


Achillea moschata Wulfen: From Ethnobotany to Phytochemistry, Morphology, and Biological Activity

 , , , , , , ,
, , , , , , ,  , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Ethnobotanical Research
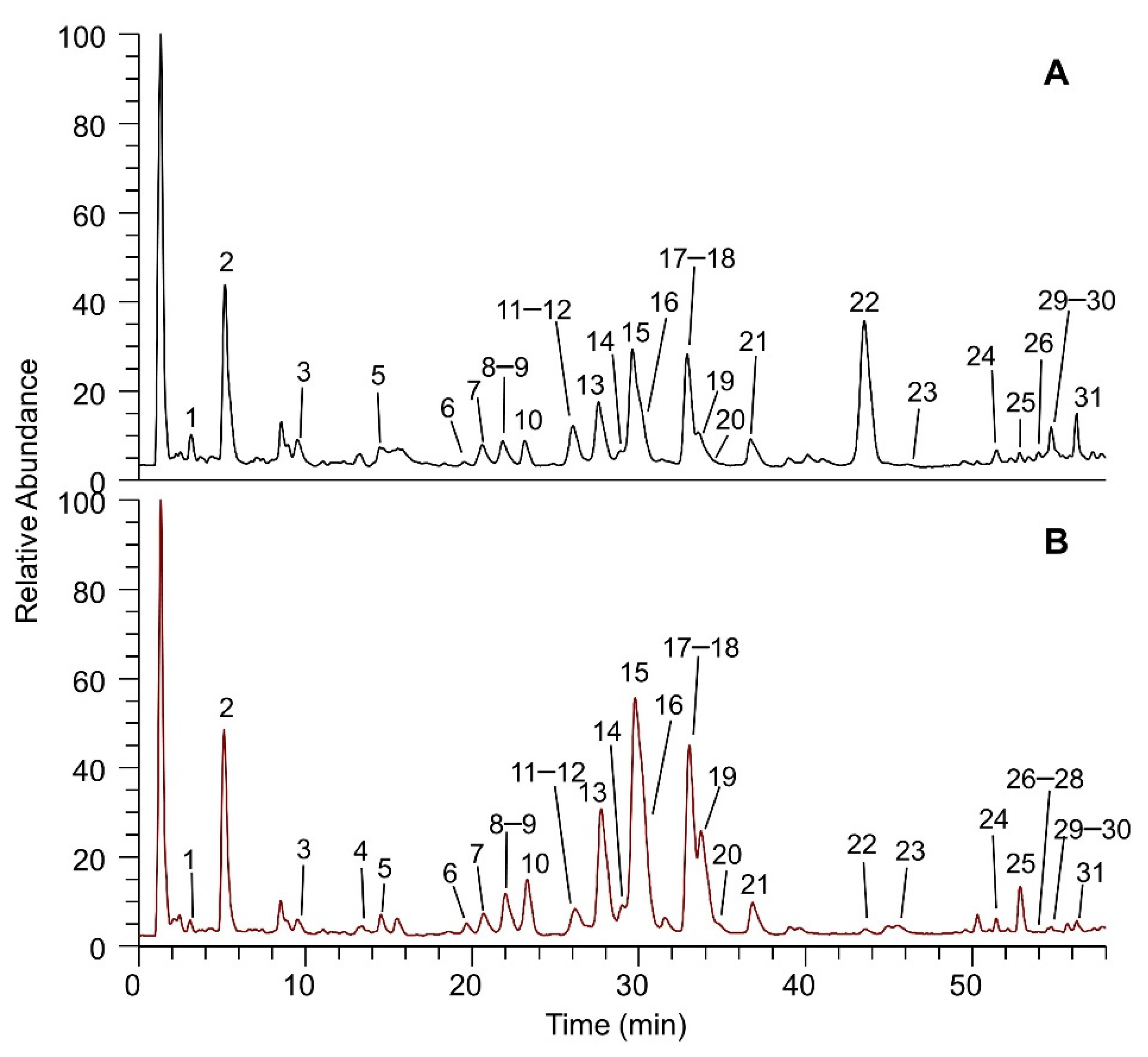
2.2. Phytochemical Investigation
2.3. Morphological Investigation
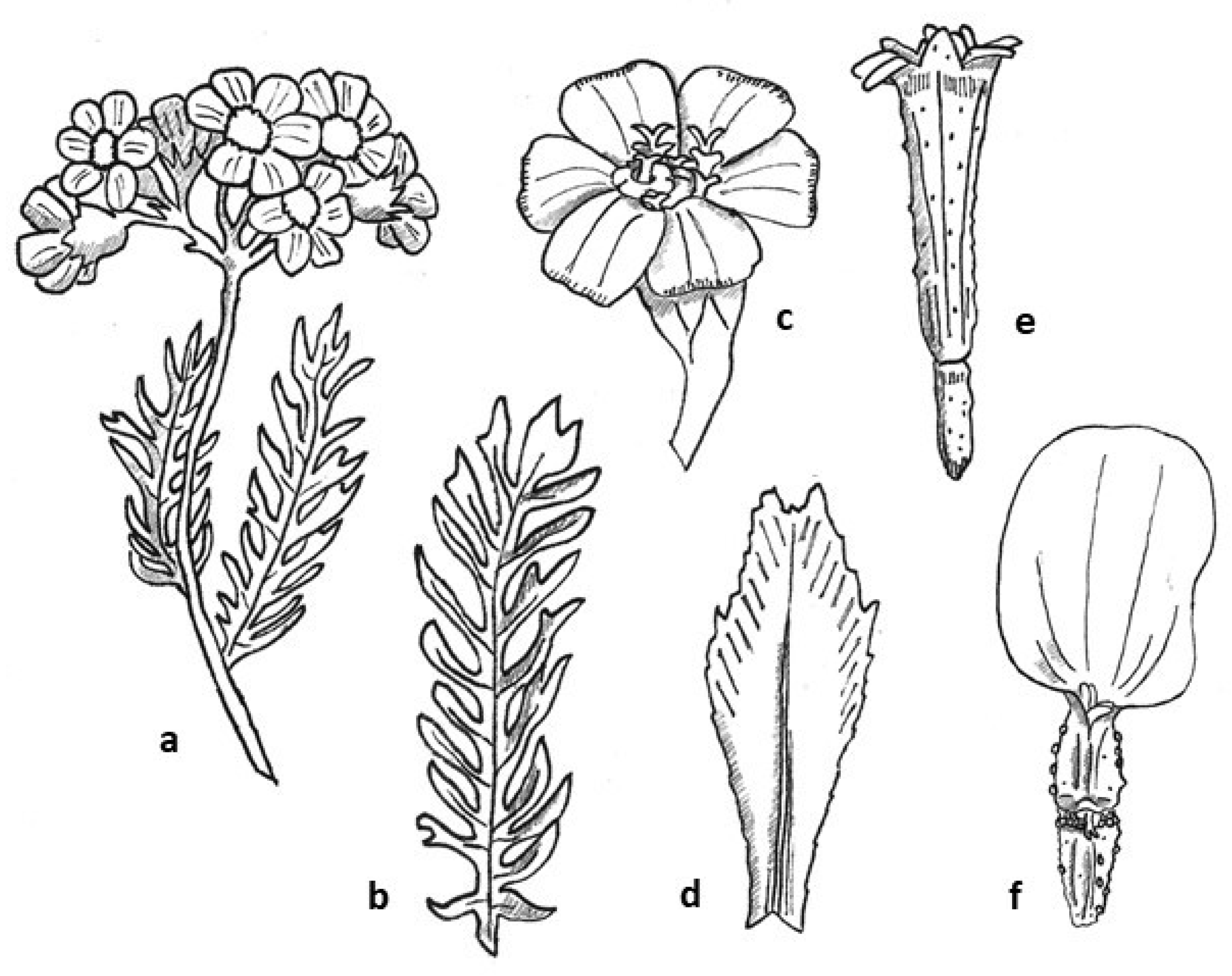
2.3.1. Botanical Description
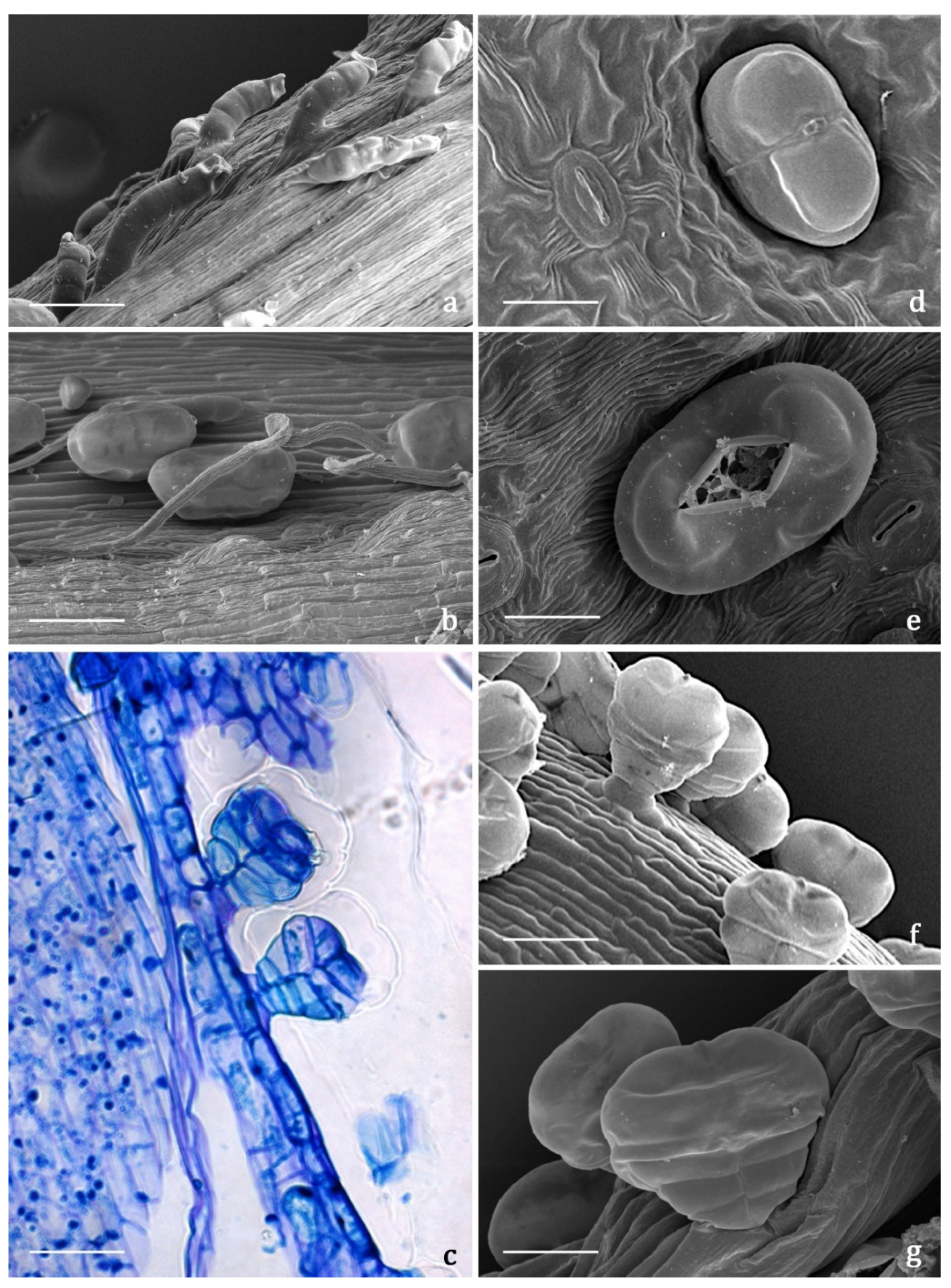
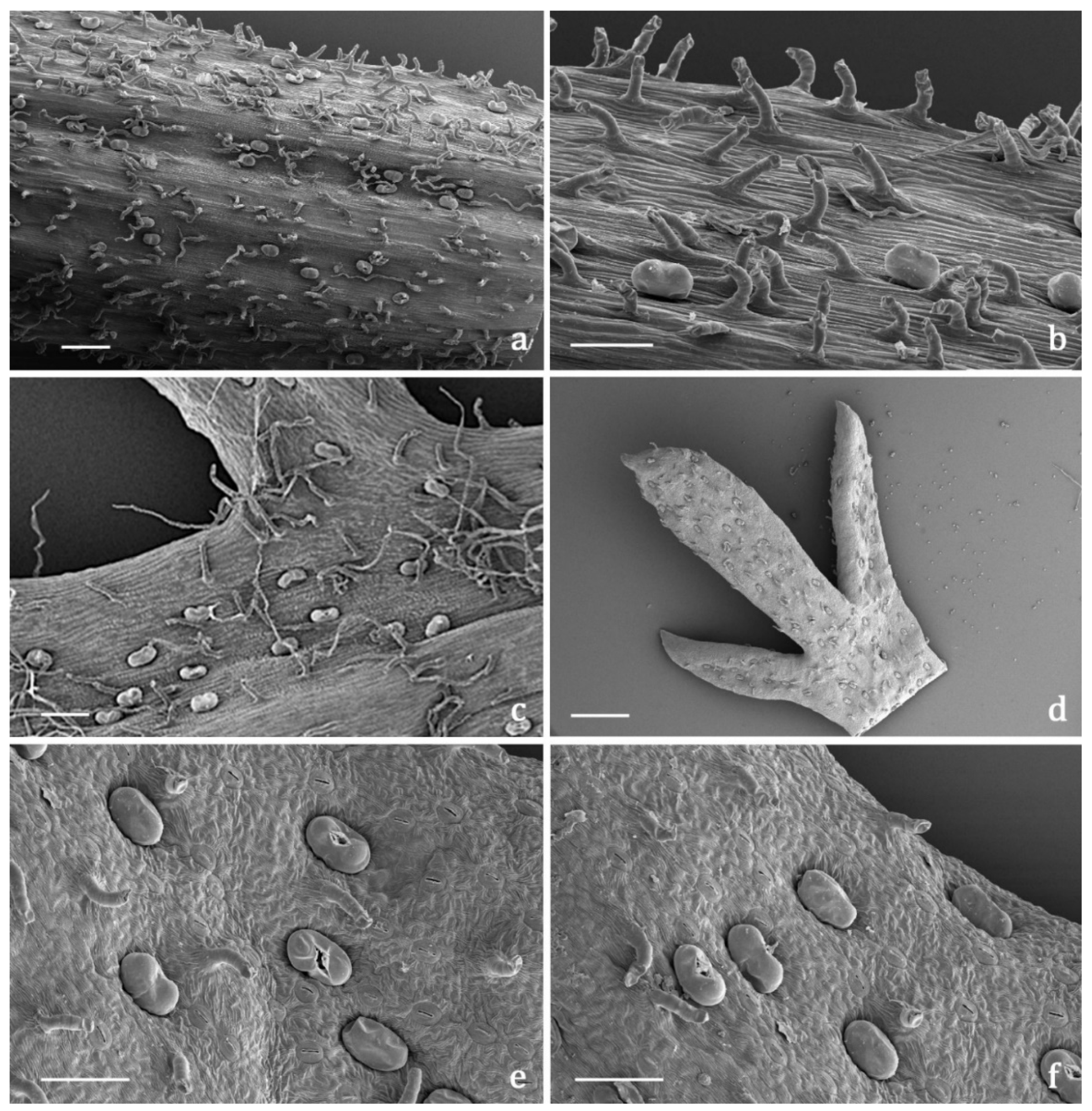
2.3.2. Indumentum
2.3.3. Histochemistry
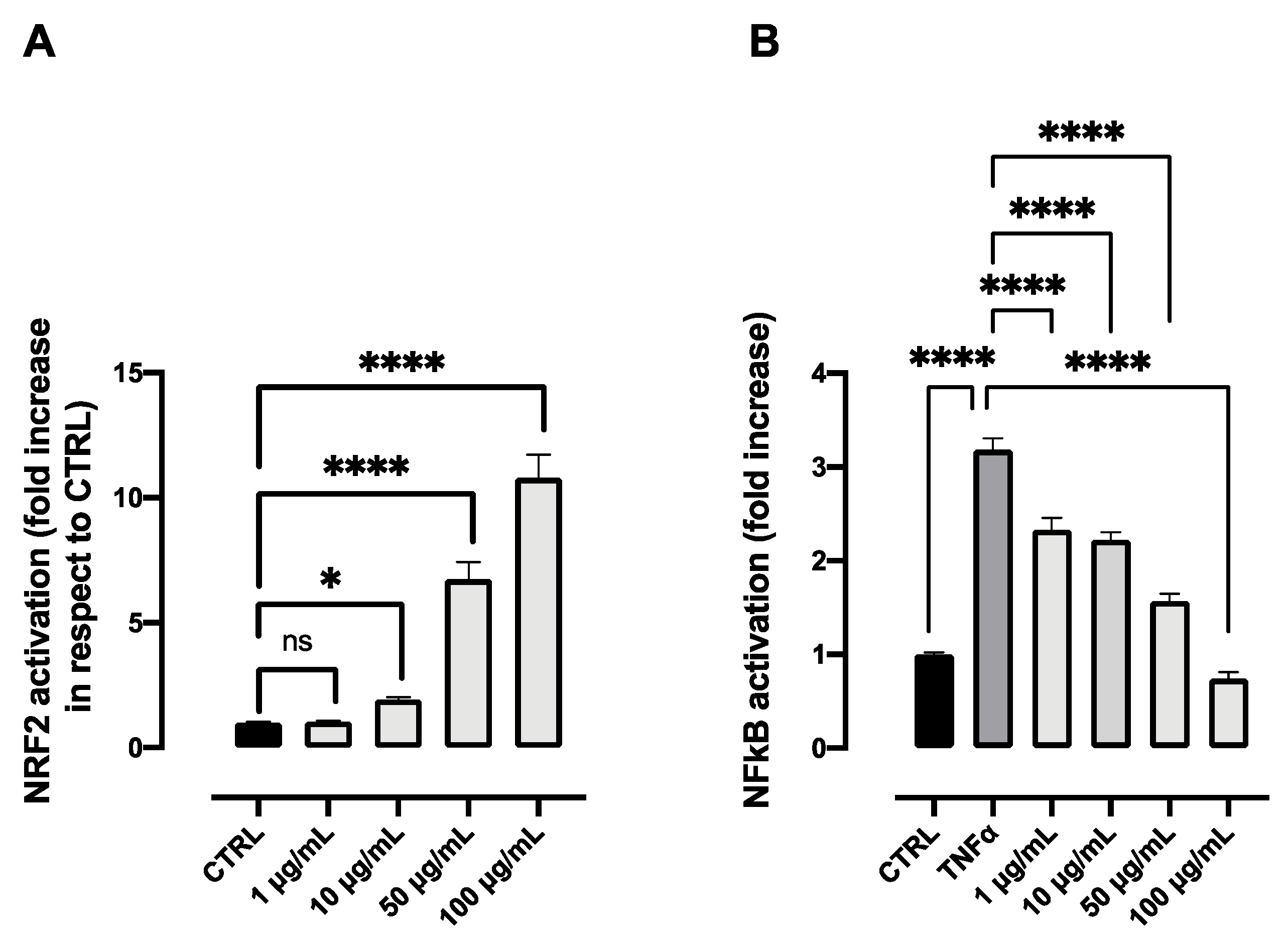
2.4. Biological Activity
3. Materials and Methods
3.1. Ethnobotanical Research—Interview Management and Data Archiving
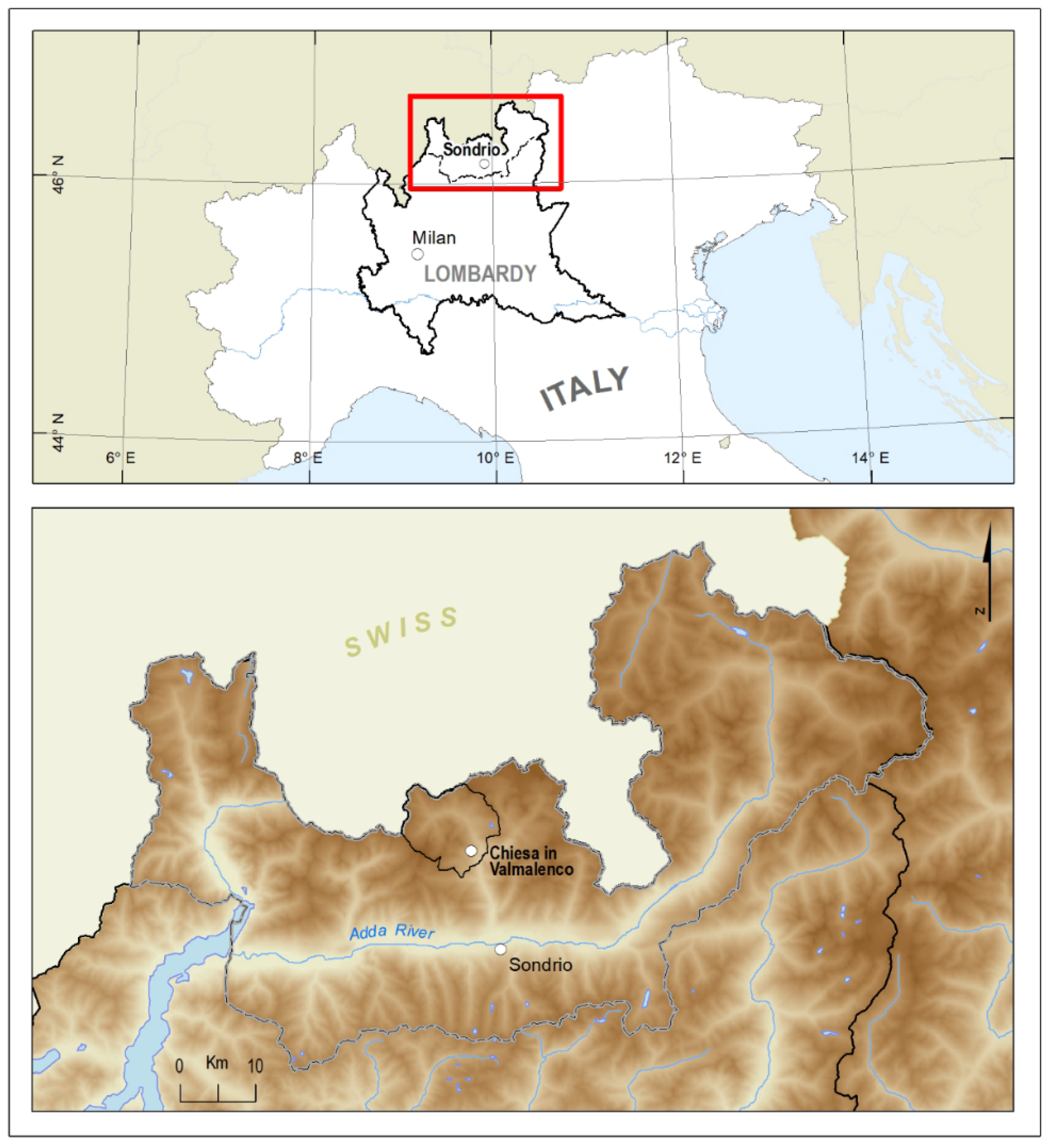
3.2. Plant Material
3.3. Phytochemical Investigation
3.3.1. Preparation of the Decoction
3.3.2. Preparation of the Methanolic Extract
3.3.3. Chemicals
3.3.4. LC-MS Conditions
3.4. Morphological Investigation
3.4.1. Botanical Line Drawings
3.4.2. Microscopy
Light Microscopy (LM) and Fluorescence Microscopy (FM)
Scanning Electron Microscopy (SEM)
3.5. Biological Activity
3.5.1. Antioxidant Activity
3.5.2. Anti-Inflammatory Activity
3.5.3. Cell Viability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bottoni, M.; Milani, F.; Colombo, L.; Nallio, K.; Colombo, P.S.; Giuliani, C.; Bruschi, P.; Fico, G. Using Medicinal Plants in Valmalenco (Italian Alps): From Tradition to Scientific Approaches. Molecules 2020, 25, 4144. [Google Scholar] [CrossRef]
- Dei Cas, L.; Pugni, F.; Fico, G. Tradition of use on medicinal species in Valfurva (Sondrio, Italy). J. Ethnopharmacol. 2015, 163, 113–134. [Google Scholar] [CrossRef]
- Abbet, C.; Mayor, R.; Roguet, D.; Spichiger, R.; Hamburger, M.; Potterat, O. Ethnobotanical survey on wild alpine food plants in Lower and Central Valais (Switzerland). J. Ethnopharmacol. 2014, 151, 624–634. [Google Scholar] [CrossRef]
- Mattalia, G.; Quave, C.L.; Pieroni, A. Traditional uses of wild food and medicinal plants among Brigasc, Kyé, and Provençal communities on the Western Italian Alps. Genet. Resour. Crop Evol. 2013, 60, 587–603. [Google Scholar] [CrossRef]
- Vitalini, S.; Iriti, M.; Puricelli, C.; Ciuchi, D.; Segale, A.; Fico, G. Traditional knowledge on medicinal and food plants used in Val San Giacomo (Sondrio, Italy)—An alpine ethnobotanical study. J. Ethnopharmacol. 2013, 145, 517–529. [Google Scholar] [CrossRef]
- Pieroni, A.; Giusti, M.E. Alpine ethnobotany in Italy: Traditional knowledge of gastronomic and medicinal plants among the Occitans of the upper Varaita valley, Piedmont. J. Ethnobiol. Ethnomed. 2009, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Maffei, M.; Chialva, F.; Codignola, A. Essential oils and chromosome numbers from Italian Achillea species. J. Essent. Oil Res. 1989, 1, 57–64. [Google Scholar] [CrossRef]
- Tava, A.; Iriti, M.; Vitalini, S. Composition and Antioxidant Activity of the Essential Oil from Achillea moschata Wulfen Growing in Valchiavenna and Valmalenco (Italian Central Alps) Introduction. Int. J. Hortic. Sci. Technol. 2020, 7, 335–341. [Google Scholar] [CrossRef]
- Vitalini, S.; Garzoli, S.; Sisto, F.; Pezzani, R.; Argentieri, M.P.; Scarafoni, A.; Ciappellano, S.; Zorzan, M.; Capraro, J.; Collazuol, D.; et al. Digestive and gastroprotective effects of Achillea erba-rotta subsp. moschata (Wulfen) I. Richardson (syn. A. moschata Wulfen) (Asteraceae): From traditional uses to preclinical studies. J. Ethnopharmacol. 2022, 298, 115670. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, S.; Grande, S.; Visioli, F.; Agradi, E.; Fico, G.; Tome, F. Antioxidant activity of wild plants collected in Valsesia, an Alpine Region of Northern Italy. Phyther. Res. 2006, 20, 576–580. [Google Scholar] [CrossRef]
- Vitalini, S.; Madeo, M.; Tava, A.; Iriti, M.; Vallone, L.; Avato, P.; Cocuzza, C.E.; Simonetti, P.; Argentieri, M.P. Chemical profile, antioxidant and antibacterial activities of Achillea moschata Wulfen, an endemic species from the alps. Molecules 2016, 21, 830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argentieri, M.P.; Madeo, M.; Avato, P.; Iriti, M.; Vitalini, S. Polyphenol content and bioactivity of Achillea moschata from the Italian and Swiss Alps. Zeitschrift fur Naturforsch. Sect. C J. Biosci. 2020, 75, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Bottoni, M.; Baron, G.; Milani, F.; Colombo, L.; Colombo, P.S.; Santagostini, L.; Giuliani, C.; Bruschi, P.; Aldini, G.; Fico, G. Achillea moschata Wulfen: Traditional uses and chemical characterization. In Proceedings of the XVII Italian Phytochemical Society Congress jointly with Third ICEMAP Congress 2022, Bari, Italy, 22–24 June 2022. [Google Scholar]
- Valant-Vetschera, K.M.; Wollenweber, E. Exudate flavonoid aglycones in the alpine species of Achillea sect. Ptarmica: Chemosystematics of A. moschata and related species (Compositae-Anthemideae). Biochem. Syst. Ecol. 2001, 29, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Apel, L.; Lorenz, P.; Urban, S.; Sauer, S.; Spring, O.; Stintzing, F.C.; Kammerer, D.R. Phytochemical characterization of different yarrow species (Achillea sp.) and investigations into their antimicrobial activity. Zeitschrift fur Naturforsch. Sect. C J. Biosci. 2021, 76, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Afshari, M.; Rahimmalek, M. Variation in essential oil composition, anatomical, and antioxidant characteristics of Achillea filipendulina Lam. as affected by different phenological stages. J. Essent. Oil Res. 2021, 33, 283–298. [Google Scholar] [CrossRef]
- Fathi, E.; Majdi, M.; Dastan, D.; Maroufi, A. The spatio-temporal expression of some genes involved in the biosynthetic pathways of terpenes/phenylpropanoids in yarrow (Achillea millefolium). Plant Physiol. Biochem. 2019, 142, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Afshari, M.; Rahimmalek, M. Variation in Essential Oil Composition, Bioactive Compounds, Anatomical and Antioxidant Activity of Achillea aucheri, an Endemic Species of Iran, at Different Phenological Stages. Chem. Biodivers. 2018, 15, e1800319. [Google Scholar] [CrossRef] [PubMed]
- Nedelcheva, A. Micro-morphology of Achillea clypeolata: Contribution to the pharmacognostical profile. J. Appl. Pharm. Sci. 2012, 2, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Ciccarelli, D.; Garbari, F.; Pagni, A.M. Glandular hairs of the ovary: A helpful character for Asteroideae (Asteraceae) taxonomy? Ann. Bot. Fenn. 2007, 44, 1–7. [Google Scholar]
- Figueiredo, A.C.; Pais, M.S.S. Ultrastructural Aspects of the Glandular Cells from the Secretory Trichomes and from the Cell Suspension Cultures of Achillea millefolium L. ssp. millefolium. Ann. Bot. 1994, 74, 179–190. [Google Scholar] [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Raudone, L.; Radušiene, J.; Seyis, F.; Yayla, F.; Vilkickyte, G.; Marksa, M.; Ivanauskas, L.; Cırak, C. Distribution of phenolic compounds and antioxidant activity in plant parts and populations of seven underutilized wild Achillea species. Plants 2022, 11, 447. [Google Scholar] [CrossRef] [PubMed]
- Afshari, M.; Rahimmalek, M.; Miroliaei, M. Variation in polyphenolic profiles, antioxidant and antimicrobial activity of different Achillea species as natural sources of antiglycative compounds. Chem. Biodivers. 2018, 15, e1800075. [Google Scholar] [CrossRef]
- Vitalini, S.; Tomè, F.; Fico, G. Traditional uses of medicinal plants in Valvestino (Italy). J. Ethnopharmacol. 2009, 121, 106–116. [Google Scholar] [CrossRef]
- Cornara, L.; La Rocca, A.; Terrizzano, L.; Dente, F.; Mariotti, M.G. Ethnobotanical and phytomedical knowledge in the North-Western Ligurian Alps. J. Ethnopharmacol. 2014, 155, 463–484. [Google Scholar] [CrossRef]
- Vitalini, S.; Puricelli, C.; Mikerezi, I.; Iriti, M. Plants, people and traditions: Ethnobotanical survey in the Lombard Stelvio National Park and neighbouring areas (Central Alps, Italy). J. Ethnopharmacol. 2015, 173, 435–458. [Google Scholar] [CrossRef]
- Bruschi, P.; Sugni, M.; Moretti, A.; Signorini, M.A.; Fico, G. Children’s versus adult’s knowledge of medicinal plants: An ethnobotanical study in Tremezzina (Como, Lombardy, Italy). Brazilian J. Pharmacogn. 2019, 29, 644–655. [Google Scholar] [CrossRef]
- Si, X.T.; Zhang, M.L.; Shi, Q.W.; Kiyota, H. Chemical constituents of the plants in the genus Achillea. Chem. Biodivers. 2006, 3, 1163–1180. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole—New Business Media: Bologna, Italy, 2017. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Forman, H.J.; Davies, K.J.; Ursini, F. How do nutritional antioxidants really work: Nucleophilic tone and para-hormesis versus free radical scavenging in vivo. Free Radic. Biol. Med. 2014, 66, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Luo, L.; Namani, A.; Wang, X.; Tang, X. Nrf2 Signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta. Mol. Basis. Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef] [PubMed]
- Bensouici, C.; Kabouche, A.; Karioti, A.; Öztürk, M.; Duru, M.E.; Bilia, A.R.; Kabouche, Z. Compounds from Sedum caeruleum with antioxidant, anticholinesterase, and antibacterial activities. Pharm. Biol. 2016, 54, 174–179. [Google Scholar] [CrossRef]
- Villalva, M.; Jaime, L.; Villanueva-Bermejo, D.; Lara, B.; Fornari, T.; Reglero, G.; Santoyo, S. Supercritical anti-solvent fractionation for improving antioxidant and anti-inflammatory activities of an Achillea millefolium L. extract. Food Res. Int. 2019, 115, 128–134. [Google Scholar] [CrossRef]
- Vitalini, S.; Beretta, G.; Iriti, M.; Orsenigo, S.; Basilico, N.; Dall’Acqua, S.; Iorizzi, M.; Fico, G. Phenolic compounds from Achillea millefolium L. and their bioactivity. Acta Biochim. Pol. 2011, 58, 203–209. [Google Scholar] [CrossRef]
- Wang, W.; Yue, R.F.; Jin, Z.; He, L.M.; Shen, R.; Du, D.; Tang, Y.Z. Efficiency comparison of apigenin-7-O-glucoside and trolox in antioxidative stress and anti-inflammatory properties. J. Pharm. Pharmacol. 2020, 72, 1645–1656. [Google Scholar] [CrossRef]
- Witkowska-Banaszczak, E.; Krajka-Kuźniak, V.; Papierska, K. The effect of luteolin 7-glucoside, apigenin 7-glucoside and Succisa pratensis extracts on NF-κB activation and α-amylase activity in HepG2 cells. Acta Biochim. Pol. 2020, 67, 41–47. [Google Scholar] [CrossRef]
- Sun, Q.; Lu, N.N.; Feng, L. Apigetrin inhibits gastric cancer progression through inducing apoptosis and regulating ROS-modulated STAT3/JAK2 pathway. Biochem. Biophys. Res. Commun. 2018, 498, 164–170. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Sun-Waterhouse, D. The potential of dandelion in the fight against gastrointestinal diseases: A review. J. Ethnopharmacol. 2022, 293, 115272. [Google Scholar] [CrossRef]
- Puangpraphant, S.; Berhow, M.A.; Vermillion, K.; Potts, G.; Gonzalez de Mejia, E. Dicaffeoylquinic acids in Yerba mate (Ilex paraguariensis St. Hilaire) inhibit NF-κB nucleus translocation in macrophages and induce apoptosis by activating caspases-8 and -3 in human colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 1509–1522. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Xie, M.; Chen, G.; Dai, Z.; Hu, B.; Zeng, X.; Sun, Y. Anti-inflammatory effects of dicaffeoylquinic acids from Ilex kudingcha on lipopolysaccharide-treated RAW264.7 macrophages and potential mechanisms. Food Chem. Toxicol. 2019, 126, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Karbab, A.; Charef, N.; Abu Zarga, M.H.; Qadri, M.I.; Mubarak, M.S. Ethnomedicinal documentation and anti-inflammatory effects of n-butanol extract and of four compounds isolated from the stems of Pituranthos scoparius: An in vitro and in vivo investigation. J. Ethnopharmacol. 2021, 267, 113488. [Google Scholar] [CrossRef]
- Han, Y.; Zhou, M.; Wang, L.; Ying, X.; Peng, J.; Jiang, M.; Bai, G.; Luo, G. Comparative evaluation of different cultivars of Flos Chrysanthemi by an anti-inflammatory-based NF-κB reporter gene assay coupled to UPLC-Q/TOF MS with PCA and ANN. J. Ethnopharmacol. 2015, 174, 387–395. [Google Scholar] [CrossRef]
- Martínez-Vázquez, M.; Estrada-Reyes, R.; Martínez-Laurrabaquio, A.; López-Rubalcava, C.; Heinze, G. Neuropharmacological study of Dracocephalum moldavica L. (Lamiaceae) in mice: Sedative effect and chemical analysis of an aqueous extract. J. Ethnopharmacol. 2012, 141, 908–917. [Google Scholar] [CrossRef]
- Cheng, D.M.; Roopchand, D.E.; Poulev, A.; Kuhn, P.; Armas, I.; Johnson, W.D.; Oren, A.; Ribnicky, D.; Zelzion, E.; Bhattacharya, D.; et al. High phenolics Rutgers Scarlet Lettuce improves glucose metabolism in high fat diet-induced obese mice. Mol. Nutr. Food Res. 2016, 60, 2367–2378. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Zhang, H.; Yang, R.; Hui, Q.; Chen, Y.; Mats, L.; Tsao, R.; Yang, C. Red-osier dogwood extracts prevent inflammatory responses in Caco-2 cells and a Caco-2 BBe1/EA.hy926 cell co-culture model. Antioxidants 2019, 8, 428. [Google Scholar] [CrossRef] [Green Version]
- Baron, G.; Ferrario, G.; Marinello, C.; Carini, M.; Morazzoni, P.; Aldini, G. Effect of extraction solvent and temperature on polyphenol profiles, antioxidant and anti-inflammatory effects of red grape skin by-product. Molecules 2021, 26, 5454. [Google Scholar] [CrossRef]
- Giuliani, C.; Maleci Bini, L. Insight into the structure and chemistry of glandular trichomes of Labiatae, with emphasis on subfamily Lamioideae. Plant Syst. Evol. 2008, 276, 199–208. [Google Scholar] [CrossRef]
- Giuliani, C.; Bottoni, M.; Milani, F.; Todero, S.; Berera, P.; Maggi, F.; Santagostini, L.; Fico, G. Botanic garden as a factory of molecules: Myrtus communis L. subsp. communis as a case study. Plants 2022, 11, 754. [Google Scholar] [CrossRef]
- Ferrario, G.; Baron, G.; Gado, F.; Della Vedova, L.; Bombardelli, E.; Carini, M.; D’Amato, A.; Aldini, G.; Altomare, A. Polyphenols from thinned young apples: HPLC-HRMS profile and evaluation of their anti-oxidant and anti-inflammatory activities by proteomic studies. Antioxidants 2022, 11, 1577. [Google Scholar] [CrossRef] [PubMed]
- Baron, G.; Altomare, A.; Mol, M.; Garcia, J.L.; Correa, C.; Raucci, A.; Mancinelli, L.; Mazzotta, S.; Fumagalli, L.; Trunfio, G.; et al. Analytical profile and antioxidant and anti-inflammatory activities of the enriched polyphenol fractions isolated from bergamot fruit and leave. Antioxidants 2021, 10, 141. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Number | Name | RT (min) | [M − H]− exp | MS/MS | [M + H]+ exp | MS/MS |
|---|---|---|---|---|---|---|
| 1 | Caffeoylquinic acid isomer 1 | 3.1 | 353.08667 | 179-191 | 355.10245 | 163 |
| 2 | Caffeoylquinic acid isomer 2 | 5.1 | 353.08630 | 161-179-191 | 355.10239 | 163 |
| 3 | Vicenin-2 | 9.5 | 593.14972 | 353-383-473-503 | 595.16553 | 475 |
| 4 | Luteolin C-glucoside * | 13.7 | 447.09320 | 285-327-357 | 449.10782 | 383-353-329-287 |
| 5 | Schaftoside | 14.5 | 563.14001 | 353-383-443-473-503 | 565.15497 | 475-445 |
| 6 | Quercetin 3-O-rutinoside (Rutin) | 19.9 | 609.14416 | 301 | 611.16046 | 449-303 |
| 7 | Kaempferol O-glucoside | 21.2 | 447.09320 | 285 | 449.10779 | 287 |
| 8 | Luteolin O-glucoside | 22.1 | 447.09320 | 285 | 449.10782 | 287 |
| 9 | Mearnsetin hexoside isomer 1 | 22.1 | 493.09900 | 316-331 | 495.11279 | 333 |
| 10 | Mearnsetin hexoside isomer 2 | 23.3 | 493.09903 | 316-331 | - | - |
| 11 | Dicaffeoylquinic acid isomer 1 | 26.2 | 515.11975 | 179-335-353 | - | - |
| 12 | Isorhamnetin O-hexoside isomer 1 | 26.0 | 477.10420 | 300-315 | 479.11804 | 317 |
| 13 | Dicaffeoylquinic acid isomer 2 | 27.6 | 515.11987 | 179-191-335-353 | - | - |
| 14 | Isorhamnetin O-rutinoside | 29.0 | 623.15900 | 300-315 | 625.17625 | 479-317 |
| 15 | Apigenin 7-glucoside (Cosmosiin) | 29.8 | 431.09821 | 269 | 433.11282 | 271 |
| 16 | Isorhamnetin O-hexoside isomer 2 | 30.3 | 477.10403 | 300-315 | 479.11876 | 317 |
| 17 | Syringetin 3-O-glucoside | 33.0 | 507.11465 | 345 | 509.12839 | 347 |
| 18 | Isoorientin 7-methyl ether | 33.3 | 461.10919 | 284-299 | 463.12344 | 301 |
| 19 | Dicaffeoylquinic acid isomer 3 | 33.8 | 515.11969 | 179-191-335-353 | - | - |
| 20 | Luteolin 7-malonyl-glucoside | 34.5 | 533.09357 | 489-285 | 535.10779 | 449-287 |
| 21 | Eupatolin | 36.8 | 491.11972 | 299313-314-328-329-343 | 493.13373 | 331 |
| 22 | Apigenin 7-malonyl-glucoside | 43.6 | 517.09894 | 473-269 | 519.11273 | 433-271 |
| 23 | Luteolin | 45.9 | 285.04028 | 107-133-151-175-199-217-241-257 | 287.05527 | 241-153 |
| 24 | Axillarin | 51.4 | 345.06122 | 271-287-300-315 | 347.07651 | 314-285-257 |
| 25 | Apigenin | 52.9 | 269.04556 | 107-149-225 | 271.06033 | 153-119 |
| 26 | Chrysoeriol/Hispidulin | 53.9 | 299.05542 | 256-284 | 301.0708 | 286-153 |
| 27 | Desmethoxycentaureidin * | 54.1 | 329.06641 | 299-314 | 331.08151 | 270-243 |
| 28 | Isorhamnetin * | 54.4 | 315.05066 | 300 | 317.06577 | 285-257-229-153-119 |
| 29 | Quercetin 3,3’-dimethyl ether | 54.8 | 329.06674 | - | 331.08145 | 301-273 |
| 30 | 6-Hydroxykaempferol 3,6-dimethyl ether | 55.5 | 329.06650 | - | 331.08142 | 315-287 |
| 31 | Chrysosplenol-D/Jaceidin | 56.4 | 359.07712 | 314-329-341 | 361.09195 | 331 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bottoni, M.; Baron, G.; Gado, F.; Milani, F.; Santagostini, L.; Colombo, L.; Colombo, P.S.; Caporali, E.; Spada, A.; Biagi, M.; et al. Achillea moschata Wulfen: From Ethnobotany to Phytochemistry, Morphology, and Biological Activity. Molecules 2022, 27, 8318. https://doi.org/10.3390/molecules27238318
Bottoni M, Baron G, Gado F, Milani F, Santagostini L, Colombo L, Colombo PS, Caporali E, Spada A, Biagi M, et al. Achillea moschata Wulfen: From Ethnobotany to Phytochemistry, Morphology, and Biological Activity. Molecules. 2022; 27(23):8318. https://doi.org/10.3390/molecules27238318
Chicago/Turabian StyleBottoni, Martina, Giovanna Baron, Francesca Gado, Fabrizia Milani, Laura Santagostini, Lorenzo Colombo, Paola Sira Colombo, Elisabetta Caporali, Alberto Spada, Marco Biagi, and et al. 2022. "Achillea moschata Wulfen: From Ethnobotany to Phytochemistry, Morphology, and Biological Activity" Molecules 27, no. 23: 8318. https://doi.org/10.3390/molecules27238318
APA StyleBottoni, M., Baron, G., Gado, F., Milani, F., Santagostini, L., Colombo, L., Colombo, P. S., Caporali, E., Spada, A., Biagi, M., Giuliani, C., Bruschi, P., Aldini, G., & Fico, G. (2022). Achillea moschata Wulfen: From Ethnobotany to Phytochemistry, Morphology, and Biological Activity. Molecules, 27(23), 8318. https://doi.org/10.3390/molecules27238318