


Inhibitory Effect of Isoliquiritigenin in Niemann-Pick C1-Like 1-Mediated Cholesterol Uptake
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
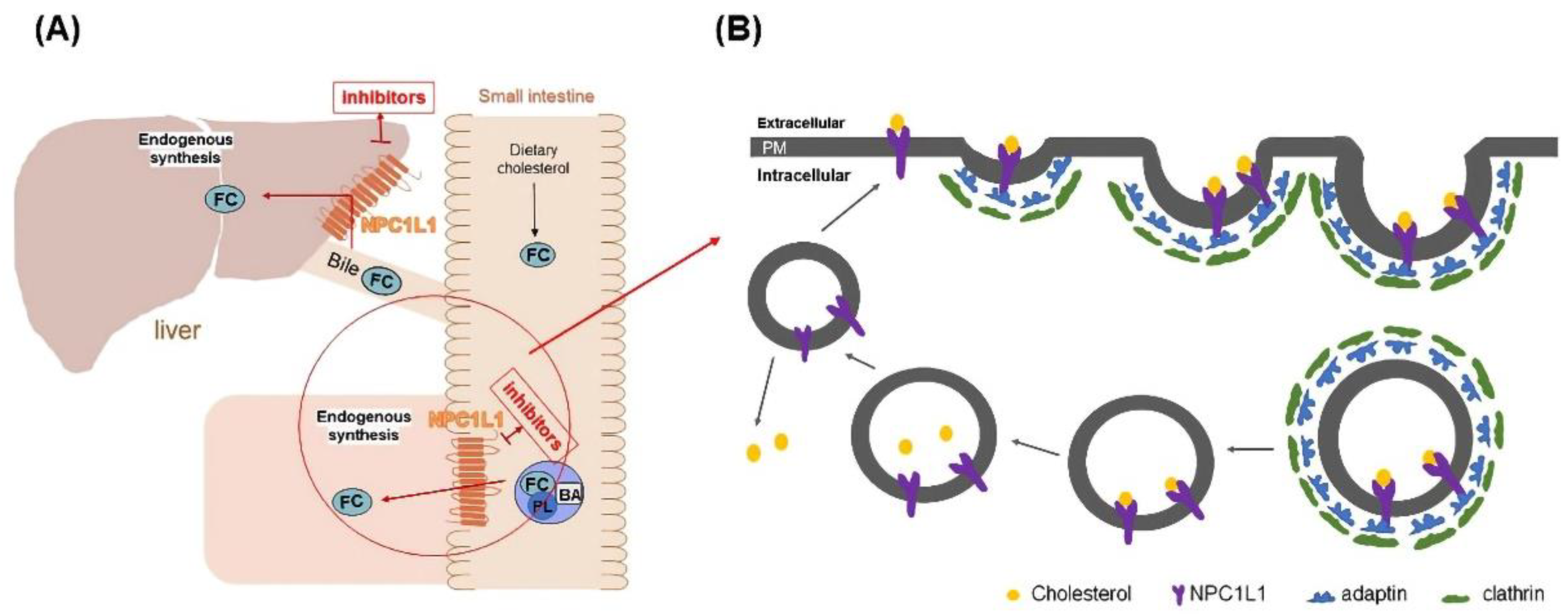
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Western Blot Analysis
2.4.1. Extraction and Preparation of Total Cellular Proteins
2.4.2. Protein Blot Analysis
2.5. Molecular Docking Methods
2.6. NBD-Cholesterol Uptake Assays
2.7. Filipin Bacteriocin Staining
2.8. Molecular Kinetic Analysis
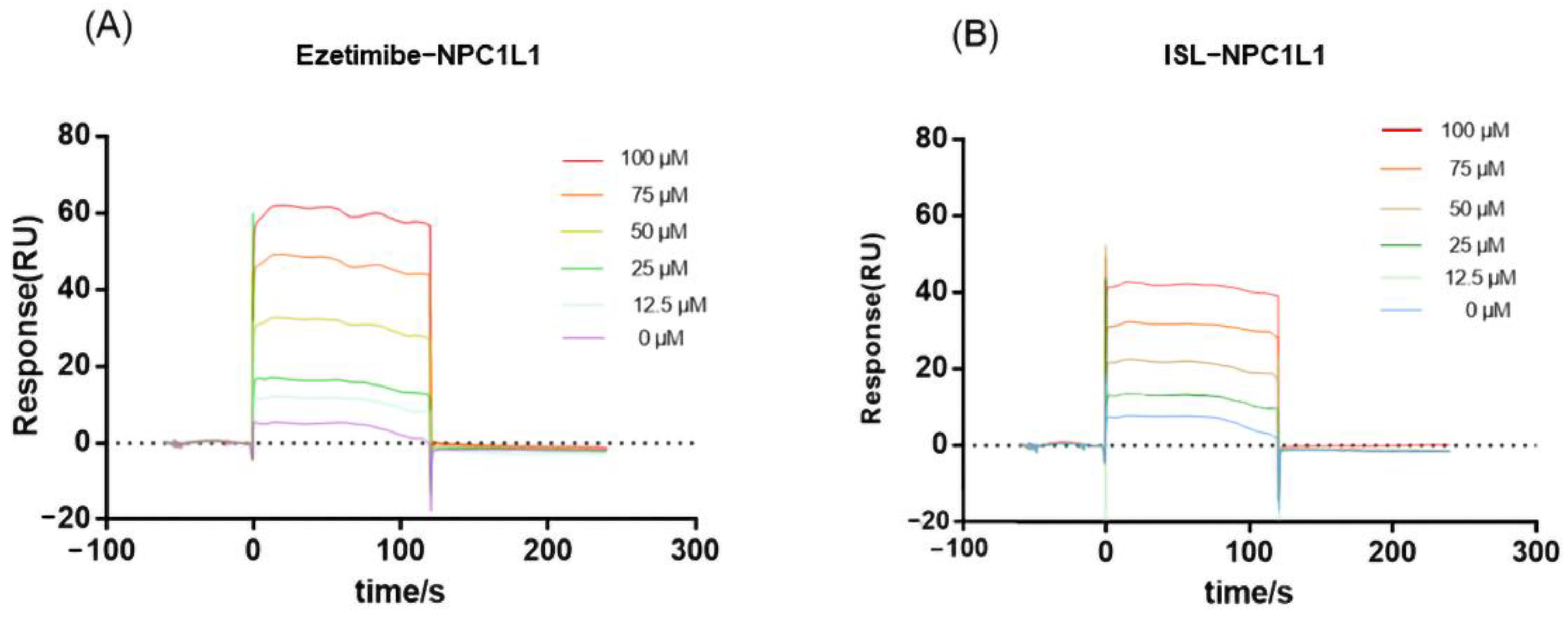
2.9. SPR
2.10. Statistical Analysis
3. Results
3.1. SPR Results Show an Interaction between ISL and NPC1L1
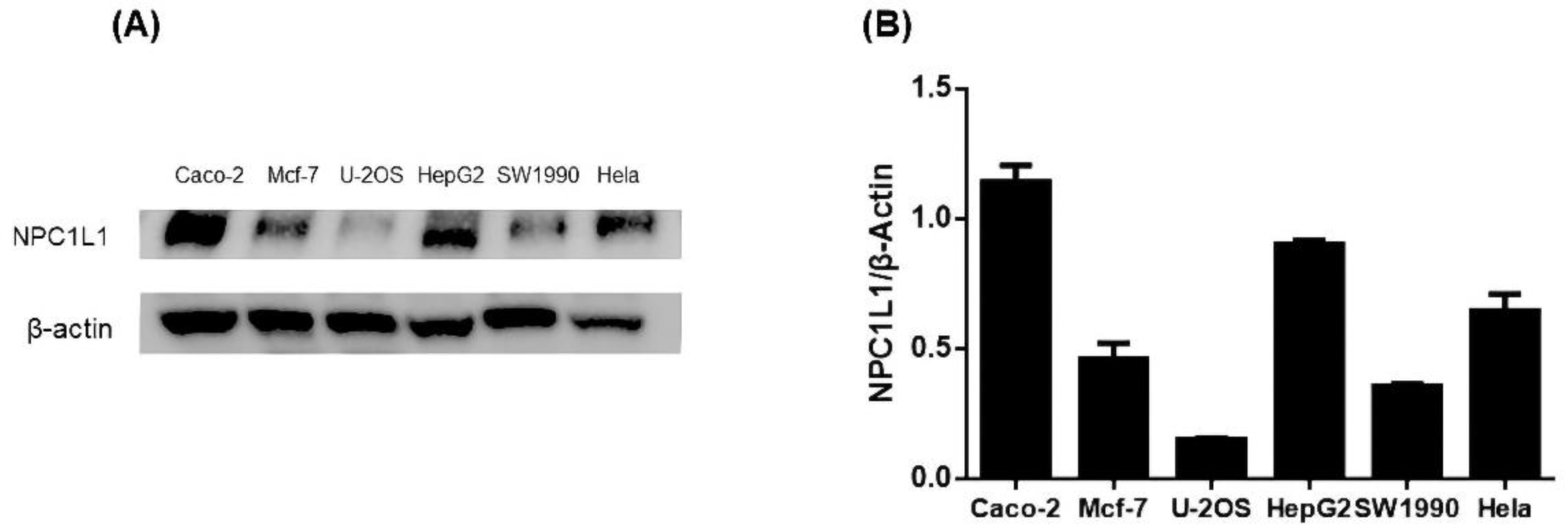
3.2. Results of Protein Immunoblot Analysis Defining the Cell Model
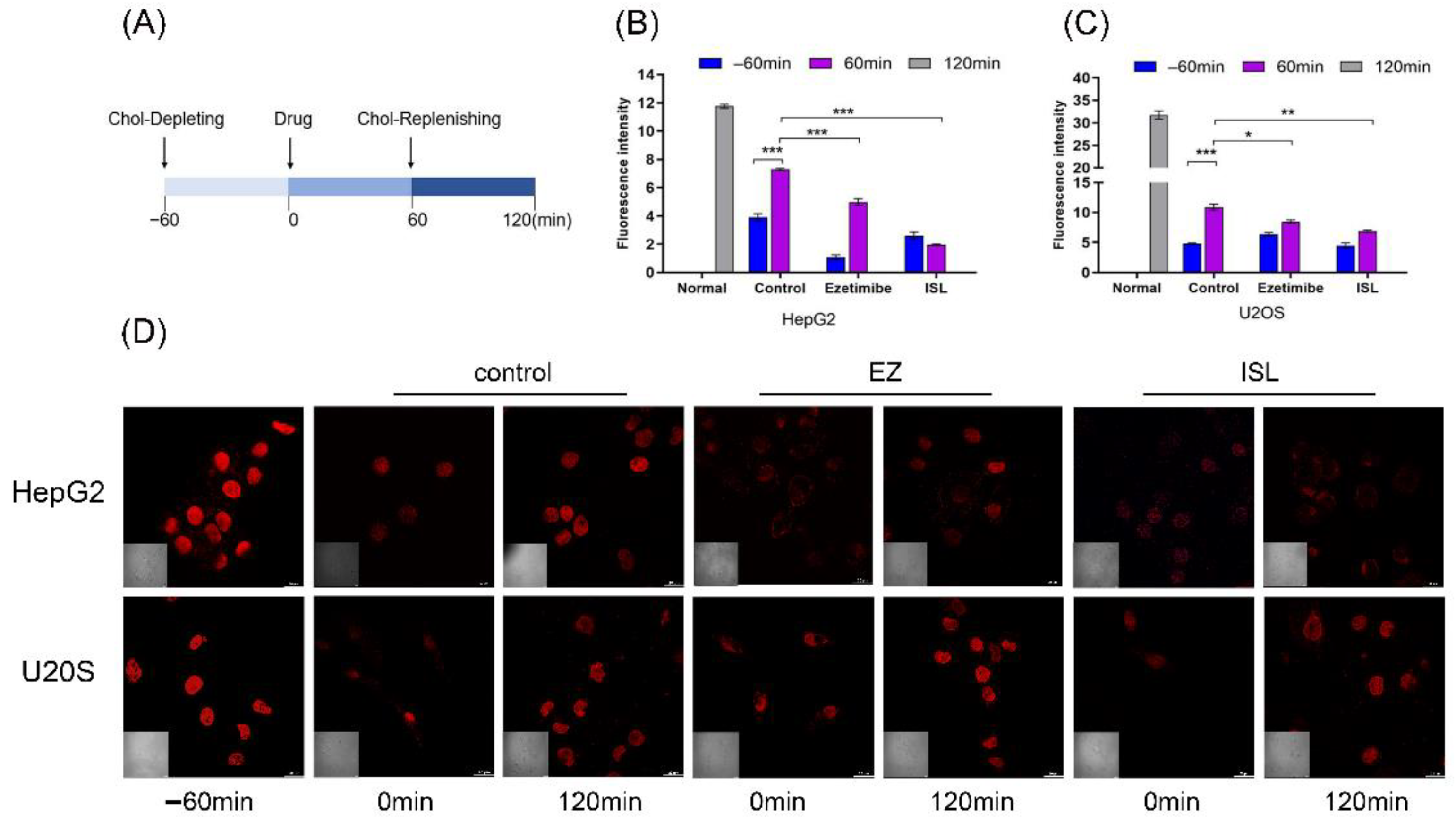
3.3. ISL Inhibited Uptake of Cholesterol in HepG2/Caco-2 Cells
3.4. Filipin Staining Results Showed That ISL Reduced Cellular Uptake of Cholesterol
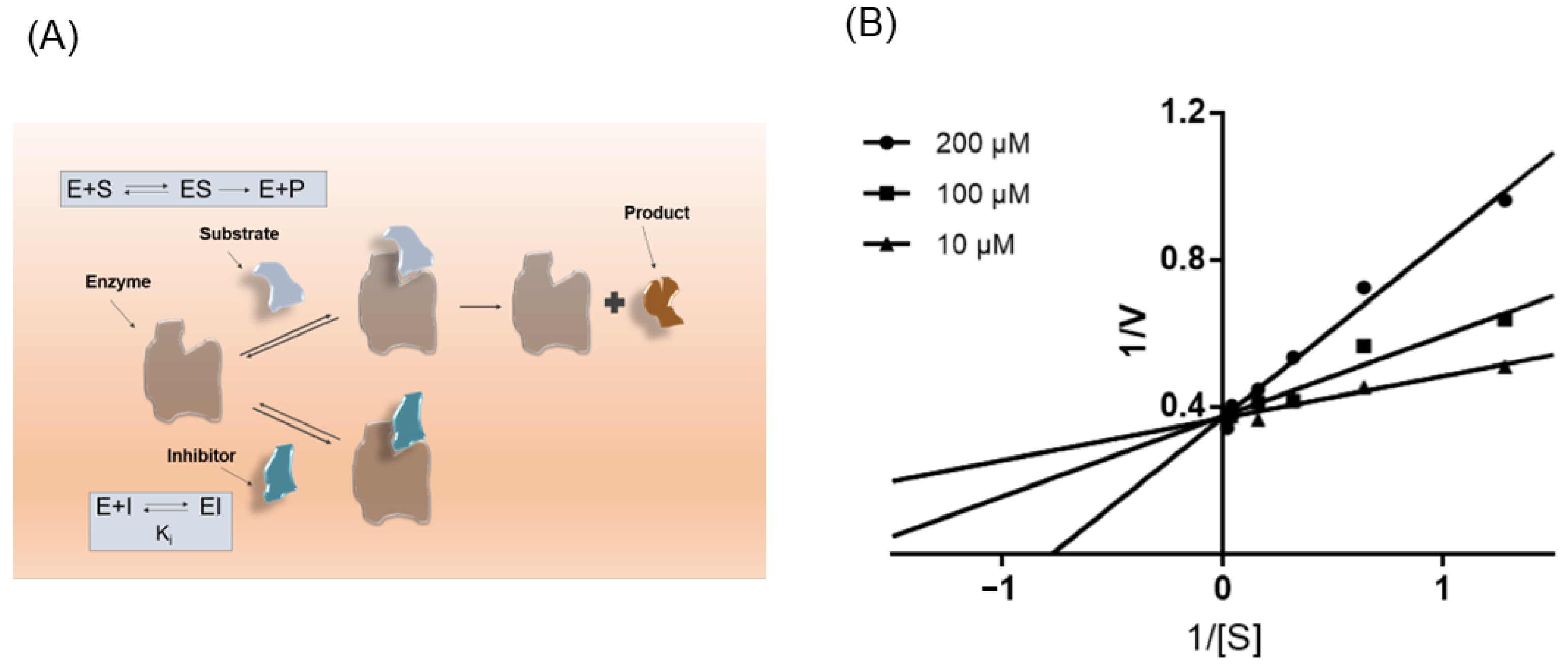
3.5. ISL Is a Competitive Inhibitor of NPC1L1
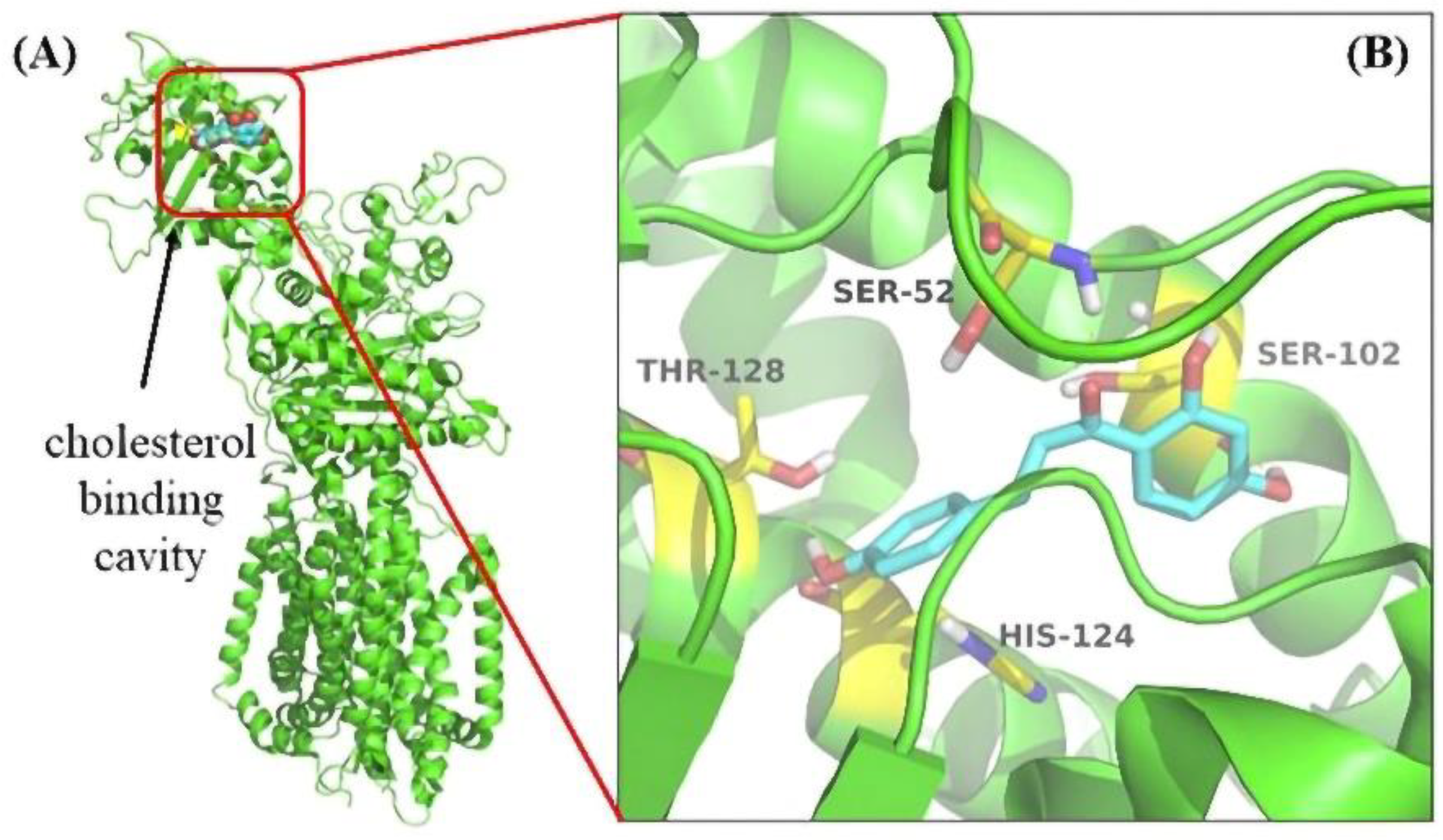
3.6. Molecular Docking Revealed the Binding Pocket of NPC1L1 with ISL
3.7. ISL Reduced the Expression of NPC1L1 in HepG2 Cells
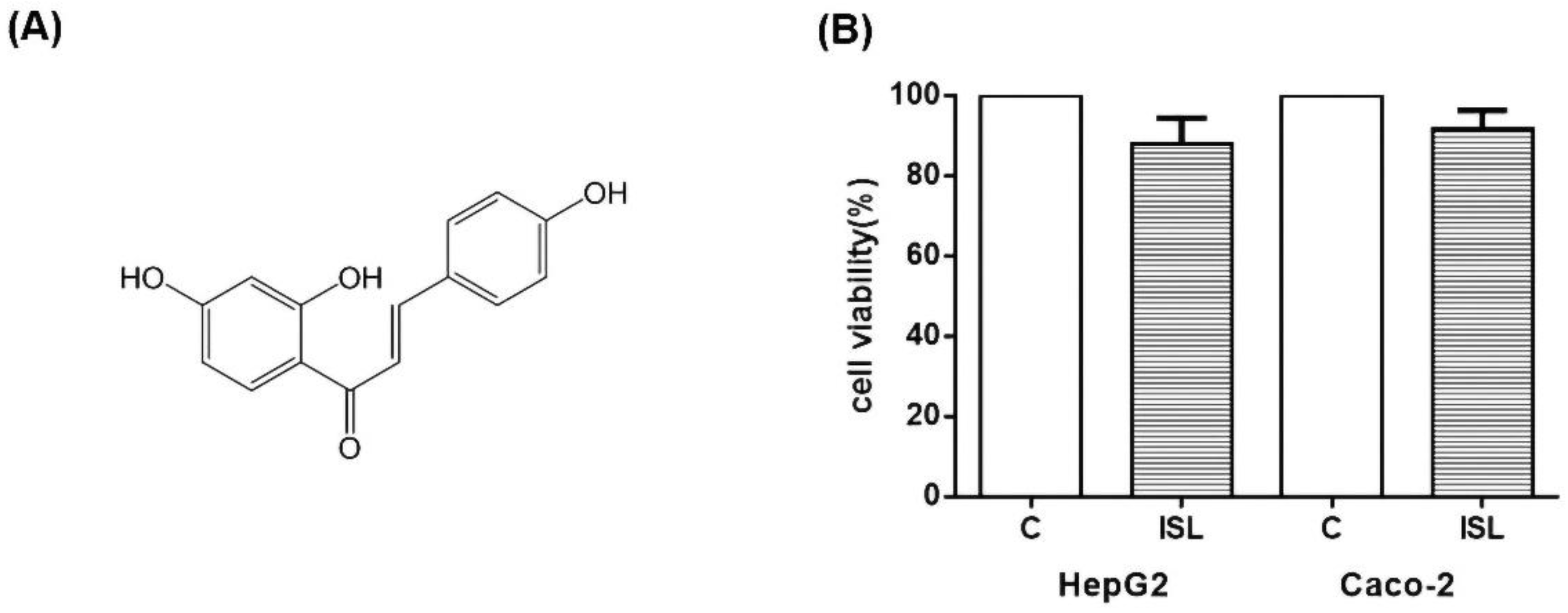
3.8. ISL Has Low Cytotoxicity against HepG2/Caco-2 Cells
4. Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gadgil, M.D.; Anderson, C.A.M.; Kandula, N.R.; Kanaya, A.M. Dietary patterns in Asian Indians in the United States: An analysis of the metabolic syndrome and atherosclerosis in South Asians Living in America study. J. Acad. Nutr. Diet. 2014, 114, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Gielen, S.; Landmesser, U. The Year in Cardiology 2013: Cardiovascular disease prevention. Eur. Heart J. 2014, 35, 307–312. [Google Scholar] [CrossRef]
- Lu, K.; Lee, M.-H.; Patel, S.B. Dietary cholesterol absorption; more than just bile. Trends Endocrinol. Metab. 2001, 12, 314–320. [Google Scholar] [CrossRef]
- Kruit, J.K.; Groen, A.K.; van Berkel, T.J.; Kuipers, F. Emerging roles of the intestine in control of cholesterol metabolism. World J. Gastroenterol. 2006, 12, 6429–6439. [Google Scholar] [CrossRef]
- Gylling, H.; Miettinen, T.A. The effect of cholesterol absorption inhibition on low density lipoprotein cholesterol level. Atherosclerosis 1995, 117, 305–308. [Google Scholar] [CrossRef]
- Jia, L.; Betters, J.L.; Yu, L. Niemann-pick C1-like 1 (NPC1L1) protein in intestinal and hepatic cholesterol transport. Annu. Rev. Physiol. 2011, 73, 239–259. [Google Scholar] [CrossRef] [Green Version]
- Betters, J.L.; Yu, L. NPC1L1 and cholesterol transport. FEBS Lett. 2010, 584, 2740–2747. [Google Scholar] [CrossRef] [Green Version]
- Altmann, S.W.; Davis, H.R., Jr.; Zhu, L.J.; Yao, X.; Hoos, L.M.; Tetzloff, G.; Iyer, S.P.N.; Maguire, M.; Golovko, A.; Zeng, M.; et al. Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 2004, 303, 1201–1204. [Google Scholar] [CrossRef] [Green Version]
- Kosoglou, T.; Statkevich, P.; Johnson-Levonas, A.O.; Paolini, J.F.; Bergman, A.J.; Alton, K.B. Ezetimibe: A review of its metabolism, pharmacokinetics and drug interactions. Clin. Pharmacokinet. 2005, 44, 467–494. [Google Scholar] [CrossRef]
- Ge, L.; Wang, J.; Qi, W.; Miao, H.H.; Cao, J.; Qu, Y.X.; Li, B.-L.; Song, B.-L. The cholesterol absorption inhibitor ezetimibe acts by blocking the sterol-induced internalization of NPC1L1. Cell Metab. 2008, 7, 508–519. [Google Scholar] [CrossRef]
- Nutescu, E.A.; Shapiro, N.L. Ezetimibe: A selective cholesterol absorption inhibitor. Pharmacotherapy 2003, 23, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Calvo, M.; Lisnock, J.M.; Bull, H.G.; Hawes, B.E.; Burnett, D.A.; Braun, M.P.; Crona, J.H.; Davis, H.R., Jr.; Dean, D.C.; Detmers, P.A.; et al. The target of ezetimibe is Niemann-Pick C1-Like 1 (NPC1L1). Proc. Natl. Acad. Sci. USA 2005, 102, 8132–8137. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Liu, W.; Zeng, J.; Meng, J.; Jiang, H.; Wang, J.; Xing, D. Niemann-Pick C1-Like 1 inhibitors for reducing cholesterol absorption. Eur. J. Med. Chem. 2022, 230, 114111. [Google Scholar] [CrossRef]
- Yao, H.; Liu, J.; Xu, S.; Zhu, Z.; Xu, J. The structural modification of natural products for novel drug discovery. Expert Opin. Drug Discov. 2017, 12, 121–140. [Google Scholar] [CrossRef]
- Chopra, B.; Dhingra, A.K. Natural products: A lead for drug discovery and development. Phytother. Res. 2021, 35, 4660–4702. [Google Scholar] [CrossRef] [PubMed]
- Robles, O.; Romo, D. Chemo-and site-selective derivatizations of natural products enabling biological studies. Nat. Prod. Rep. 2014, 31, 318–334. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Shibata, S. A drug over the millennia: Pharmacognosy, chemistry, and pharmacology of licorice. Yakugaku Zasshi 2000, 120, 849–862. [Google Scholar] [CrossRef]
- Honda, H.; Nagai, Y.; Matsunaga, T.; Okamoto, N.; Watanabe, Y.; Tsuneyama, K.; Hayashi, H.; Fujii, I.; Ikutani, M.; Hirai, Y.; et al. Isoliquiritigenin is a potent inhibitor of NLRP3 inflammasome activation and diet-induced adipose tissue inflammation. J. Leukoc. Biol. 2014, 96, 1087–1100. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, C.Y.; Bai, L.P.; Pan, H.D.; Shu, L.M.; Kong, A.N.; Leung, E.L.-H.; Liu, L.; Li, T. Flavonoids derived from liquorice suppress murine macrophage activation by up-regulating heme oxygenase-1 independent of Nrf2 activation. Int. Immunopharmacol. 2015, 28, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.K.; Son, S.H.; Park, K.K.; Park, J.H.; Lim, S.S.; Chung, W.Y. Isoliquiritigenin inhibits tumor growth and protects the kidney and liver against chemotherapy-induced toxicity in a mouse xenograft model of colon carcinoma. J. Pharmacol. Sci. 2008, 106, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; Du, Q.; Peng, C.; Wang, N.; Tang, H.; Xie, X.; Shen, J.; Chen, J. A Review: The Pharmacology of Isoliquiritigenin. Phytother. Res. 2015, 29, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Kim, T.H.; Kim, Y.W.; Yang, Y.M.; Ryu, D.H.; Hwang, S.J.; Lee, J.R.; Kim, S.C.; Kim, S.G. Inhibition of liver X receptor-alpha-dependent hepatic steatosis by isoliquiritigenin, a licorice antioxidant flavonoid, as mediated by JNK1 inhibition. Free Radic. Biol. Med. 2010, 49, 1722–1734. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Choe, Y.G.; Kim, J.H.; Chang, K.T.; Lee, H.S.; Lee, D.S. Isoliquiritigenin impairs insulin signaling and adipocyte differentiation through the inhibition of protein-tyrosine phosphatase 1B oxidation in 3T3-L1 preadipocytes. Food Chem. Toxicol. 2016, 93, 5–12. [Google Scholar] [CrossRef]
- Du, F.; Gesang, Q.; Cao, J.; Qian, M.; Ma, L.; Wu, D.; Yu, H. Isoliquiritigenin Attenuates Atherogenesis in Apolipoprotein E-Deficient Mice. Int. J. Mol. Sci. 2016, 17, 1932. [Google Scholar] [CrossRef] [Green Version]
- Kamisoyama, H.; Honda, K.; Tominaga, Y.; Yokota, S.; Hasegawa, S. Investigation of the anti-obesity action of licorice flavonoid oil in diet-induced obese rats. Biosci. Biotechnol. Biochem. 2008, 72, 3225–3231. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, K.; Kitano, M.; Kishida, H.; Hidaka, T.; Nabae, K.; Kawabe, M.; Hosoe, K. 90-Day repeated-dose toxicity study of licorice flavonoid oil (LFO) in rats. Food Chem. Toxicol. 2008, 46, 2349–2357. [Google Scholar] [CrossRef]
- Qi, J.; Cui, J.; Mi, B.; Yan, X.; Xu, W.; Ma, H.; Zhang, Q.; Xu, F. Isoliquiritigenin Inhibits Atherosclerosis by Blocking TRPC5 Channel Expression. Cardiovasc. Ther. 2020, 2020, 1926249. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Song, Z.; Wang, X.; Xue, J.; Xing, D. One-step modification to identify dual-inhibitors targeting both pancreatic triglyceride lipase and Niemann-Pick C1-like 1. Eur. J. Med. Chem. 2021, 216, 113358. [Google Scholar] [CrossRef]
- Thilavech, T.; Adisakwattana, S. Cyanidin-3-rutinoside acts as a natural inhibitor of intestinal lipid digestion and absorption. BMC Complement. Altern. Med. 2019, 19, 242. [Google Scholar] [CrossRef]
- Davies, J.P.; Scott, C.; Oishi, K.; Liapis, A.; Ioannou, Y.A. Inactivation of NPC1L1 causes multiple lipid transport defects and protects against diet-induced hypercholesterolemia. J. Biol. Chem. 2005, 280, 12710–12720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, D.; Ohlsson, L.; Duan, R.D. Curcumin inhibits cholesterol uptake in Caco-2 cells by down-regulation of NPC1L1 expression. Lipids Health Dis. 2010, 9, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.J.; Sun, L.P.; Feramisco, J.D.; Brown, M.S.; Goldstein, J.L. Cholesterol addition to ER membranes alters conformation of SCAP, the SREBP escort protein that regulates cholesterol metabolism. Mol. Cell 2002, 10, 237–245. [Google Scholar] [CrossRef]
- Zhang, J.H.; Ge, L.; Qi, W.; Zhang, L.; Miao, H.H.; Li, B.L.; Yang, M.; Song, B.-L. The N-terminal domain of NPC1L1 protein binds cholesterol and plays essential roles in cholesterol uptake. J. Biol. Chem. 2011, 286, 25088–25097. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Yang, J.; Chang, Y.; Qiao, L.; Dang, H.; Luo, K.; Guo, H.; An, Y.; Ma, C.; Shao, H.; et al. Caffeine-free hawk tea lowers cholesterol by reducing free cholesterol uptake and the production of very-low-density lipoprotein. Commun. Biol. 2019, 2, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, T.; Sakurada, T.; Watanabe, R.; Yamaguchi, K.; Kimura, Y.; Kioka, N.; Kawagishi, H.; Matsuo, M.; Ueda, K. Fomiroid A, a novel compound from the mushroom Fomitopsis nigra, inhibits NPC1L1-mediated cholesterol uptake via a mode of action distinct from that of ezetimibe. PLoS ONE 2014, 9, e116162. [Google Scholar] [CrossRef] [Green Version]
- Nekohashi, M.; Ogawa, M.; Ogihara, T.; Nakazawa, K.; Kato, H.; Misaka, T.; Abe, K.; Kobayashi, S. Luteolin and quercetin affect the cholesterol absorption mediated by epithelial cholesterol transporter niemann-pick c1-like 1 in caco-2 cells and rats. PLoS ONE 2014, 9, e97901. [Google Scholar] [CrossRef] [Green Version]
- Na, A.Y.; Yang, E.J.; Jeon, J.M.; Ki, S.H.; Song, K.S.; Lee, S. Protective Effect of Isoliquiritigenin against Ethanol-Induced Hepatic Steatosis by Regulating the SIRT1-AMPK Pathway. Toxicol. Res. 2018, 34, 23–29. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, J.; Liu, W.; Liang, B.; Shi, L.; Yang, S.; Meng, J.; Chang, J.; Hu, X.; Zhang, R.; Xing, D. Inhibitory Effect of Isoliquiritigenin in Niemann-Pick C1-Like 1-Mediated Cholesterol Uptake. Molecules 2022, 27, 7494. https://doi.org/10.3390/molecules27217494
Zeng J, Liu W, Liang B, Shi L, Yang S, Meng J, Chang J, Hu X, Zhang R, Xing D. Inhibitory Effect of Isoliquiritigenin in Niemann-Pick C1-Like 1-Mediated Cholesterol Uptake. Molecules. 2022; 27(21):7494. https://doi.org/10.3390/molecules27217494
Chicago/Turabian StyleZeng, Jun, Wenjing Liu, Bing Liang, Lingyu Shi, Shanbo Yang, Jingsen Meng, Jing Chang, Xiaokun Hu, Renshuai Zhang, and Dongming Xing. 2022. "Inhibitory Effect of Isoliquiritigenin in Niemann-Pick C1-Like 1-Mediated Cholesterol Uptake" Molecules 27, no. 21: 7494. https://doi.org/10.3390/molecules27217494
APA StyleZeng, J., Liu, W., Liang, B., Shi, L., Yang, S., Meng, J., Chang, J., Hu, X., Zhang, R., & Xing, D. (2022). Inhibitory Effect of Isoliquiritigenin in Niemann-Pick C1-Like 1-Mediated Cholesterol Uptake. Molecules, 27(21), 7494. https://doi.org/10.3390/molecules27217494