLCFA | 1.3 | 3.0 | - | - | - | - | 1.5 | - | - | 1.3 | - | - | - | - | 2.7 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mező, E.; Hartmann-Balogh, F.; Madarászné Horváth, I.; Bufa, A.; Marosvölgyi, T.; Kocsis, B.; Makszin, L. Effect of Culture Conditions on Fatty Acid Profiles of Bacteria and Lipopolysaccharides of the Genus Pseudomonas—GC-MS Analysis on Ionic Liquid-Based Column. Molecules 2022, 27, 6930. https://doi.org/10.3390/molecules27206930
Mező E, Hartmann-Balogh F, Madarászné Horváth I, Bufa A, Marosvölgyi T, Kocsis B, Makszin L. Effect of Culture Conditions on Fatty Acid Profiles of Bacteria and Lipopolysaccharides of the Genus Pseudomonas—GC-MS Analysis on Ionic Liquid-Based Column. Molecules. 2022; 27(20):6930. https://doi.org/10.3390/molecules27206930
Chicago/Turabian StyleMező, Emerencia, Fruzsina Hartmann-Balogh, Ibolya Madarászné Horváth, Anita Bufa, Tamás Marosvölgyi, Béla Kocsis, and Lilla Makszin. 2022. "Effect of Culture Conditions on Fatty Acid Profiles of Bacteria and Lipopolysaccharides of the Genus Pseudomonas—GC-MS Analysis on Ionic Liquid-Based Column" Molecules 27, no. 20: 6930. https://doi.org/10.3390/molecules27206930
APA StyleMező, E., Hartmann-Balogh, F., Madarászné Horváth, I., Bufa, A., Marosvölgyi, T., Kocsis, B., & Makszin, L. (2022). Effect of Culture Conditions on Fatty Acid Profiles of Bacteria and Lipopolysaccharides of the Genus Pseudomonas—GC-MS Analysis on Ionic Liquid-Based Column. Molecules, 27(20), 6930. https://doi.org/10.3390/molecules27206930