
In Silico Study and Effects of BDMC33 on TNBS-Induced BMP Gene Expressions in Zebrafish Gut Inflammation-Associated Arthritis

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
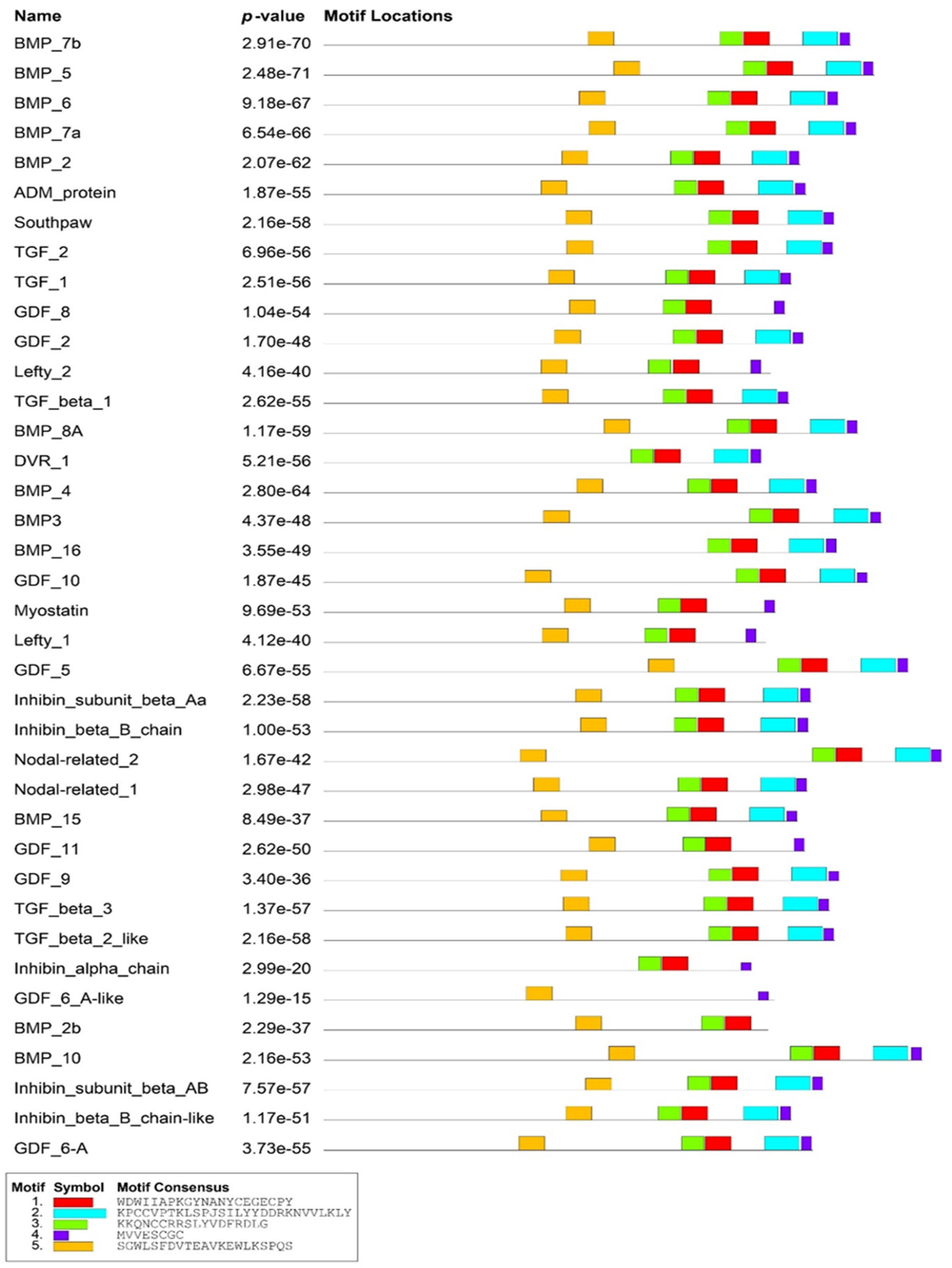
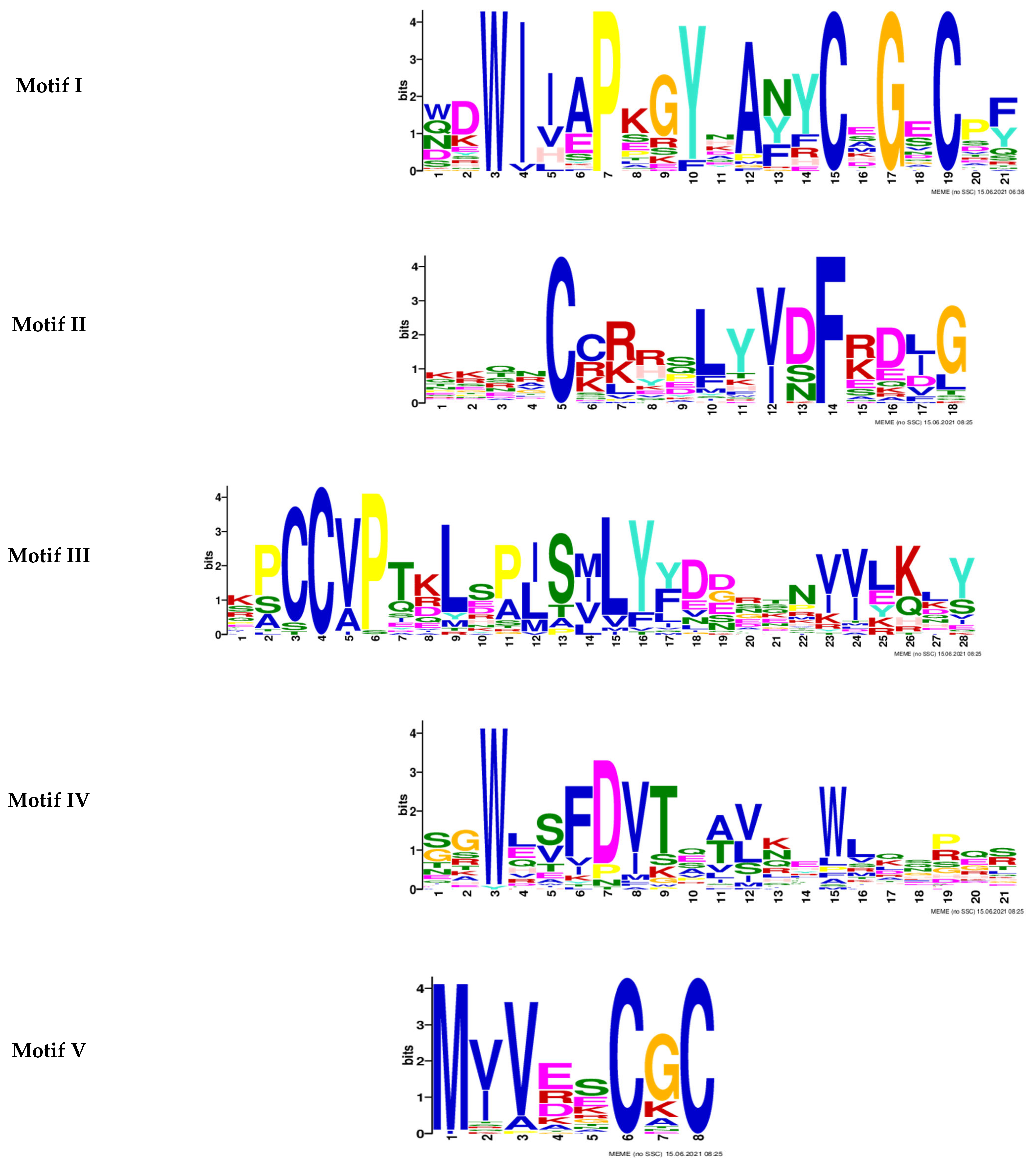
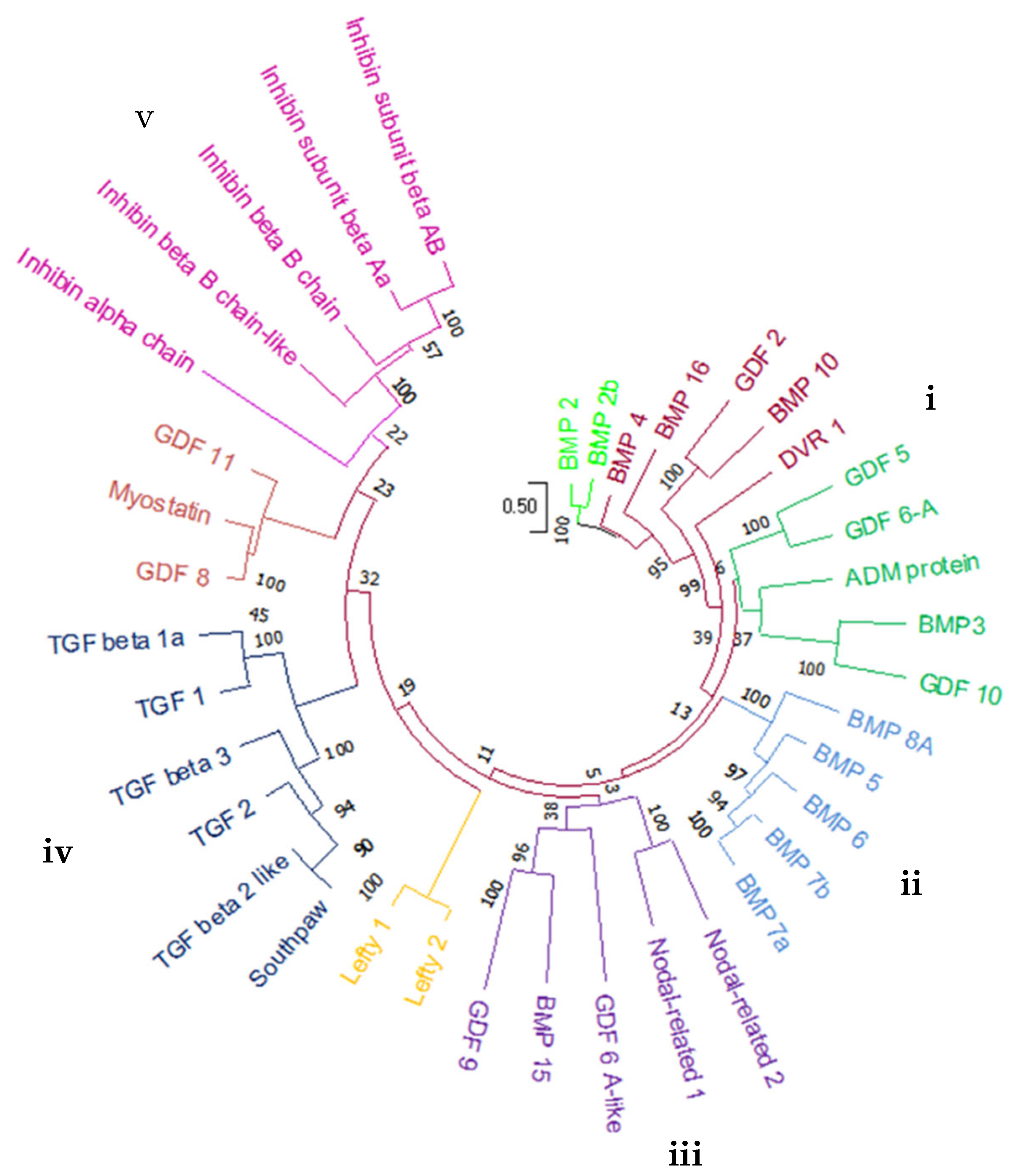
2.1. Genome-Wide Analysis of BMP Family in Zebrafish
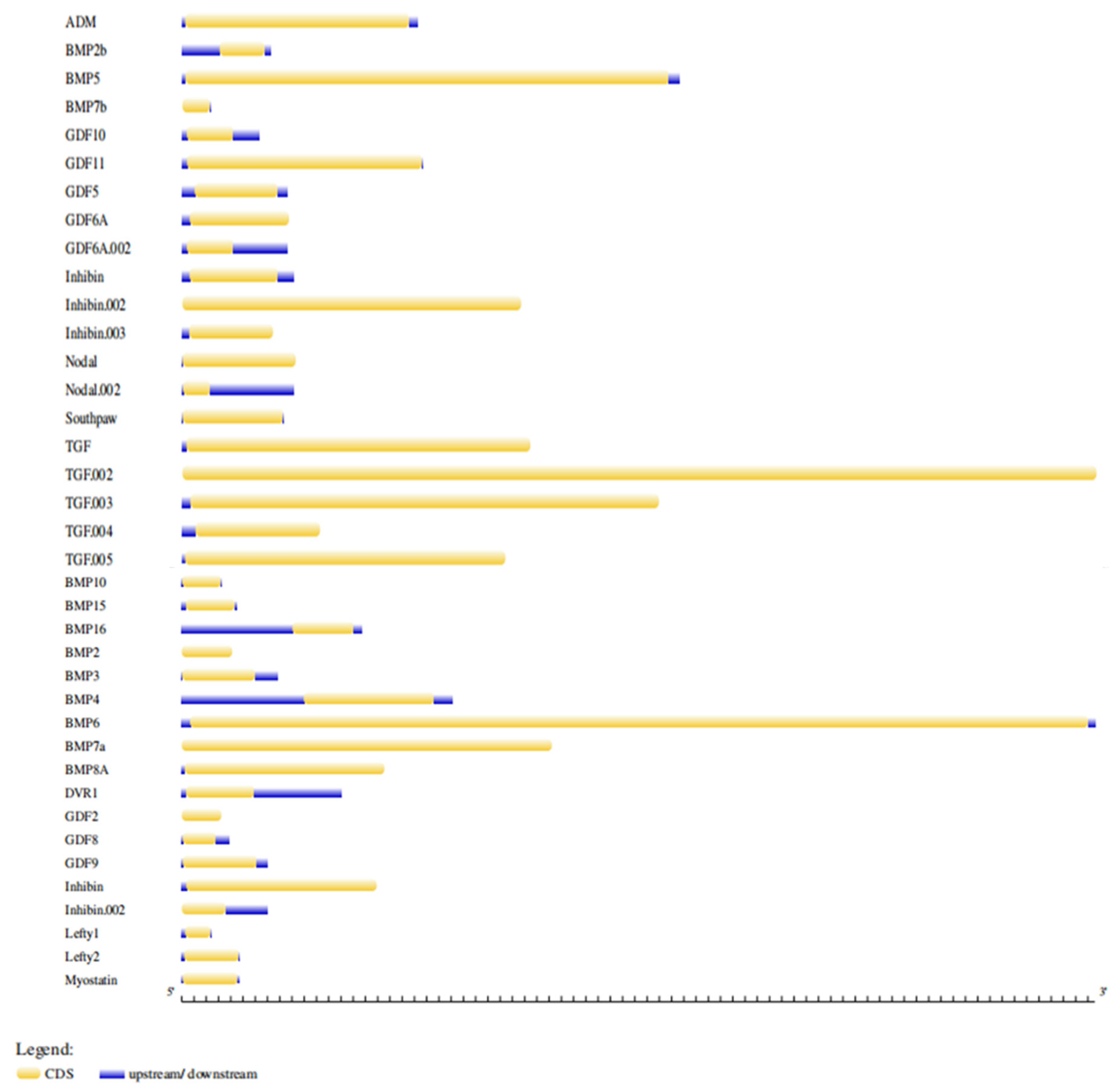
2.2. Gene Structure Analysis of BMP Family in Zebrafish Genome
2.3. Physicochemical Properties of BMP Family of Zebrafish Genome
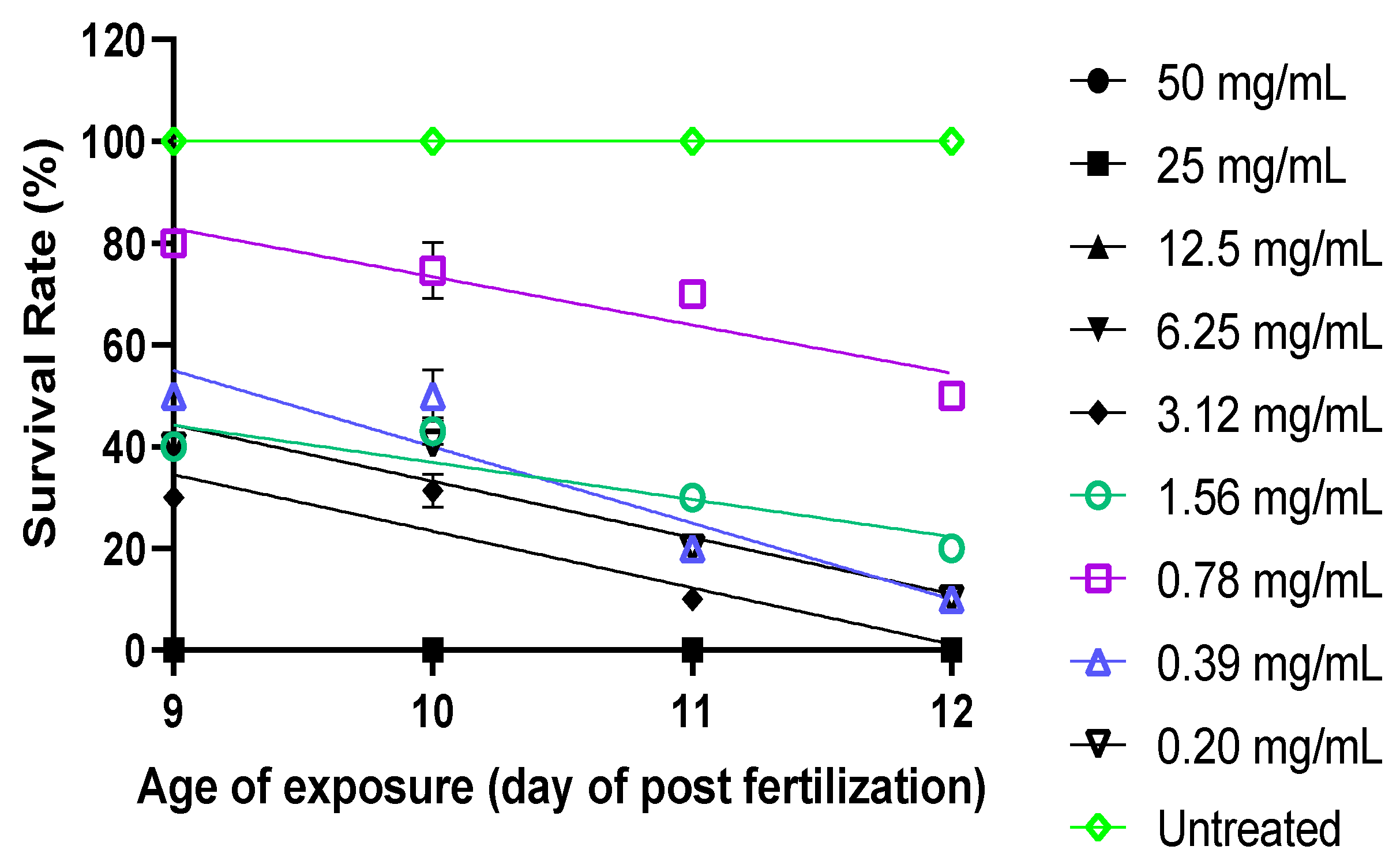
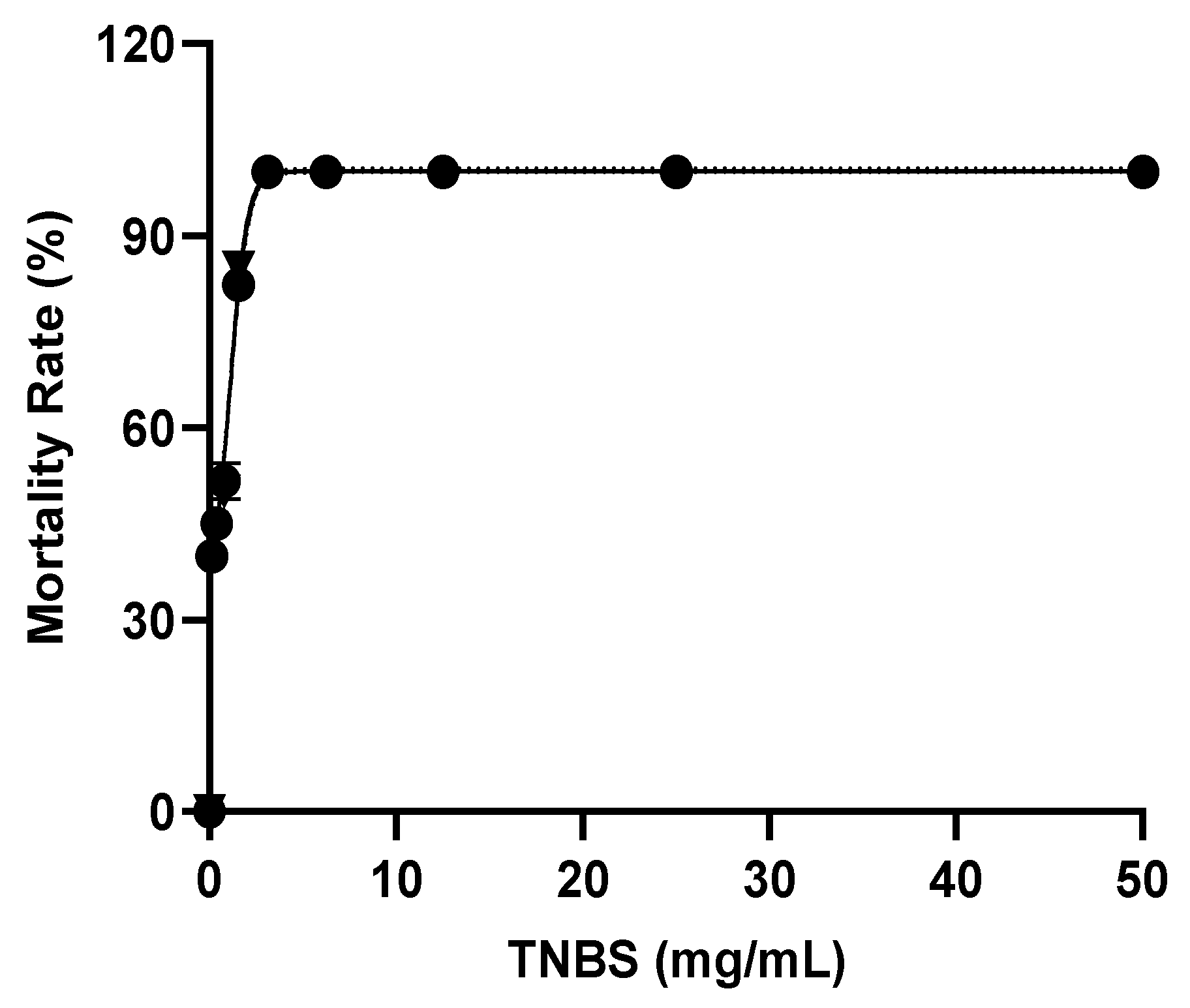
2.4. Optimization of TNBS Concentration
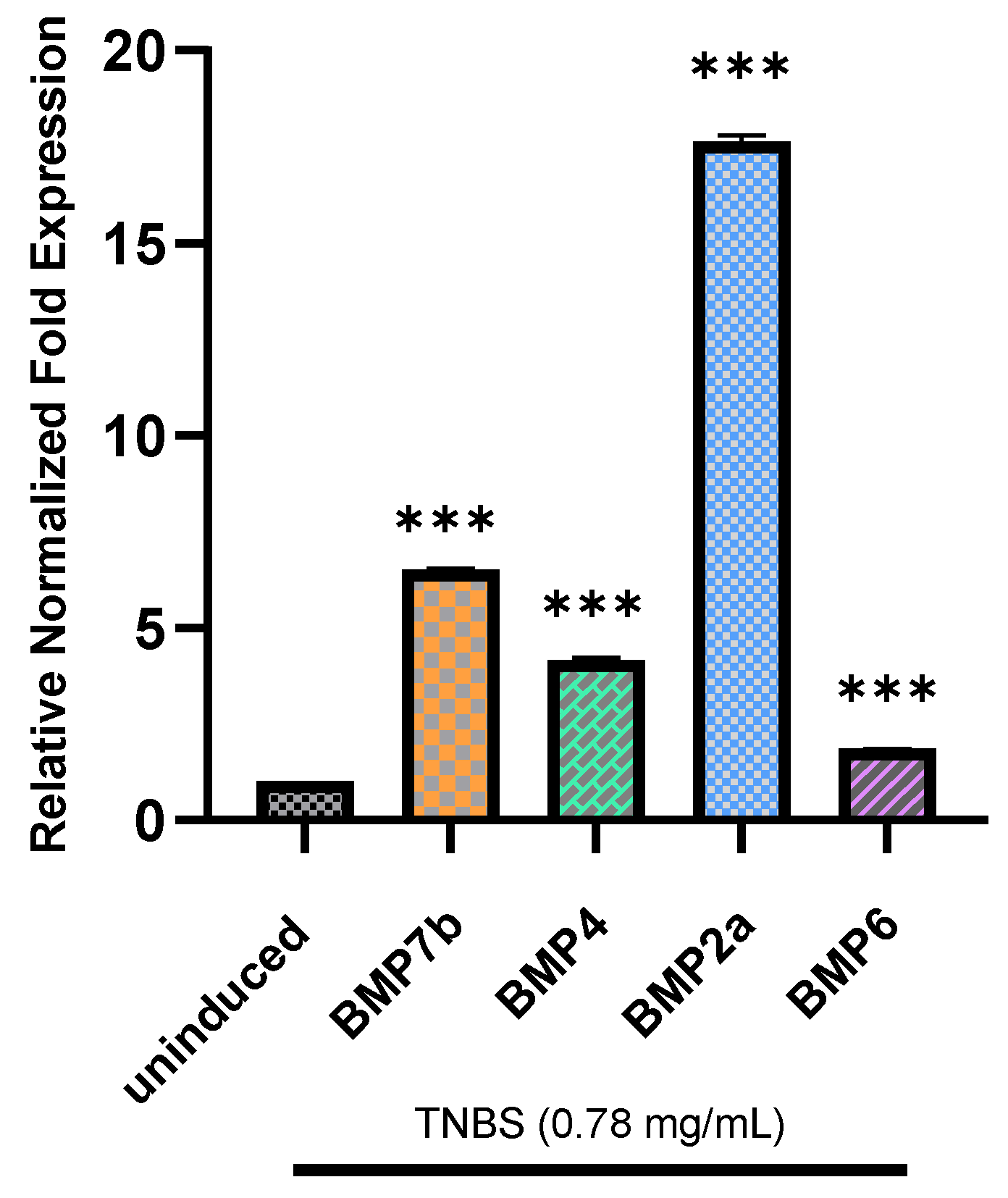
2.5. Relative Normalized Fold Expression of BMP7b, BMP4, BMP2a, and BMP6
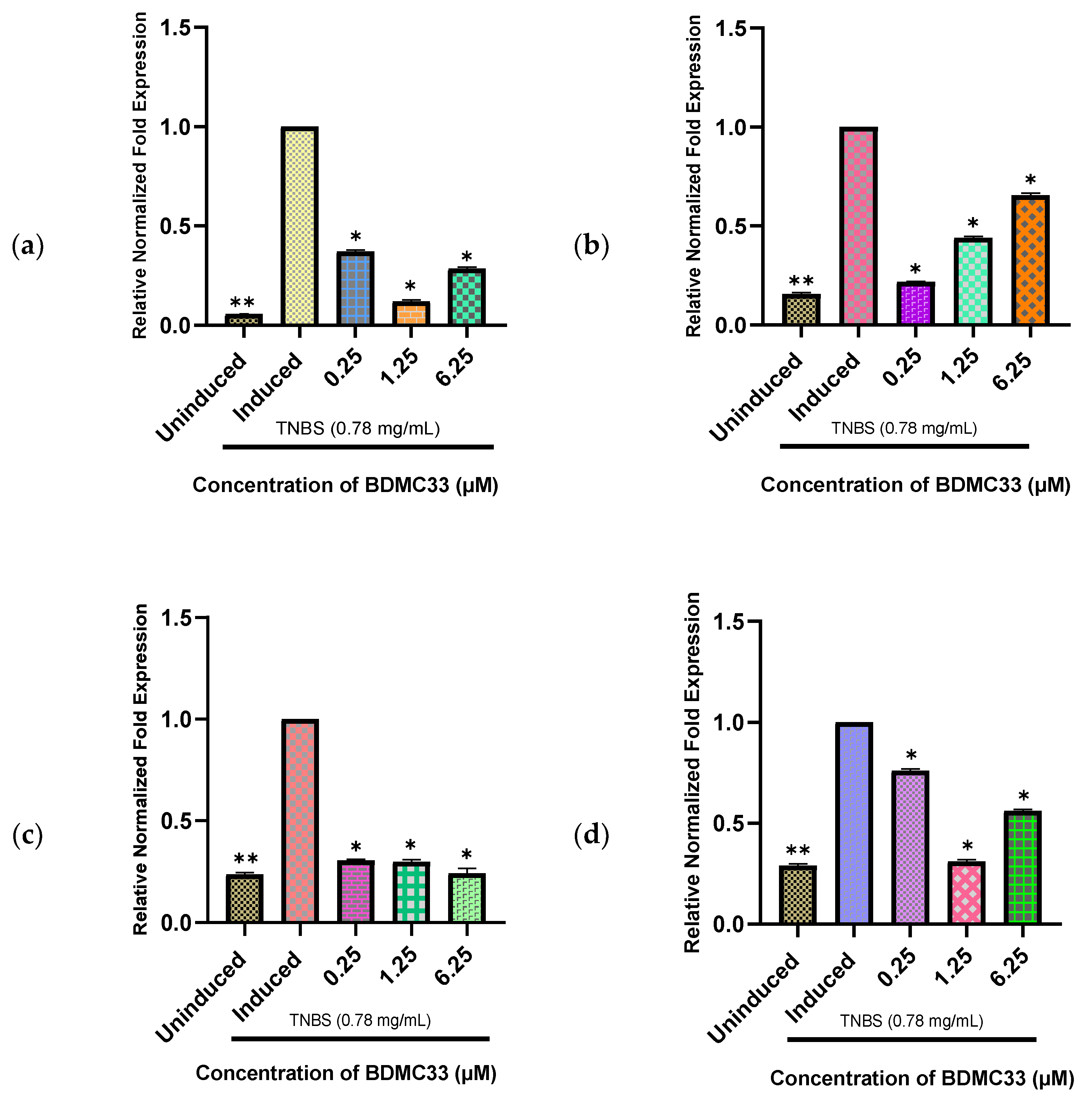
2.6. BDMC33 Suppressed Gene Expression of BMP2a, BMP4, BMP6, and BMP7b in Inflammatory State
3. Discussion
4. Materials and Methods
4.1. Animal Care and Husbandry
4.2. Identification of BMP Family
4.3. Sequence Analyses of Zebrafish BMP Family Members
4.4. Optimization of TNBS Exposure to Larvae
4.5. BDMC33 Preparation
4.6. Total RNA Extraction and cDNA Synthesis
4.7. Gene Expression Analysis
4.8. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP Signaling and Other Molecular Events: Regulation of Osteoblastogenesis and Bone Formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, L.M.; Miller, C.; Kuczma, M.; Pietrzak, M.; Jing, Y.; Rempala, G.; Muranski, P.; Ignatowicz, L.; Kraj, P. Bone Morphogenic Proteins Are Immunoregulatory Cytokines Controlling FOXP3+ Treg Cells. Cell Rep. 2020, 33, 108219. [Google Scholar] [CrossRef] [PubMed]
- Aluganti Narasimhulu, C.; Singla, D.K. The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases. Cells 2020, 9, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP Signalling in Skeletal Development, Disease and Repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef]
- Maric, I.; Turk, T.; Smoljan, I.; Orlic, Z.; Bobinac, D. Bone Morphogenetic Proteins and Signaling Pathway in Inflammatory Bowel Disease. AJP-Gastrointest. Liver Physiol. 2012, 15, G1151–G1162. [Google Scholar] [CrossRef] [Green Version]
- Fleming, A.; Jankowski, J.; Goldsmith, P. In Vivo Analysis of Gut Function and Disease Changes in a Zebrafish Larvae Model of Inflammatory Bowel Disease: A Feasibility Study. Inflamm. Bowel Dis. 2010, 16, 1162–1172. [Google Scholar] [CrossRef]
- Wakim, S.; Grewal, M. Disorders of the Skeletal System. Available online: https://bio.libretexts.org/Bookshelves/Human_Biology/Book%3A_Human_Biology_(Wakim_and_Grewal)/14%3A_Skeletal_System/14.7%3A_Disorders_of_the_Skeletal_System (accessed on 15 March 2022).
- Mortier, C.; Govindarajan, S.; Venken, K.; Elewaut, D. It Takes “Guts” to Cause Joint Inflammation: Role of Innate-Like T Cells. Front. Immunol. 2018, 9, 1489. [Google Scholar] [CrossRef]
- Ashrafi, M.; Kuhn, K.A.; Weisman, M.H. The Arthritis Connection to Inflammatory Bowel Disease (IBD): Why Has It Taken So Long to Understand It? RMD Open 2021, 7, e001558. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Yao, Y. Biology of Interleukin-17 and Its Pathophysiological Significance in Sepsis. Front. Immunol. 2020, 11, 1558. [Google Scholar] [CrossRef]
- Briolay, A.; El Jamal, A.; Arnolfo, P.; Le Goff, B.; Blanchard, F.; Magne, D.; Bougault, C. Enhanced BMP-2/BMP-4 Ratio in Patients with Peripheral Spondyloarthritis and in Cytokine- and Stretch-Stimulated Mouse Chondrocytes. Arthritis Res. Ther. 2020, 22, 234. [Google Scholar] [CrossRef] [PubMed]
- Maric, I.; Kucic, N.; Turk Wensveen, T.; Smoljan, I.; Grahovac, B.; Zoricic Cvek, S.; Celic, T.; Bobinac, D.; Vukicevic, S. BMP Signaling in Rats with TNBS-Induced Colitis Following BMP7 Therapy. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1151–G1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Puerto, M.C.; Iyengar, P.V.; García de Vinuesa, A.; Ten Dijke, P.; Sanchez-Duffhues, G. Bone Morphogenetic Protein Receptor Signal Transduction in Human Disease. J. Pathol. 2019, 247, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhu, K.; Guo, H.; Zhang, N.; Jiang, S.; Zhang, D. Genome-Wide Comparative Analysis Ofbone Morphogenetic Proteins: Genomic Structure, Phylogeny, and Expression Patterns in the Golden Pompano, Trachinotus Ovatus(Linnaeus, 1758). Fish Physiol. Biochem. 2020, 46, 345–358. [Google Scholar] [CrossRef]
- Cheng, J.; Duan, X.; Fu, X.; Jiang, Y.; Yang, P.; Cao, C.; Li, Q.; Zhang, J.; Hu, X.; Zhang, X.; et al. RIP1 Perturbation Induces Chondrocyte Necroptosis and Promotes Osteoarthritis Pathogenesis via Targeting BMP7. Front. Cell Dev. Biol. 2021, 9, 638382. [Google Scholar] [CrossRef]
- Park, S.C.; Jeen, Y.T. Genetic Studies of Inflammatory Bowel Disease-Focusing on Asian Patients. Cells 2019, 8, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oehlers, S.H.; Flores, M.V.; Okuda, K.S.; Hall, C.J.; Crosier, K.E.; Crosier, P.S. A Chemical Enterocolitis Model in Zebrafish Larvae That Is Dependent on Microbiota and Responsive to Pharmacological Agents. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2011, 240, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Meijer, A.H.; Schaaf, M.J.M. Modeling Inflammation in Zebrafish for the Development of Anti-Inflammatory Drugs. Front. Cell Dev. Biol. 2020, 8, 620984. [Google Scholar] [CrossRef]
- Maric, I.; Smoljan, I.; Turk, T.; Celic, T.; Zoricic, S.; Orlic, Z.; Bobinac, D. BMP Signaling Pathway in Experimental Inflammatory Bowel Disease. Bone 2011, 48, S238. [Google Scholar] [CrossRef]
- Lu, P.-D.; Zhao, Y.-H. Targeting NF-ΚB Pathway for Treating Ulcerative Colitis: Comprehensive Regulatory Characteristics of Chinese Medicines. Chin. Med. 2020, 15, 1–15. [Google Scholar] [CrossRef]
- Ferrero, E.; Dunham, I.; Sanseau, P. In Silico Prediction of Novel Therapeutic Targets Using Gene–Disease Association Data. J. Transl. Med. 2017, 15, 182. [Google Scholar] [CrossRef] [Green Version]
- Teame, T.; Zhang, Z.; Ran, C.; Zhang, H.; Yang, Y.; Ding, Q.; Xie, M.; Gao, C.; Ye, Y.; Duan, M.; et al. The Use of Zebrafish (Danio Rerio) as Biomedical Models. Anim. Front. 2019, 9, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanyang, L.; Xuanzhe, L.; Xuyang, C.; Yujia, Q.; Jiarong, F.; Jun, S.; Zhihua, R. Application of Zebrafish Models in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 501. [Google Scholar] [CrossRef] [Green Version]
- Belo, M.A.A.; Oliveira, M.F.; Oliveira, S.L.; Aracati, M.F.; Rodrigues, L.F.; Costa, C.C.; Conde, G.; Gomes, J.M.M.; Prata, M.N.L.; Barra, A.; et al. Zebrafish as a Model to Study Inflammation: A Tool for Drug Discovery. Biomed. Pharmacother. 2021, 144, 112310. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.Z.; Benslimane, F.M.; Nasrallah, G.K.; Shurbaji, S.; Younes, N.N.; Mraiche, F.; Da’as, S.I.; Yalcin, H.C. Using Zebrafish for Investigating the Molecular Mechanisms of Drug-Induced Cardiotoxicity. Biomed Res. Int. 2018, 2018, 1642684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Li, J.; Lu, F.; Zhang, J.; Guo, L. Application of Zebrafish in the Study of the Gut Microbiome. Anim. Model. Exp. Med. 2022, 5, 323–336. [Google Scholar] [CrossRef]
- Prattis, S.; Jurjus, A. Spontaneous and Transgenic Rodent Models of Inflammatory Bowel Disease. Lab. Anim. Res. 2015, 31, 47–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [Green Version]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the Golden Nutraceutical: Multitargeting for Multiple Chronic Diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.-H.; Loo, C.-Y.; Bebawy, M.; Luk, F.; Mason, R.S.; Rohanizadeh, R. Curcumin and Its Derivatives: Their Application in Neuropharmacology and Neuroscience in the 21st Century. Curr. Neuropharmacol. 2013, 11, 338–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ka Heng, L.; Aziz, F.; Ahmad, S.; Abas, F.; Israf, D.; Nordin, N.H. Synthesis and Biological Evaluation of Curcumin-like Diarylpentanoid Analogues for Anti-Inflammatory, Antioxidant and Anti-Tyrosinase Activities. Eur. J. Med. Chem. 2009, 44, 3195–3200. [Google Scholar] [CrossRef]
- Yakubu, N.; Ahmad, S.; Usman, H.; Abubakar, H.; Amuzat, A. Acute and Chronic Toxicity Effects Of 2, 6-Bis (2, 5-Dimethoxybenzy-Lidene) Cyclohexanone (BDMC33) In Zebrafish (Danio rerio) Tested on Adulthood Life Stage. IOSR J. Pharm. Biol. Sci. 2018, 8, 18–27. [Google Scholar]
- Razali, N.A.; Nazarudin, N.A.; Lai, K.S.; Abas, F.; Ahmad, S. Curcumin Derivative, 2,6-Bis(2-Fluorobenzylidene) Cyclohexanone (MS65) Inhibits Interleukin-6 Production through Suppression of NF-ΚB and MAPK Pathways in Histamine-Induced Human Keratinocytes Cell (HaCaT). BMC Complement. Altern. Med. 2018, 18, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zha, Z.; Shen, W.; Li, D.; Kang, N.; Chen, Z.; Liu, Y.; Xu, G.; Xu, Q. Anemoside B4 Ameliorates TNBS-Induced Colitis through S100A9/MAPK/NF-ΚB Signaling Pathway. Chin. Med. 2021, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a Promising Target for the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Lee, K.-H.; Abas, F.; Mohamed Alitheen, N.B.; Shaari, K.; Lajis, N.H.; Israf, D.A.; Syahida, A. Chemopreventive Effects of a Curcumin-like Diarylpentanoid [2,6-Bis(2,5-Dimethoxybenzylidene) Cyclohexanone] in Cellular Targets of Rheumatoid Arthritis in Vitro. Int. J. Rheum. Dis. 2015, 18, 616–627. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP Signaling in Osteoblast, Skeletal Development, and Bone Formation, Homeostasis and Disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Que, J. BMP Signaling in Development, Stem Cells, and Diseases of the Gastrointestinal Tract. Annu. Rev. Physiol. 2020, 82, 251–273. [Google Scholar] [CrossRef] [PubMed]
- Dick, A.; Hild, M.; Bauer, H.; Imai, Y.; Maifeld, H.; Schier, A.F.; Talbot, W.S.; Bouwmeester, T.; Hammerschmidt, M. Essential Role of Bmp7 (Snailhouse) and Its Prodomain in Dorsoventral Patterning of the Zebrafish Embryo. Development 2000, 127, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Honsawek, S.; Chayanupatkul, M.; Tanavalee, A.; Sakdinakiattikoon, M.; Deepaisarnsakul, B.; Yuktanandana, P.; Ngarmukos, S. Relationship of Plasma and Synovial Fluid BMP-7 with Disease Severity in Knee Osteoarthritis Patients: A Pilot Study. Int. Orthop. 2009, 33, 1171–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fioruci-Fontanelli, B.A.; Chuffa, L.G.A.; Mendes, L.O.; Pinheiro, P.F.F.; Delella, F.K.; Kurokawa, C.S.; Felisbino, S.L.; Martinez, F.E. MMP-2 and MMP-9 Activities and TIMP-1 and TIMP-2 Expression in the Prostatic Tissue of Two Ethanol-Preferring Rat Models. Anal. Cell. Pathol. 2015, 2015, 954548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, S.; Jakaria, M.; Kim, I.-S.; Kim, J.; Haque, M.E.; Choi, D.-K. Regulation of Toll-Like Receptor (TLR) Signaling Pathway by Polyphenols in the Treatment of Age-Linked Neurodegenerative Diseases: Focus on TLR4 Signaling. Front. Immunol. 2019, 10, 1000. [Google Scholar] [CrossRef]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) Signaling in Development and Human Diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.-L.; Yuan, Y.; Zou, G.-M.; Liu, G.; Tu, J.; Li, Q. LPS-Stimulated Inflammatory Environment Inhibits BMP-2-Induced Osteoblastic Differentiation through Crosstalk between TLR4/MyD88/NF-ΚB and BMP/Smad Signaling. Stem Cells Dev. 2014, 23, 277–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-H.; Abas, F.; Alitheen, N.B.M.; Shaari, K.; Lajis, N.H.; Ahmad, S. A Curcumin Derivative, 2,6-Bis(2,5-Dimethoxybenzylidene)-Cyclohexanone (BDMC33) Attenuates Prostaglandin E2 Synthesis via Selective Suppression of Cyclooxygenase-2 in IFN-γ/LPS-Stimulated Macrophages. Molecules 2011, 16, 9728–9738. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, K.I. The Chemistry of Curcumin: From Extraction to Therapeutic Agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad Jafari, R.; Shayesteh, S.; Ala, M.; Yousefi-Manesh, H.; Rashidian, A.; Hashemian, S.M.; Sorouri, M.; Dehpour, A.R. Dapsone Ameliorates Colitis through TLR4/NF-KB Pathway in TNBS Induced Colitis Model in Rat. Arch. Med. Res. 2021, 52, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-H.; Chow, Y.-L.; Sharmili, V.; Abas, F.; Alitheen, N.B.M.; Shaari, K.; Israf, D.A.; Lajis, N.H.; Syahida, A. BDMC33, A Curcumin Derivative Suppresses Inflammatory Responses in Macrophage-like Cellular System: Role of Inhibition in NF-ΚB and MAPK Signaling Pathways. Int. J. Mol. Sci. 2012, 13, 2985–3008. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information (NCBI). Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 12 January 2022).
- European Molecular Biology Laboratory-European Bioinformatic Institiute, Hinxton, Cambridgeshire, UK. Available online: http://pfam.xfam.org (accessed on 12 January 2022).
- The European Molecular Biology Laboratory-European Bioinformatic Institiute, Hinxton, Cambridgeshire, UK. Available online: http://hmmer.org (accessed on 12 January 2022).
- National Center for Biotechnology Information (NCBI). Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml/ (accessed on 12 January 2022).
- National Center for Biotechnology Information (NCBI). Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins/ (accessed on 13 January 2022).
- Auclair, S.; Rossetti, R.; Meslin, C.; Monestier, O.; Pasquale, E.; Pascal, G.; Persani, L.; Fabre, S. Positive Selection in Bone Morphogenetic Protein 15 Targets a Natural Mutation Associated with Primary Ovarian Insufficiency in Human. PLoS ONE 2013, 8, e78199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information (NCBI). Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/tools/primer-blast/ (accessed on 10 March 2022).
- Integarted DNA Technology, USA. Available online: https://sg.idtdna.com/pages/tools/oligoanalyzer (accessed on 15 March 2022).
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Properties | Values |
|---|---|
| Molecular weights | 40,201.62–54,217.72 Da |
| Isoelectric point (pI) | 6.13–9.85 |
| Total number of negatively charged values Asp + G/Glu | 1883 |
| Total number of positively charged residues Arg + Lys | 1605 |
| Protein Length | 347–501 AA |
| Instability Index | 41.07–81.74 |
| Aliphatic Index | 71.07–90.28 |
| GRAVY | −0.225–(−0.786) |
| Accession No | Gene Name | Forward Primer | Reverse Primer | Tm |
|---|---|---|---|---|
| NM_131359.1 | BMP2a | 5′-TTCGCTAAACAACGCAAGTG-3′ | 5′-GAACAAGCCTGGTGTCCAAT-3′ | 58.3 °C |
| NM_131342.2 | BMP4 | 5′-TGAGGCACAACACCTCCAAA-3′ | 5′-ACTTTTGCCGTCATGTCCGA-3′ | 61 °C |
| NM_001013339.1 | BMP6 | 5′ACCTGTTTCTGCTGGAATCTC3′ | 5′CCCTGCATCCTTTGGACTG3′ | 56.9 °C |
| NM_131321.2 | BMP7b | 5′-TTGACCTTTCTCGCATCCCG-3′ | 5′-GCAGGTACAGCTCCCTTCTG-3′ | 61 °C |
| NM_001115114.1 | GAPDH | 5′-CATCTTTGACGCTGGTGCTG-3′ | 5′-TGGGAGAATGGTCGCGTATC-3′ | 59.3 °C |
| NM_131031.2 | β Actin | 5′-TACCACTTTGCCCTCCTCAC-3′ | 5′-GACACCCTGGCTTACATTTTCA-3′ | 58.6 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostofa, F.; Yasid, N.A.; Shamsi, S.; Ahmad, S.A.; Mohd-Yusoff, N.F.; Abas, F.; Ahmad, S. In Silico Study and Effects of BDMC33 on TNBS-Induced BMP Gene Expressions in Zebrafish Gut Inflammation-Associated Arthritis. Molecules 2022, 27, 8304. https://doi.org/10.3390/molecules27238304
Mostofa F, Yasid NA, Shamsi S, Ahmad SA, Mohd-Yusoff NF, Abas F, Ahmad S. In Silico Study and Effects of BDMC33 on TNBS-Induced BMP Gene Expressions in Zebrafish Gut Inflammation-Associated Arthritis. Molecules. 2022; 27(23):8304. https://doi.org/10.3390/molecules27238304
Chicago/Turabian StyleMostofa, Farhana, Nur Adeela Yasid, Suhaili Shamsi, Siti Aqlima Ahmad, Nur Fatihah Mohd-Yusoff, Faridah Abas, and Syahida Ahmad. 2022. "In Silico Study and Effects of BDMC33 on TNBS-Induced BMP Gene Expressions in Zebrafish Gut Inflammation-Associated Arthritis" Molecules 27, no. 23: 8304. https://doi.org/10.3390/molecules27238304
APA StyleMostofa, F., Yasid, N. A., Shamsi, S., Ahmad, S. A., Mohd-Yusoff, N. F., Abas, F., & Ahmad, S. (2022). In Silico Study and Effects of BDMC33 on TNBS-Induced BMP Gene Expressions in Zebrafish Gut Inflammation-Associated Arthritis. Molecules, 27(23), 8304. https://doi.org/10.3390/molecules27238304