


Copper(II) Complex Containing 4-Fluorophenoxyacetic Acid Hydrazide and 1,10-Phenanthroline: A Prostate Cancer Cell-Selective and Low-Toxic Copper(II) Compound
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Complex 1 Downregulates Proliferation Markers on PCa Cells
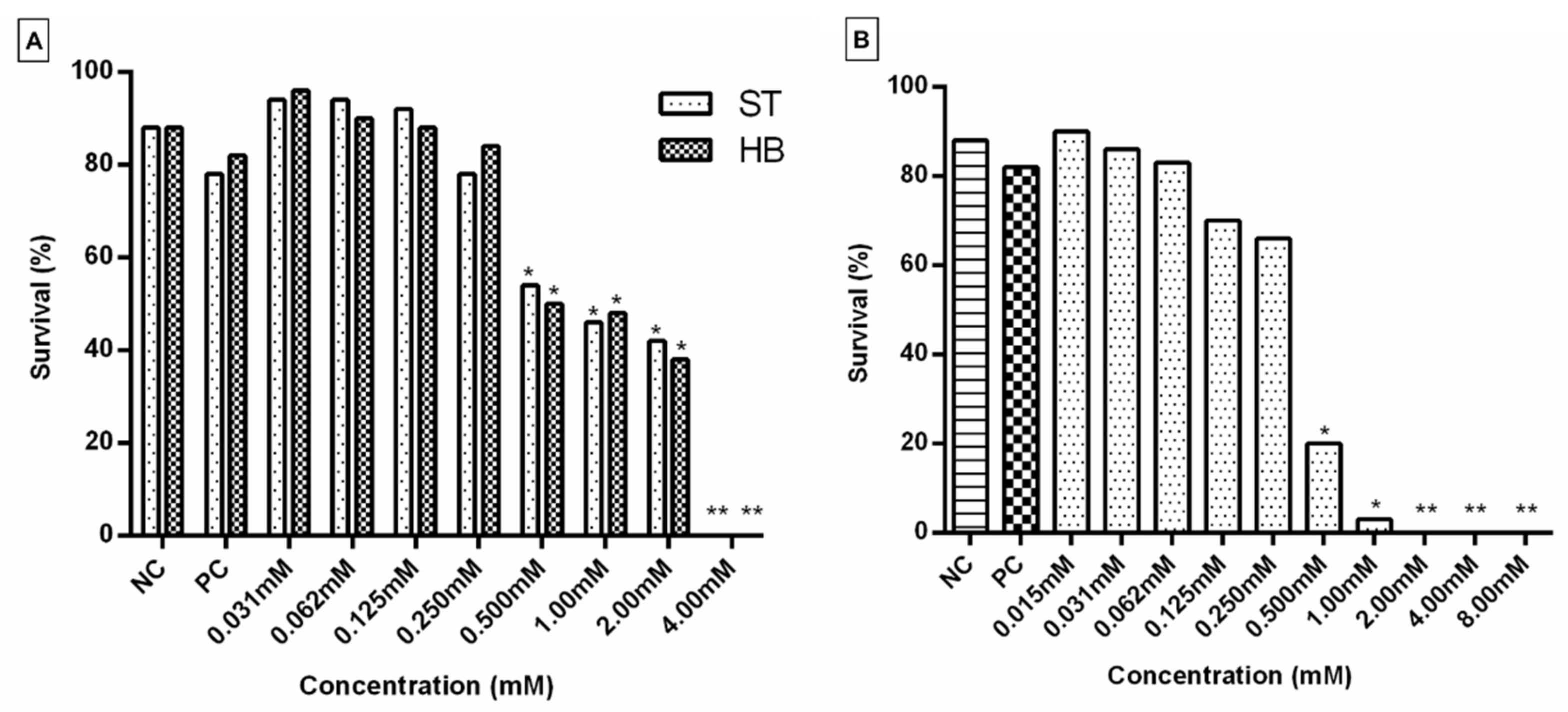
2.2. Toxicity Test (TX)
2.3. Somatic Mutation and Recombination Test (SMART)
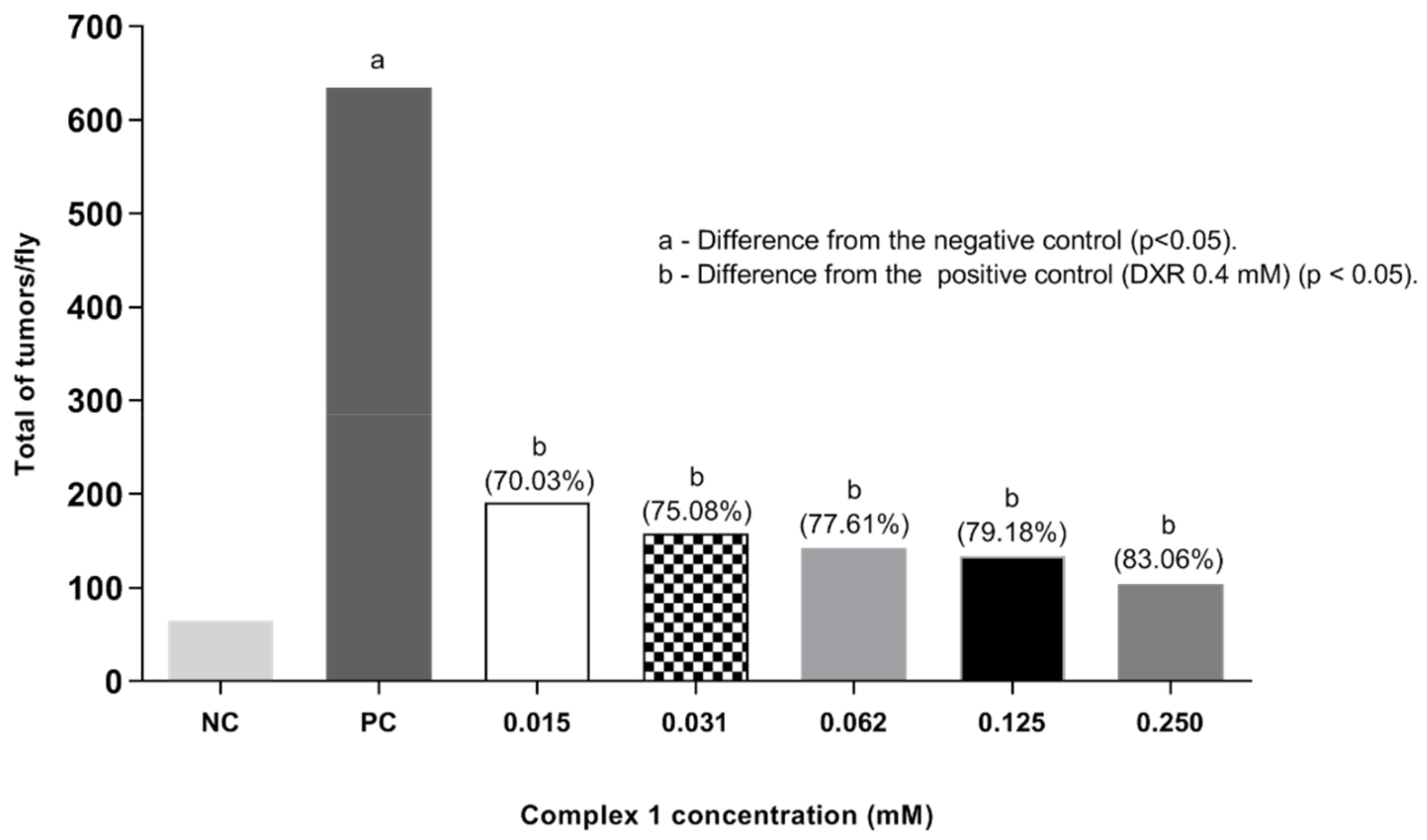
2.4. Epithelial Tumor Test (ETT)
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complexes | Cell Lines | IC50 (24 h) | Reference |
|---|---|---|---|
| [Cu(dox)(phen)(H2O)(ClO4)](ClO4) | K562 | 1.93 µM | [27] |
| [Cu(tc)(phen)(H2O)(ClO4)](ClO4) | K562 | 2.59 µM | [27] |
| [Cu(OH-PIP)(Phe)Cl] | CAL-51 | 0.52 µM | [28] |
| [Cu(OH-PIP)(Phe)Cl] | MDA-MB-231 | 18.89 µM | [28] |
| [Cu(OH-PIP)(Phe)Cl] | MCF-7 | 30.88 µM | [28] |
| [Cu(dox)(phen)]2+ | B16F10 | 1.4 µM | [29] |
| [Cu(dox)(phen)]2+ | Sarcoma TG180 | 6.2 μM | [29] |
| [Cu(dox)(phen)]2+ | Sarcoma S180 | 13.3 μM | [29] |
| bis[(µ2-chloro)chloro(1,10-phenanthroline)copper(II)] | MDA-MB-32 | 0.65 μM | [30] |
| bis[(µ2-chloro)chloro(1,10-phenanthroline)copper(II)] | HT-29 | 0.60 μM | [30] |
| bis[(µ2-chloro)chloro(1,10-phenanthroline)copper(II)] | A549 | 0.85 μM | [30] |
| bis[(µ2-chloro)chloro(1,10-phenanthroline)copper(II)] | B16F10 | 0.91 μM | [30] |
| [Cu(bta)(1,10-phenanthroline)(ClO4)] | Sarcoma TG180 | 7.4 μM | [31] |
| [Cu(tdp)(phen)]+ | MCF-7 | 1.6 μM | [32] |
| [Cu(tdp)(phen)]+ | MDA-MB-231 | 1.9 μM | [32] |
| [Cu(4-FH)(phen)(ClO4)2] | K562 | 1.8 μM | [10] |
| [Cu(4-FH)(phen)(ClO4)2] | K562 | 26.2 μM | [10] |
| [Cu(4-FH)(phen)(ClO4)2] | K562 | 1.6 μM | [10] |
| [Cu(4-FH)(phen)(ClO4)2] | K562 | 28 μM | [10] |
| [Cu(4-FH)(phen)(ClO4)2] | K562 | 15 μM | [10] |
| Complex 1 | PC3 | 45.58 μM | Present paper |
| Complex 1 | LNCap | 9.05 μM | Present paper |
4. Materials and Methods
4.1. Cu(II) Complex and Starting Materials
4.2. Cell Lines
4.3. MTT Assay
4.4. Flow Cytometry
4.5. Statistical Analysis
4.6. Toxicity Test in D. melanogaster
4.7. SMART Test in D. melanogaster Wings
4.8. ETT Test in D. melanogaster
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement:
Acknowledgments
Conflicts of Interest
Abbreviations
| BH | Balancer-heterozygous individuals |
| DXR | Doxorubicin |
| LD | Lethal Dose |
| ETT | Epithelial tumor test |
| FM | Frequency of mutation |
| FR | Frequency of recombination |
| HB | High Bioactivation cross |
| LATS1 | Large tumor suppressor kinase 1 |
| MH | Trans-heterozygous individuals |
| MTT | 3-(4,5-dimethylthiazol2-yl)-2,5-diphenyltetrazolium bromide |
| PCa | Prostate Cancer |
| ROS | Reactive oxygen species |
| SI | Selectivity Index |
| SMART | Somatic Mutation and Recombination Test |
| SMN | Subsequent malignant neoplasm |
| ST | Standard cross |
| TX | Toxicity Test |
References
- Akdemir, K.C.; Le, V.T.; Kim, J.M.; Killcoyne, S.; King, D.A.; Lin, Y.-P.; Tian, Y.; Inoue, A.; Amin, S.B.; Robinson, F.S.; et al. Somatic mutation distributions in cancer genomes vary with three-dimensional chromatin structure. Nat. Genet. 2020, 52, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
- Boros, E.; Dyson, P.J.; Gasser, G. Classification of Metal-Based Drugs according to Their Mechanisms of Action. Chem 2020, 6, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Cheff, D.M.; Hall, M.D. A Drug of Such Damned Nature. 1 Challenges and Opportunities in Translational Platinum Drug Research: Miniperspective. J. Med. Chem. 2017, 60, 4517–4532. [Google Scholar] [CrossRef] [PubMed]
- Vojtek, M.; Marques, M.P.; Ferreira, I.M.; Mota-Filipe, H.; Diniz, C. Anticancer activity of palladium-based complexes against triple-negative breast cancer. Drug Discov. Today 2019, 24, 1044–1058. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Elmehrath, A.O.; Afifi, A.M.; Al-Husseini, M.J.; Saad, A.M.; Wilson, N.; Shohdy, K.S.; Pilie, P.; Sonbol, M.B.; Alhalabi, O. Causes of Death Among Patients with Metastatic Prostate Cancer in the US From 2000 to 2016. JAMA Netw. Open 2021, 4, e2119568. [Google Scholar] [CrossRef]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [Green Version]
- Twomey, J.D.; Brahme, N.N.; Zhang, B. Drug-biomarker co-development in oncology—20 years and counting. Drug Resist. Updat. 2017, 30, 48–62. [Google Scholar] [CrossRef]
- Xue, D.; Zhou, X.; Qiu, J. Emerging role of NRF2 in ROS-mediated tumor chemoresistance. Biomed. Pharmacother. 2020, 131, 110676. [Google Scholar] [CrossRef]
- Paixão, D.A.; Marzano, I.M.; Jaimes, E.H.; Pivatto, M.; Campos, D.L.; Pavan, F.R.; Deflon, V.M.; Maia, P.I.D.S.; Ferreira, A.M.D.C.; Uehara, I.A.; et al. Novel copper (II) complexes with hydrazides and heterocyclic bases: Synthesis, structure and biological studies. J. Inorg. Biochem. 2017, 172, 138–146. [Google Scholar] [CrossRef]
- Tahmasvand, R.; Bayat, P.; Vahdaniparast, S.M.; Dehghani, S.; Kooshafar, Z.; Khaleghi, S.; Almasirad, A.; Salimi, M. Design and synthesis of novel 4-thiazolidinone derivatives with promising anti-breast cancer activity: Synthesis, characterization, in vitro and in vivo results. Bioorganic Chem. 2020, 104, 104276. [Google Scholar] [CrossRef]
- Yousef, T. Structural, optical, morphology characterization and DFT studies of nano sized Cu(II) complexes containing schiff base using green synthesis. J. Mol. Struct. 2020, 1215, 128180. [Google Scholar] [CrossRef]
- Simpson, P.V.; Desai, N.M.; Casari, I.; Massi, M.; Falasca, M. Metal-based antitumor compounds: Beyond cisplatin. Futur. Med. Chem. 2019, 11, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.N.; Colacci, A.; Corvi, R.; Vaccari, M.; Aguila, M.C.; Corvaro, M.; Delrue, N.; Desaulniers, D.; Ertych, N.; Jacobs, A.; et al. Chemical carcinogen safety testing: OECD expert group international consensus on the development of an integrated approach for the testing and assessment of chemical non-genotoxic carcinogens. Arch. Toxicol. 2020, 94, 2899–2923. [Google Scholar] [CrossRef] [PubMed]
- Baran, E.J. Trace elements supplementation: Recent advances and perspectives. Mini-Rev. Med. Chem. 2004, 4, 1–9. [Google Scholar] [CrossRef]
- Graf, U.; Würgler, F.E.; Katz, A.J.; Frei, H.; Juon, H.; Hall CBKale, P.G. Somatic mutation and recombination test in Drosophila melanogaster. Environ. Mutagen. 1984, 6, 153–188. [Google Scholar] [CrossRef]
- Spanó, M.A.; Frei, H.; Würgler, F.E.; Graf, U. Recombinagenic activity of four compounds in the standard and high bioactivation crosses of Drosophila melanogaster in the wing spot test. Mutagenesis 2001, 16, 385–394. [Google Scholar] [CrossRef] [Green Version]
- El-Hefny, I.M.; Hozayen, W.; AlSenosy, N.; Basal, W.; Ahmed, A.; Diab, A. Evaluation of genotoxicity of three food preservatives in drosophila melanogaster using smart and comet assays. J. Microbiol. Biotechnol. Food Sci. 2020, 10, 38–41. [Google Scholar] [CrossRef]
- Li, H. Single-cell RNA sequencing in Drosophila: Technologies and applications. WIREs Dev. Biol. 2021, 10, e396. [Google Scholar] [CrossRef]
- Ertuğrul, H.; Yalçın, B.; Güneş, M.; Kaya, B. Ameliorative effects of melatonin against nano and ionic cobalt induced genotoxicity in two in vivo Drosophila assays. Drug Chem. Toxicol. 2020, 43, 279–286. [Google Scholar] [CrossRef]
- Lima, P.M.A.P.; Orsoli, P.C.; Araújo, T.; Cardoso, D.B.; Nepomuceno, J.C. Effects of a Carbonated Soft Drink on Epitheial Tumor Incidence in Drosophila melanogaster. J. Pharm. Pharmacol. 2018, 6, 240–247. [Google Scholar] [CrossRef]
- da Silva, T.T.; Martins, J.B.; Lopes, M.D.S.D.B.; de Almeida, P.M.; Sá, J.L.S.; Martins, F.A. Modulating effect of DL-kavain on the mutagenicity and carcinogenicity induced by doxorubicin in Drosophila melanogaster. J. Toxicol. Environ. Health Part A 2021, 84, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, M.A.; Orsolin, P.C.; Silva-Oliveira, R.G.; Nepomuceno, J.C.; Spanó, M.A. Assessment of the carcinogenic potential of high intense-sweeteners through the test for detection of epithelial tumor clones (warts) in Drosophila melanogaster. Food Chem. Toxicol. 2017, 101, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Demir, F.T. In vivo effects of 1,4-dioxane on genotoxic parameters and behavioral alterations in Drosophila melanogaster. J. Toxicol. Environ. Health Part A 2022, 85, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Frei, H.; Würgler, F. Statistical methods to decide whether mutagenicity test data from Drosophila assays indicate a positive, negative, or inconclusive result. Mutat. Res. Mutagen. Relat. Subj. 1988, 203, 297–308. [Google Scholar] [CrossRef]
- Fung, C.; Travis, L.B. Risk of solid cancer after chemotherapy. Nat. Rev. Urol. 2018, 15, 660–661. [Google Scholar] [CrossRef]
- Silva, P.P.; Guerra, W.; Silveira, J.N.; Ferreira, A.M.D.C.; Bortolotto, T.; Fischer, F.L.; Terenzi, H.; Neves, A.; Pereira-Maia, E.C. Two New Ternary Complexes of Copper(II) with Tetracycline or Doxycycline and 1,10-Phenanthroline and Their Potential as Antitumoral: Cytotoxicity and DNA Cleavage. Inorg. Chem. 2011, 50, 6414–6424. [Google Scholar] [CrossRef]
- Li, D.-D.; Yagüe, E.; Wang, L.-Y.; Dai, L.-L.; Yang, Z.-B.; Zhi, S.; Zhang, N.; Zhao, X.-M.; Hu, Y.-H. Novel Copper Complexes That Inhibit the Proteasome and Trigger Apoptosis in Triple-Negative Breast Cancer Cells. ACS Med. Chem. Lett. 2019, 10, 1328–1335. [Google Scholar] [CrossRef]
- Lopes, J.C.; Botelho, F.V.; Silva, M.J.B.; Silva, S.F.; Polloni, L.; Machado, P.H.A.; de Souza, T.R.; Goulart, L.R.; Caldeira, P.P.S.; Maia, E.C.P.; et al. In vitro and in vivo antitumoral activity of a ternary copper (II) complex. Biochem. Biophys. Res. Commun. 2020, 533, 1021–1026. [Google Scholar] [CrossRef]
- Mroueh, M.; Daher, C.; Hariri, E.; Demirdjian, S.; Isber, S.; Choi, E.S.; Mirtamizdoust, B.; Hammud, H.H. Magnetic property, DFT calculation, and biological activity of bis[(μ2-chloro)chloro(1,10-phenanthroline)copper(II)] complex. Chem. Interact. 2015, 231, 53–60. [Google Scholar] [CrossRef]
- Polloni, L.; Silva, A.C.D.S.; Teixeira, S.C.; Azevedo, F.V.P.D.V.; Zóia, M.A.P.; da Silva, M.S.; Lima, P.M.A.P.; Correia, L.I.V.; Almeida, J.D.C.; da Silva, C.V.; et al. Action of copper(II) complex with β-diketone and 1,10-phenanthroline (CBP-01) on sarcoma cells and biological effects under cell death. Biomed. Pharmacother. 2019, 112, 108586. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, V.; Riyasdeen, A.; Rajendiran, V.; Palaniandavar, M.; Krishnamurthy, H.; Alshatwi, A.; Akbarsha, M. Induction of Redox-Mediated Cell Death in ER-Positive and ER-Negative Breast Cancer Cells by a Copper(II)-Phenolate Complex: An In Vitro and In Silico Study. Molecules 2020, 25, 4504. [Google Scholar] [CrossRef] [PubMed]
- Richardsen, E.; Andersen, S.; Al-Saad, S.; Rakaee, M.; Nordby, Y.; Pedersen, M.I.; Ness, N.; Grindstad, T.; Movik, I.; Dønnem, T.; et al. Evaluation of the proliferation marker Ki-67 in a large prostatectomy cohort. PLoS ONE 2017, 12, e0186852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Yao, N.; Liao, X.; Xie, X.; Wu, Y.; Fan, C.; Zhao, J.; Li, G. Effects of metal ions on the structure and activity of a human anti-cyclin D1 single-chain variable fragment AD5. Mol. Med. Rep. 2017, 16, 1314–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheinberg, T.; Kench, J.; Stockler, M.; Mahon, K.L.; Sebastian, L.; Stricker, P.; Joshua, A.M.; Woo, H.; Thanigasalam, R.; Ahmadi, N.; et al. Pharmacodynamics effects of CDK4/6 inhibitor LEE011 (ribociclib) in high-risk, localised prostate cancer: A study protocol for a randomised controlled phase II trial (LEEP study: LEE011 in high-risk, localised Prostate cancer). BMJ Open 2020, 10, e033667. [Google Scholar] [CrossRef] [Green Version]
- Jardim, D.L.; Millis, S.Z.; Ross, J.S.; Woo, M.S.-A.; Ali, S.M.; Kurzrock, R. Landscape of Cyclin Pathway Genomic Alterations Across 5,356 Prostate Cancers: Implications for Targeted Therapeutics. Oncologist 2021, 26, e715–e718. [Google Scholar] [CrossRef]
- Koh, K.; Evans, J.M.; Hendricks, J.C.; Sehgal, A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc. Natl. Acad. Sci. USA 2006, 103, 13843–13847. [Google Scholar] [CrossRef] [Green Version]
- Marcos, R.; Carmona, E.R. The Wing-Spot and the Comet Tests as Useful Assays for Detecting Genotoxicity in Drosophila. Genotoxic. Assess. 2019, 2031, 337–348. [Google Scholar] [CrossRef]
- González SE, F.; Anguiano, E.A.; Herrera, A.M.; Calzada, D.E.; Pichardo, C.O. Cytotoxic, pro-apoptotic, pro-oxidant, and non-genotoxic activities of a novel copper (II) complex against human cervical cancer. Toxicology 2013, 314, 155–165. [Google Scholar] [CrossRef]
- Cadavid-Vargas, J.F.; Arnal, P.M.; Mojica Sepúlveda, R.D.; Rizzo, A.; Soria, D.B.; Di Virgilio, A.L. Copper complex with sulfamethazine and 2, 2′-bipyridine supported on mesoporous silica microspheres improves its antitumor action toward human osteosarcoma cells: Cyto-and genotoxic effects. BioMetals 2019, 32, 21–32. [Google Scholar] [CrossRef]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno, J.C. Using the Drosophila melanogaster to Assessment Carcinogenic Agents through the Test for Detection of Epithelial Tumor Clones (Warts). Adv. Technol. Biol. Med. 2015, 3, 1–8. [Google Scholar] [CrossRef]
- Ahmad, M.; Suhaimi, S.N.; Chu, T.-L.; Aziz, N.A.; Kornain, N.-K.M.; Samiulla, D.S.; Lo, K.-W.; Ng, C.-H.; Khoo, A.S.-B. Ternary copper(II) complex: NCI60 screening, toxicity studies, and evaluation of efficacy in xenograft models of nasopharyngeal carcinoma. PLoS ONE 2018, 13, e0191295. [Google Scholar] [CrossRef] [Green Version]
- Arjmand, F.; Parveen, S.; Afzal, M.; Shahid, M. Synthesis, characterization, biological studies (DNA binding, cleavage, antibacterial and topoisomerase I) and molecular docking of copper (II) benzimidazole complexes. J. Photochem. Photobiol. B Biol. 2012, 114, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, S.; Afzal, M.; Arjmand, F. New heterobimetallic CuII–Sn2IV complex as potential topoisomerase I inhibitor: In vitro DNA binding, cleavage and cytotoxicity against human cancer cell lines. J. Photochem. Photobiol. B Biol. 2012, 115, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Vutey, V.; Castelli, S.; D’Annessa, I.; Sâmia, L.B.; Souza-Fagundes, E.M.; Beraldo, H.; Desideri, A. Human topoisomerase IB is a target of a thiosemicarbazone copper (II) complex. Arch. Biochem. Biophys. 2016, 606, 34–40. [Google Scholar] [CrossRef]
- Machado, N.M.; Lopes, J.C.; Saturnino, R.S.; Fagan, E.B.; Nepomuceno, J.C. Lack of mutagenic effect by multi-walled functionalized carbon nanotubes in the somatic cells of Drosophila melanogaster. Food Chem. Toxicol. 2013, 62, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Orsolin, P.C.; Silva-Oliveira, R.G.; Nepomuceno, J.C. Modulating effect of synthetic statins against damage induced by doxorubicin in somatic cells of Drosophila melanogaster. Food Chem. Toxicol. 2015, 81, 111–119. [Google Scholar] [CrossRef]
- Silva-Oliveira, R.; Orsolin, P.; Nepomuceno, J. Modulating effect of losartan potassium on the mutagenicity and recombinogenicity of doxorubicin in somatic cells of Drosophila melanogaster. Food Chem. Toxicol. 2016, 95, 211–218. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Pereira, F.D.C.; Lima, B.A.; de Lima, A.P.; Pires, W.C.; Monteiro, T.; Magalhães, L.F.; Costa, W.; Graminha, A.E.; Batista, A.A.; Ellena, J.; et al. Cis-[RuCl(BzCN)(N–N)(P–P)]PF6 complexes: Synthesis and in vitro antitumor activity. J. Inorg. Biochem. 2015, 149, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Graf, U.; van Schaik, N. Improved high bioactivation cross for the wing somatic mutation and recombination test in Drosophila melanogaster. Mutat. Res. Environ. Mutagen. Relat. Subj. 1992, 271, 59–67. [Google Scholar] [CrossRef]
- Guzmán-Rincón, J.; Graf, U. Drosophila melanogaster somatic mutation and recombination test as a biomonitor. Environ. Sci. Res. 1995, 50, 169–182. [Google Scholar]
- Andrade, H.H.R.d.; Lehmann, M. Teste para detecção de mutação e recombinação somática (SMART) em Drosophila melanogaster. Mutagênese Ambient. 2003, 1, 281–307. [Google Scholar]
- Kastenbaum, M.A.; Bowman, K. Tables for determining the statistical significance of mutation frequencies. Mutat. Res. Mol. Mech. Mutagen. 1970, 9, 527–549. [Google Scholar] [CrossRef]
- Abraham, S.K. Antigenotoxicity of coffee in the Drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef]
- Siam, R.; Harada, R.; Cadieux, C.; Battat, R.; Vadnais, C.; Nepveu, A. Transcriptional activation of the Lats1 tumor suppressor gene in tumors of CUX1 transgenic mice. Mol. Cancer 2009, 8, 60. [Google Scholar] [CrossRef]





| Treatments | Number of Flies (n) | Spots per Fly (N° of Spots); Statistical Diagnosis a | Spots with mwh Clone c (n) | Frequency of Clone Formation/ 105 Cells per Cell Division d | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DXR (mM) | Complex 1 (mM) | Small Single (1–2 Cells) b m = 2 | Large Single (>2 Cells) b m = 5 | Twin m = 5 | Total Spots m = 2 | Observed | Control Corrected | ||||||||||
| mwh/flr3 | |||||||||||||||||
| ST Cross | |||||||||||||||||
| 0 | 0 | 60 | 0.40 | (24) | 0.03 | (2) | 0.02 | (1) | 0.45 | (27) | 27 | 0.92 | |||||
| 0.4 | 0 | 60 | 2.62 | (157) | + | 4.52 | (271) | + | 5.33 | (320) | + | 12.47 | (748) | + | 697 | 23.80 | 22.80 |
| 0 | 0.015 | 60 | 0.32 | (19) | - | 0.03 | (2) | i | 0.08 | (5) | i | 0.43 | (26) | - | 23 | 0.79 | −0.14 |
| 0 | 0.031 | 60 | 0.35 | (21) | - | 0.05 | (3) | i | 0.02 | (1) | i | 0.42 | (25) | - | 23 | 0.79 | −0.14 |
| 0 | 0.062 | 60 | 0.35 | (21) | - | 0.03 | (2) | i | 0.03 | (2) | i | 0.42 | (25) | - | 25 | 0.85 | −0.07 |
| 0 | 0.125 | 60 | 0.23 | (14) | - | 0.05 | (3) | i | 0.12 | (7) | i | 0.40 | (24) | - | 21 | 0.72 | −0.20 |
| 0 | 0.250 | 60 | 0.25 | (15) | - | 0.00 | (0) | i | 0.12 | (7) | i | 0.37 | (22) | - | 16 | 0.55 | −0.38 |
| HB Cross | |||||||||||||||||
| 0 | 0 | 60 | 0.62 | (37) | 0.20 | (12) | 0.00 | (0) | 0.82 | (49) | 48 | 1.64 | |||||
| 0.4 | 0 | 60 | 1.30 | (78) | + | 1.10 | (606) | 1.98 | (119) | + | 13.38 | (803) | + | 782 | 26.71 | 25.07 | |
| 0 | 0.015 | 60 | 0.27 | (16) | - | 0.13 | (8) | - | 0.03 | (2) | i | 0.43 | (26) | - | 26 | 0.89 | −0.75 |
| 0 | 0.031 | 60 | 0.30 | (18) | - | 0.12 | (7) | - | 0.02 | (1) | i | 0.43 | (26) | - | 26 | 0.89 | −0.75 |
| 0 | 0.062 | 60 | 0.38 | (23) | - | 0.03 | (2) | i | 0.00 | (0) | i | 0.42 | (25) | - | 25 | 0.85 | −0.89 |
| 0 | 0.125 | 60 | 0.22 | (13) | - | 0.10 | (6) | - | 0.07 | (4) | i | 0.38 | (23) | - | 23 | 0.79 | −0.85 |
| 0 | 0.250 | 60 | 0.10 | (6) | - | 0.10 | (6) | - | 0.00 | (0) | i | 0.20 | (12) | - | 12 | 0.41 | −1.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bontempo, N.J.d.S.; Paixão, D.A.; Lima, P.M.A.P.; Barros, D.C.T.; Borges, D.S.; Orsolin, P.C.; Martins, I.C.; Machado, P.H.A.; Lino, R.C.; Souza, T.R.d.; et al. Copper(II) Complex Containing 4-Fluorophenoxyacetic Acid Hydrazide and 1,10-Phenanthroline: A Prostate Cancer Cell-Selective and Low-Toxic Copper(II) Compound. Molecules 2022, 27, 7097. https://doi.org/10.3390/molecules27207097
Bontempo NJdS, Paixão DA, Lima PMAP, Barros DCT, Borges DS, Orsolin PC, Martins IC, Machado PHA, Lino RC, Souza TRd, et al. Copper(II) Complex Containing 4-Fluorophenoxyacetic Acid Hydrazide and 1,10-Phenanthroline: A Prostate Cancer Cell-Selective and Low-Toxic Copper(II) Compound. Molecules. 2022; 27(20):7097. https://doi.org/10.3390/molecules27207097
Chicago/Turabian StyleBontempo, Nayara Júnia de Souza, Drielly Aparecida Paixão, Paula Marynella Alves Pereira Lima, Deysse Carla Tolentino Barros, Dayanne Silva Borges, Priscila Capelari Orsolin, Isabella Castro Martins, Pedro Henrique Alves Machado, Ricardo Campos Lino, Tiago Rodrigues de Souza, and et al. 2022. "Copper(II) Complex Containing 4-Fluorophenoxyacetic Acid Hydrazide and 1,10-Phenanthroline: A Prostate Cancer Cell-Selective and Low-Toxic Copper(II) Compound" Molecules 27, no. 20: 7097. https://doi.org/10.3390/molecules27207097
APA StyleBontempo, N. J. d. S., Paixão, D. A., Lima, P. M. A. P., Barros, D. C. T., Borges, D. S., Orsolin, P. C., Martins, I. C., Machado, P. H. A., Lino, R. C., Souza, T. R. d., Ramos, L. M. S., Teixeira, S. C., Siqueira, R. P., Goulart Filho, L. R., Guerra, W., Oliveira Júnior, R. J. d., & Araújo, T. G. d. (2022). Copper(II) Complex Containing 4-Fluorophenoxyacetic Acid Hydrazide and 1,10-Phenanthroline: A Prostate Cancer Cell-Selective and Low-Toxic Copper(II) Compound. Molecules, 27(20), 7097. https://doi.org/10.3390/molecules27207097