


Transcriptional Responses for Biosynthesis of Triterpenoids in Exogenous Inducers Treated Inonotus Hispidus Using RNA-Seq


Abstract
:1. Introduction
2. Results
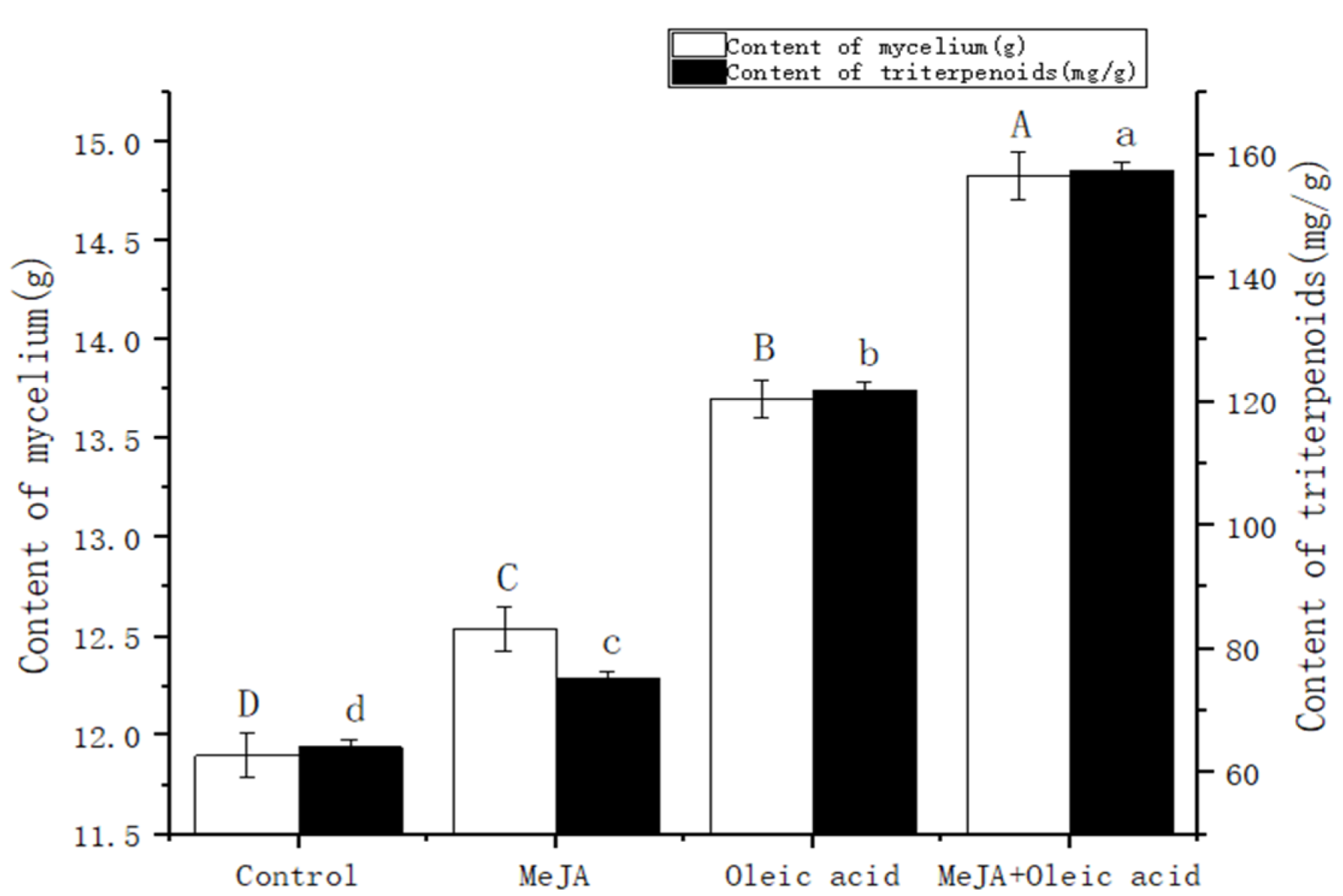
2.1. Effect of Inducers on Triterpenoid Content of I. hispidus
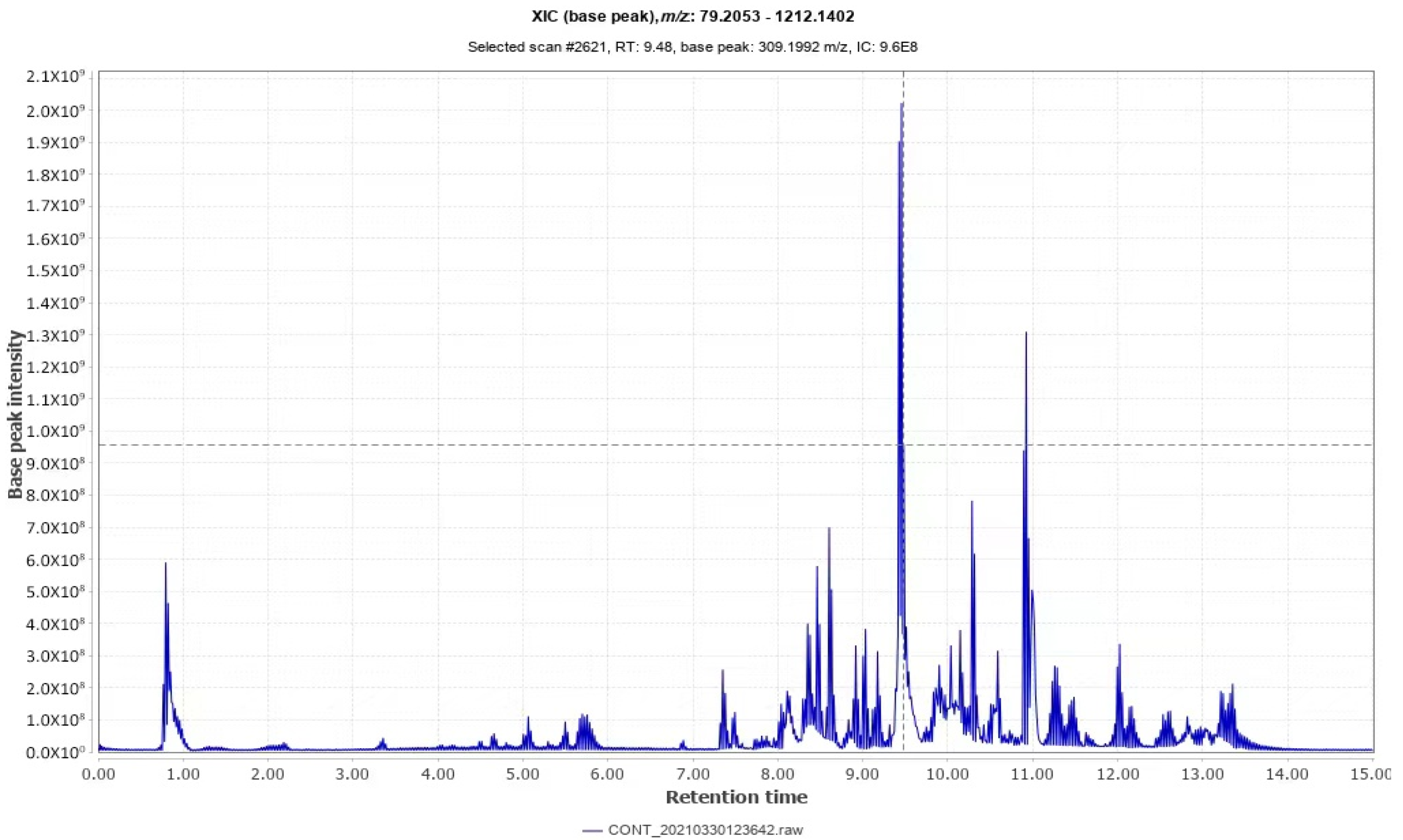
2.2. Identification of Triterpenoid of I. hispidus
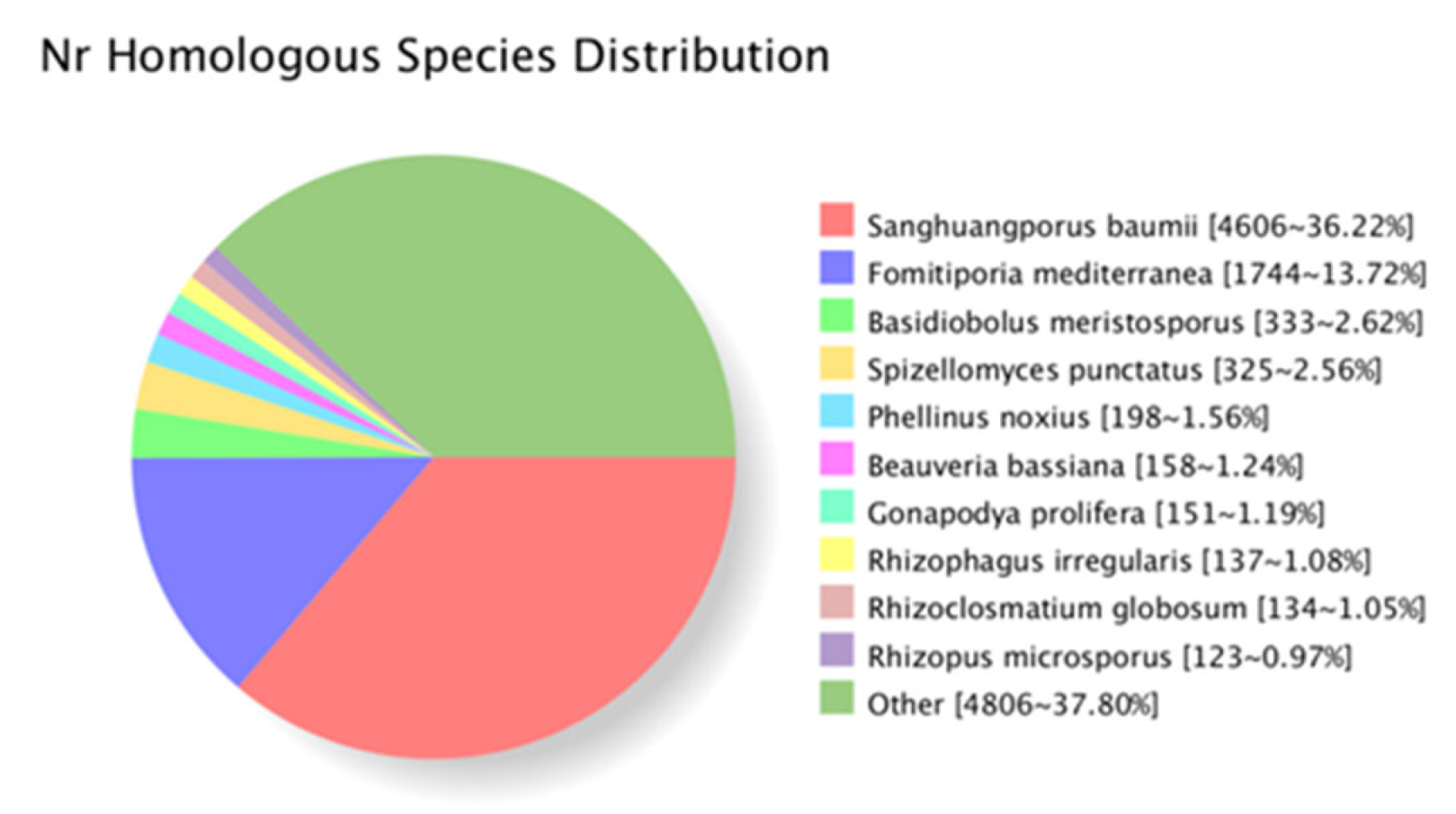
2.3. RNA-Seq and De Novo Assembly of I. hispidus Reference Transcriptome
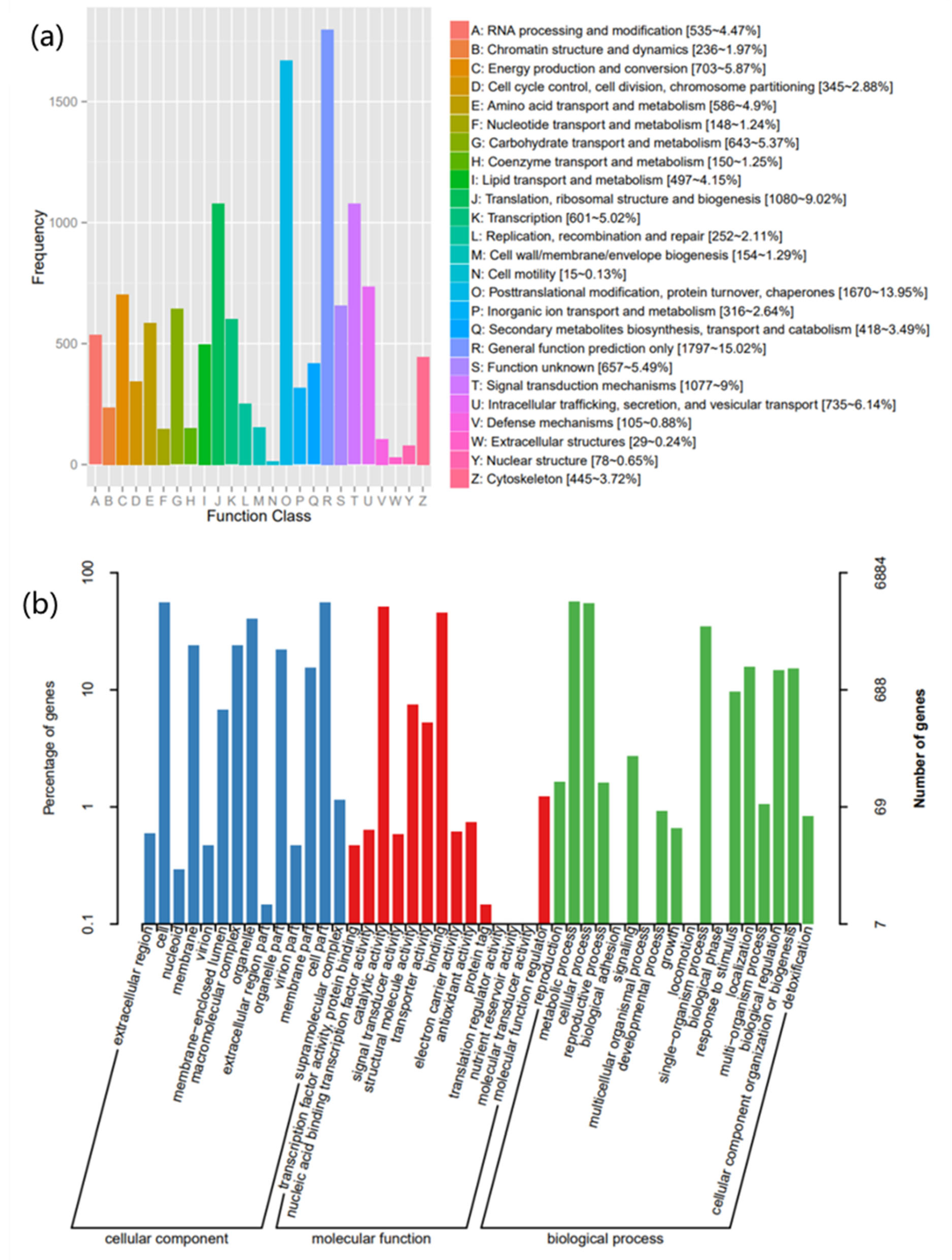
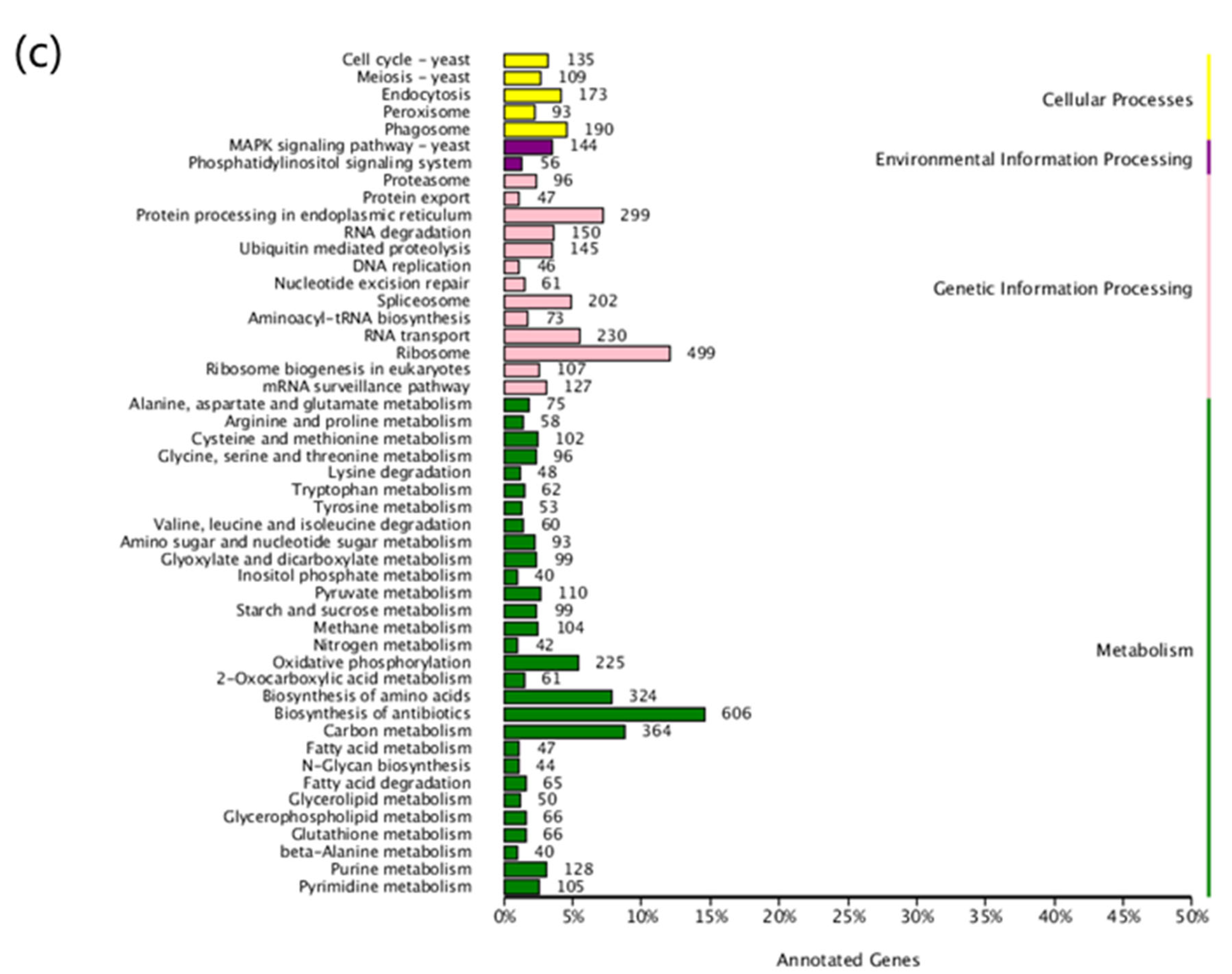
2.4. Functional Annotation and Classification
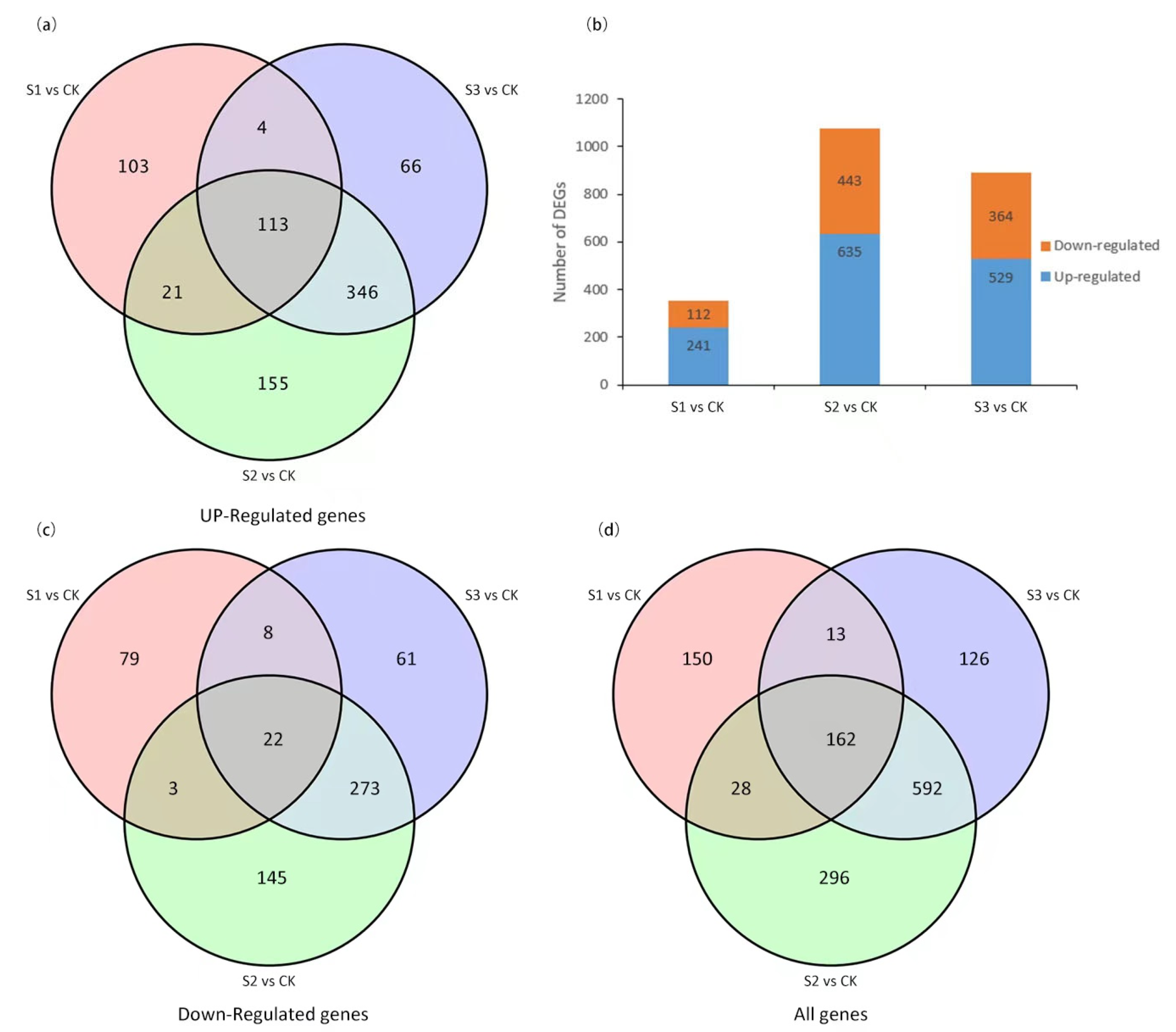
2.5. Identification and Analysis of DEGs
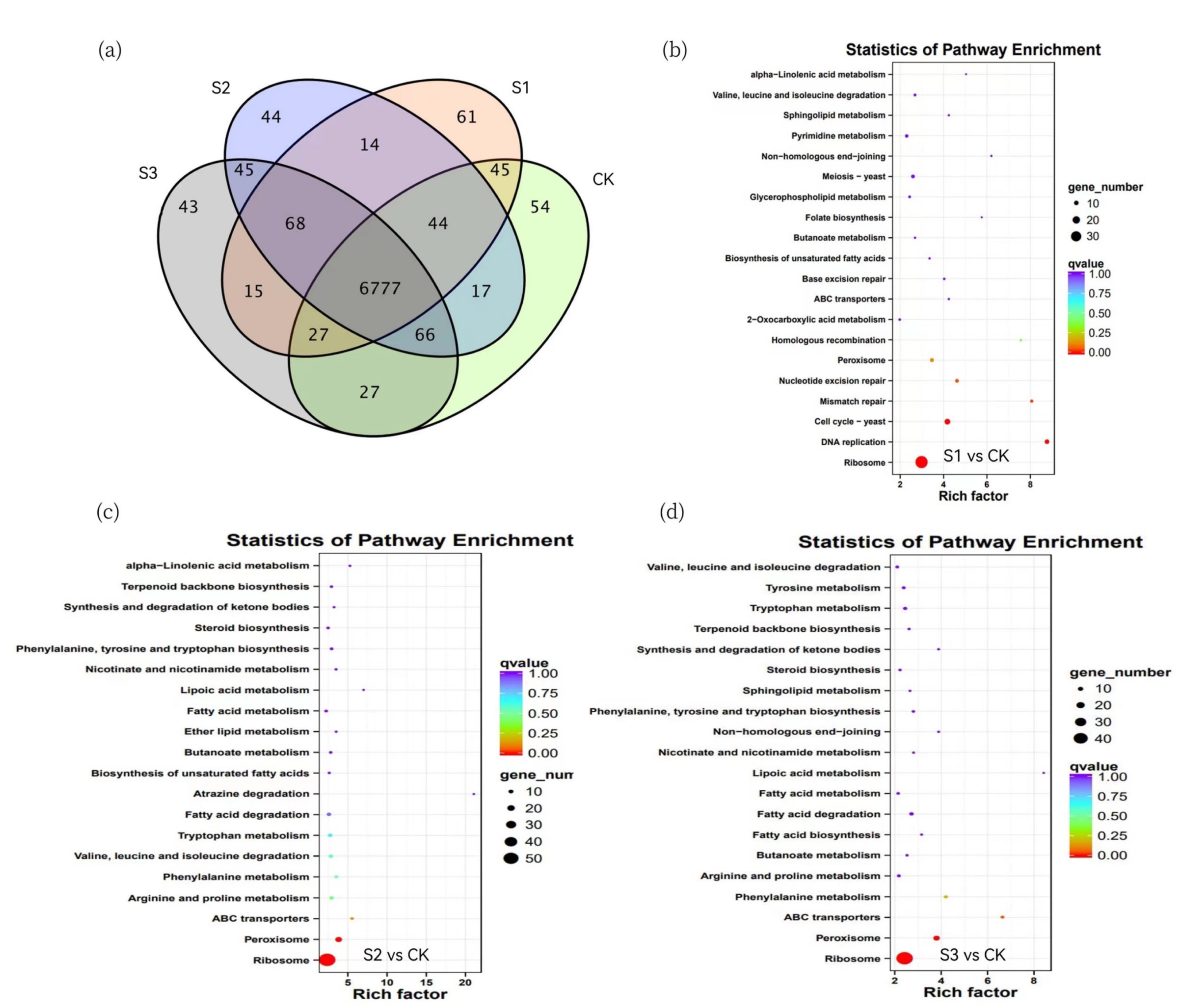
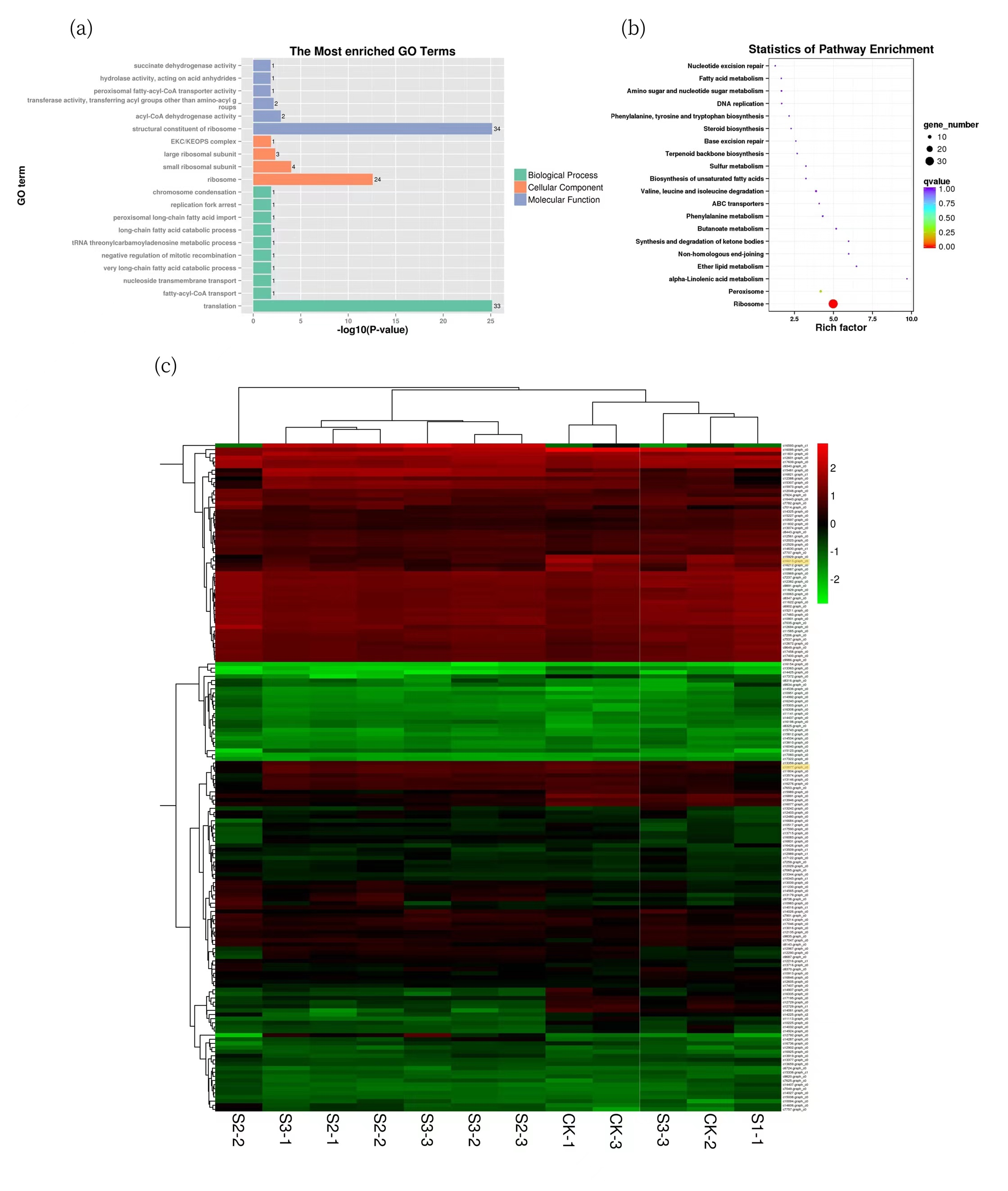
2.6. Enrichment Analysis of DEGs
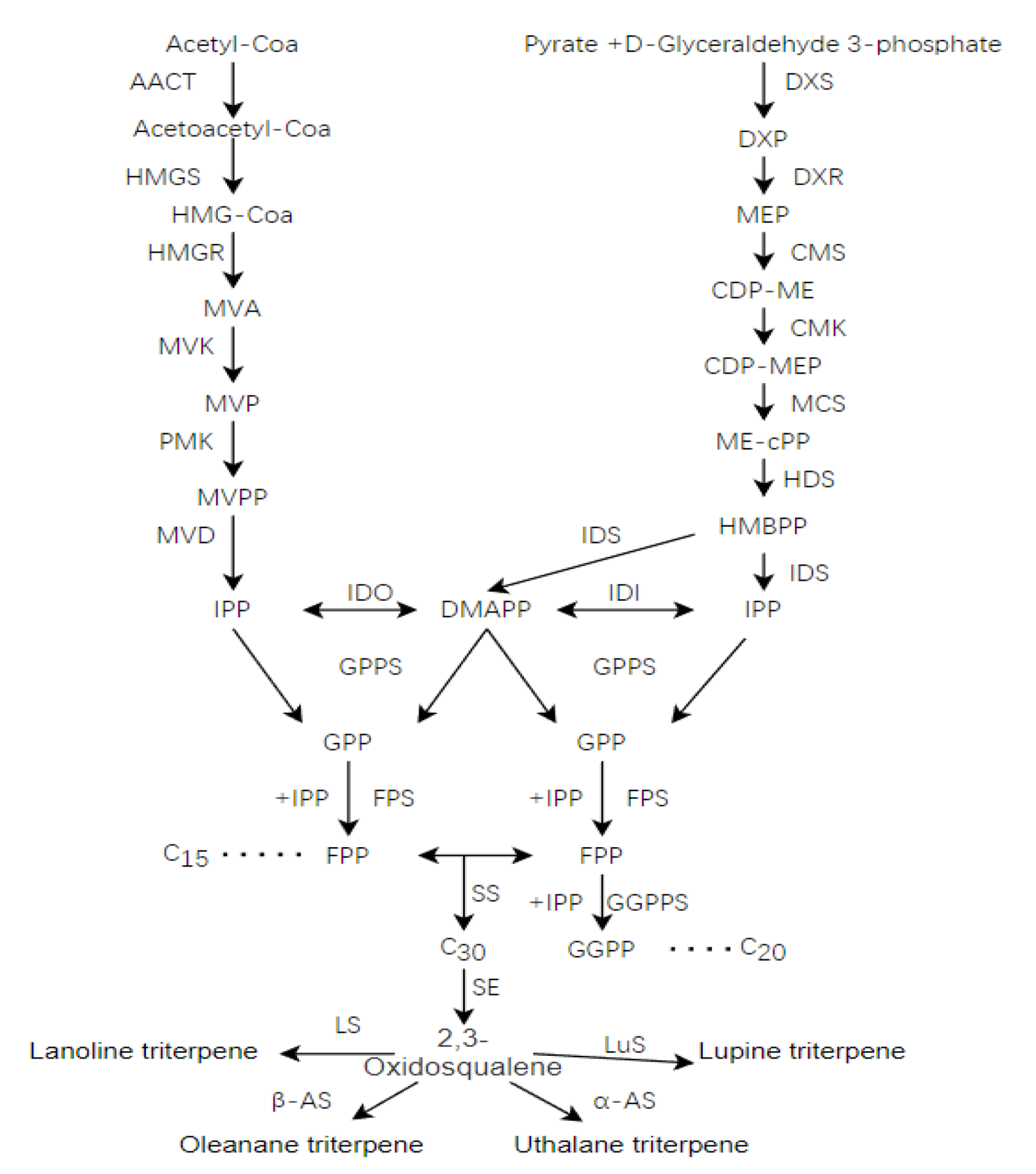
2.7. Putative Genes Involved in Triterpenoid Biosynthesis
2.8. Analysis of CYP450 Defense Genes
2.9. Analysis of Defensive Enzymatic Defense Genes
3. Discussion
4. Materials and Methods
4.1. Experimental Materials and Design
4.2. Measurement of Triterpenoid Content in I. hispidus
4.3. Metabolite Extraction Analyses in I. hispidus
4.4. Library Preparation for Transcriptome Sequencing
4.5. Functional Annotation and Metabolic Pathway Analysis of Illumina HiSeq
4.6. Analysis of Differential Genes
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Liu, X.; Hou, R.L.; Yan, J.J.; Xu, K.Q.; Wu, X.P.; Lin, W.X.; Zheng, M.F.; Fu, J.S. Purification and characterization of Inonotus hispidus exopolysaccharide and its protective effect on acute alcoholic liver injury in mice. Int. J. Biol. Macromol. 2019, 129, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Grundemann, C.; Arnhold, M.; Meier, S.; Backer, C.; Garcia-Kaufer, M.; Grunewald, F.; Steinborn, C.; Klemd, A.M.; Wille, R.; Huber, R. Effects of Inonotus hispidus extracts and compounds on human immunocompetent cells. Eur. J. Integr. Med. 2016, 8 (Suppl. 1), 54. [Google Scholar] [CrossRef]
- Renda, G.; Gokkaya, I.; Sohretoglu, D. Immunomodulatory properties of triterpenes. Phytochem Rev. 2021, 4, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Zhong, X.; Lian, D.H.; Zheng, Y.M.; Wang, H.Z.; Liu, X. Triterpenoid biosynthesis and the transcriptional response elicited by nitric oxide in submerged fermenting Ganoderma lucidum. Process Biochem. 2017, 60, 19–26. [Google Scholar] [CrossRef]
- Adil, B.; Xiang, Q.J.; He, M.L.; Wu, Y.T.; Asghar, M.A.; Arshad, M.; Qin, P.; Gu, Y.F.; Yu, X.M.; Zhao, K.; et al. Effect of sodium and calcium on polysaccharide production and the activities of enzymes involved in the polysaccharide synthesis of Lentinus edodes. AMB Expr. 2020, 10, 47. [Google Scholar]
- Zhong, Q.L.; Hu, H.L.; Fan, B.F.; Zhu, C.; Chen, Z.X. Biosynthesis and Roles of Salicylic Acid in Balancing Stress Response and Growth in Plants. Int. J. Mol. Sci. 2021, 22, 11672. [Google Scholar] [CrossRef]
- Kou, R.W.; Xia, B.; Wang, Z.J.; Li, J.N.; Yang, J.R.; Gao, Y.Q.; Yin, X.; Gao, J.M. Triterpenoids and meroterpenoids from the edible Ganoderma resinaceum and their potential anti-inflammatory, antioxidant and anti-apoptosis activities. Bioorg. Chem. 2022, 121, 105–689. [Google Scholar]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann Bot. 2017, 100, 681–697. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Su, Y.C.; Ling, H.; Ahmad, W.; Gao, S.W.; Guo, J.L.; Que, Y.X.; Xu, L.P. A sugarcane pathogenesis-related protein, ScPR10, plays a positive role in defense responses under Sporisorium scitamineum, SrMV, SA, and MeJA stresses. Plant Cell Rep. 2017, 36, 1427–1440. [Google Scholar]
- Sun, T.T.; Zou, L.; Zhang, L.F.; Zhang, J.; Wang, X.T. Methyl jasmonate induces triterpenoid biosynthesis in Inonotus baumii. Biotechnol. Biotechnol. Equip. 2017, 31, 312–317. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.Q.; Zhang, X.; Chen, C. Stimulated production of triterpenoids of Inonotus obliquus using methyl jasmonate and fatty acids. Ind. Crop Prod. 2017, 85, 49–57. [Google Scholar] [CrossRef]
- Lou, H.; Li, H.W.; Tian, Y. Stimulatory Effects of Oleic Acid and Fungal Elicitor on Betulinic Acid Production by Submerged Cultivation of Medicinal Mushroom Inonotus obliquus. J. Fungi 2021, 7, 266. [Google Scholar] [CrossRef]
- Zhou, J.; Lin, X.Y.; Liu, S.S.; Wang, Z.B.; Liu, D.C.; Huo, Y.H.; Li, D.H. Effects of Compound Elicitors on the Biosynthesis of Triterpenoids and Activity of Defense Enzymes from Inonotus hispidus (Basidiomycetes). Molecules 2021, 27, 2618. [Google Scholar]
- Lin, S.; Zou, Z.C.; Zhou, C.B.; Zhang, H.C.; Cai, Z.M. Transcriptome Analysis Reveals the Molecular Mechanisms Underlying Adenosine Biosynthesis in Anamorph Strain of Caterpillar Fungus. Biomed. Res. Int. 2019, 2019, 1864168. [Google Scholar] [CrossRef]
- Wu, F.F.; Hu, S.P.; Ran, Y.; Chen, X.N.; Lin, S. Gene Mining and Sequence Analysis of Purine Nucleosidase Based on RNA-Seq. JoVE-J. Vis. Exp. 2021, 164, e61561. [Google Scholar]
- Yang, X.Q.; Lin, R.M.; Xu, K.; Guo, L.Z.; Yu, H. Comparative Proteomic Analysis within the Developmental Stages of the Mushroom White Hypsizygus marmoreus. J. Fungi 2022, 7, 1064. [Google Scholar]
- Liu, Y.K.; Jian, Y.Q.; Cao, M.R.; Wang, B.; Qiu, Y.X.; Yuan, H.W.; Zhou, X.D.; Li, B.; Sheng, W.B.; Peng, C.Y.; et al. Seco-Tetracyclic and Seco-Pentacyclic Triterpenoids from Nature: Phyto-chemistry, Biological Activity: A Review. Mini-Rev. Org. Chem. 2022, 19, 188–230. [Google Scholar] [CrossRef]
- Guan, T.; Li, Y.M.; Sun, H.B.; Tang, X.Z.; Qian, Y.S. Effects of Maslinic Acid, a Natural Triterpene, on Glycogen Metabolism in Cultured Cortical Astrocytes. Planta Med. 2009, 75, 1141–1143. [Google Scholar]
- Li, Q.; Fan, Y.S.; Gao, Z.Q.; Fan, K.; Liu, Z.J. Effect of Ursolic Acid and Oleanolic Acid on Osteoblastic Like Cell-Line MC3T3-E1. Pak. Vet. J. 2015, 35, 414–419. [Google Scholar]
- Zhang, X.; Yu, Y.L.; Jiang, S.; Yu, H.; Xiang, Y.Y.; Liu, D.Q.; Qu, Y.; Cui, X.M.; Ge, F. Oleanane-Type Saponins Biosynthesis in Panax notoginseng via Transformation of beta-Amyrin Synthase Gene from Panax japonicus. Agricul. Food. Chem. J. 2019, 67, 1982–1989. [Google Scholar] [CrossRef]
- Hao, D.C.; Chen, S.L.; Osbourn, A.; Kontogianni, V.G.; Liu, L.W.; Jordan, M.J. Temporal transcriptome changes induced by methyl jasmonate in Salvia sclarea. Gene 2015, 558, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Biswas, T.; Dwivedi, U.N. Plant triterpenoid saponins: Biosynthesis, in vitro production, and pharmacological relevance. Protoplasma 2019, 256, 1463–1486. [Google Scholar] [PubMed]
- Li, F.F.; Liu, B.; Li, T.; Wu, Q.W.; Xu, Z.Y.; Gu, Y.H.; Li, W.; Wang, P.L.; Ma, T.; Lei, H.M. Constituents and Biological Activities of Triterpene saponins from Glycyrrhizae Radix et Rhizoma and Its Solubilization Characteristics. Molecules 2020, 25, 3904. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Park, J.; Woo, H.M. Overexpression of the key enzymes in the methylerythritol-4-phosphate pathway in Corynebacterium glutamicum for improving farnesyl diphosphate-derived terpene production. Agric. Food. Chem. 2020, 68, 10780–10796. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.J.; Dong, X.M.; Li, Q.; Chen, Z.X.; Liu, L. An update on the function and regulation of methylerythritol phosphate and mevalonate pathways and their evolutionary dynamics. J. Integr. Plant Biol. 2021, 63, 1211–1226. [Google Scholar] [CrossRef]
- Qiu, H.H.; Su, L.Y.; Wang, H.F.; Zhang, Z.S. Chitosan elicitation of saponin accumulation in Psammosilene tunicoides hairy roots by modulating antioxidant activity, nitric oxide production and differential gene expression. Plant Physiol Biochem. 2021, 166, 115–127. [Google Scholar]
- Zhang, L.Y.; Xu, X.Q.; Badawy, S.; Ihsan, A.; Liu, Z.L.; Xie, C.Q.; Wang, X.; Tao, Y.F. A Review: Effects of Macrolides on CYP450 Enzymes. Curr. Drug Metab. 2020, 21, 928–937. [Google Scholar]
- Yu, H.W.; Liu, M.L.; Yin, M.N.; Shan, T.Y.; Peng, H.S.; Wang, J.T.; Chang, X.W.; Peng, D.Y.; Zha, L.P.; Gui, S.Y. Transcriptome analysis identifies putative genes involved in triterpenoid biosynthesis in Platycodon grandiflorus. Planta 2021, 254, 16–18. [Google Scholar]
- Guo, J.; Ma, X.H.; Cai, Y.; Ma, Y.; Zhan, Z.L.; Zhou, Y.J.; Liu, W.J.; Guan, M.X.; Yang, J.; Cui, G.H. Cytochrome p450 promiscuity leads to a bifurcating biosynthetic pathway for tanshinones. New Phytol. 2016, 210, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.D.; Bao, H.Y.; Wang, H.; Li, Q.J. Anti-tumour Effect and Pharmacokinetics of an Active Ingredient Isolated from Inonotus hispidus. Biol. Pharm. Bull. 2019, 42, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Angelini, P.; Girometta, C.; Tirillini, B.; Moretti, S.; Covino, S.; Cipriani, M.; D’Ellena, E.; Angeles, G.; Federici, E.; Savino, E.; et al. A comparative study of the antimicrobial and antioxidant activities of Inonotus hispidus fruit and their mycelia extracts. Int. J. Food Prop. 2019, 22, 768–783. [Google Scholar] [CrossRef]
- Tan, J.P.; Han, M.X.; Mao, D.; Cheng, S.Y.; Ye, J.B.; Liu, X.M.; Zhang, X.; Zheng, J.R.; Xu, F.; Chen, Z.X. Comparative Transcriptomic Analysis Reveals the Regulatory Mechanism of Terpene Trilactones Improvement by Exogenous Methyl Jasmonate in Ginkgo biloba. Plant Mol. Biol. Rep. 2021, 40, 81–94. [Google Scholar]
- Xu, H.W.; Lu, Y.; Xie, Z.M.; Song, F.B. Changes in nitrogen metabolism and antioxidant enzyme activities of maize tassel in black soils region of northeast China. Front. Plant Sci. 2014, 5, 515. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.L.; Nguyen, T.G. Effects of elicitors on the enhancement of asiaticoside biosynthesis in cell cultures of centella. Chem. Pap. 2012, 66, 642–648. [Google Scholar]
- Ye, W.X.; Hossain, M.A.; Munemasa, S.; Nakamura, Y.; Mori, I.C.; Murata, Y. Endogenous abscisic acid is involved in methyl jasmonate-induced reactive oxygen species and nitric oxide production but not in cytosolic alkalization in Arabidopsis guard cells. J. Plant Physiol. 2013, 170, 1212–1215. [Google Scholar]
- Naeem, M.; Aftab, T.; Ansari, A.A.; Khan, M.M.A. Carrageenan oligomers and salicylic acid act in tandem to escalate artemisinin production by suppressing arsenic uptake and oxidative stress in Artemisia annua (sweet wormwood) cultivated in high arsenic soil. Environ. Sci. Pollut. Res. 2021, 28, 42706–42721. [Google Scholar] [CrossRef]
- Benevenuto, R.F.; Hegland, S.J.; Topper, J.P.; Rydgren, K.; Moe, S.R.; Rodriguez-Saona, C.; Seldal, T. Multiannual effects of induced plant defenses: Are defended plants good or bad neighbors? Ecol. Evol. 2018, 8, 8940–8950. [Google Scholar]
- Chuluunbaatar, B.; Beni, Z.; Dekany, M.; Kovacs, B.; Sarkozy, A.; Datki, Z.; Macsai, L.; Kalman, J.; Hohmann, J.; Vanyolos, A. Triterpenes from the Mushroom Hypholoma lateritium: Isolation, Structure Determination and Investigation in Bdelloid Rotifer Assays. Molecules 2019, 24, 301. [Google Scholar]
- Wang, Y.; Zhang, R.; Liang, Z.C.; Li, S.H. Grape-RNA: A Database for the Collection, Evaluation, Treatment, and Data Sharing of Grape RNA-Seq Datasets. Genes 2020, 11, 315. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Gotz, S.; Garcia, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar]
- Zhao, Y.D.; Li, M.C.; Konate, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts, A Comparative Study of Quantification Measures for the Analysis of RNA-seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [PubMed]
- Shilpashree, H.B.; Sudharshan, S.J.; Shasany, A.K.; Nagegowda, D.A. Molecular characterization of three CYP450 genes reveals their role in withanolides formation and defense in Withania somnifera, the Indian Ginseng. Sci. Rep. 2022, 12, 1602. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Keep Time /min | Adduct | Theoretical Value (m/z) | Measured Value (m/z) | Molecular Formula | Error (ppm) | Inferred Compound |
|---|---|---|---|---|---|---|---|
| 1 | 9.335 | [M − H]− | 488.3491 | 487.3379 | C30H48O5 | 2.1 | calyxagenin |
| 2 | 9.482 | [M − H]− | 488.3491 | 487.3372 | C30H48O5 | −0.7 | Asiatic acid |
| 3 | 9.763 | [M − H]− | 486.3334 | 485.3315 | C30H46O5 | 3.9 | Abrisapogenol I |
| 4 | 11.598 | [M − H]− | 472.3541 | 471.3237 | C30H48O4 | 2.5 | Maslinic acid. |
| 5 | 11.779 | [M − H]− | 486.3697 | 485.3518 | C31H50O4 | 0.78 | Methyl Masridate |
| 6 | 12.200 | [M − H]− | 484.3539 | 483.3307 | C30H48O3 | 3.3 | Ursolic acid. |
| 7 | 12.948 | [M − H]− | 456.3591 | 455.3528 | C30H48O3 | −2.8 | Oleanolic acid |
| 8 | 13.241 | [M − H]− | 470.3742 | 469.3512 | C31H50O3 | 4.2 | methyl oleanate |
| Sample | Clean Reads | Clean Bases | Q30 | GC |
|---|---|---|---|---|
| CK-1 | 28,043,969 | 8.35 G | 94.15 | 52.01 |
| CK-2 | 27,100,025 | 8.07 G | 94.32 | 51.96 |
| CK-3 | 27,536,171 | 8.22 G | 94.21 | 51.87 |
| S1-1 | 30,910,974 | 9.21 G | 94.21 | 51.92 |
| S1-2 | 25,327,950 | 7.55 G | 94.07 | 51.78 |
| S1-3 | 26,032,264 | 7.75 G | 94.31 | 51.91 |
| S2-1 | 34,705,547 | 10.36 G | 94.29 | 51.82 |
| S2-2 | 27,883,532 | 8.30 G | 94.60 | 51.87 |
| S2-3 | 29,087,755 | 8.67 G | 94.14 | 51.85 |
| S3-1 | 32,483,363 | 9.66 G | 94.09 | 51.81 |
| S3-2 | 30,986,102 | 9.22 G | 92.98 | 51.94 |
| S3-3 | 32,679,265 | 9.74 G | 94.17 | 51.91 |
| Length_Interval | 200–300 bp | 300–500 bp | 500 bp–1 kbp | 1 kbp–2 kbp | >2k bp | Total |
|---|---|---|---|---|---|---|
| Number of transcripts | 10947 | 7551 | 12949 | 56309 | 212667 | 300423 |
| Number of unigenes | 9520 | 4738 | 3067 | 2784 | 4197 | 24306 |
| Database | Number of Unigenes | Percentage |
|---|---|---|
| Annotated in COG | 6537 | 33.65% |
| Annotated in GO | 6884 | 35.76% |
| Annotated in KEGG | 6640 | 33.5% |
| Annotated in KOG | 11,968 | 62.17% |
| Annotated in Pfam | 12,430 | 63.98% |
| Annotated in Swiss-prot | 8355 | 43% |
| Annotated in eggNOG | 17,914 | 92.2% |
| Annotated in NR | 12,754 | 65.60% |
| Total unigenes | 19,429 | 100% |
| Gene | Gene Number | Gene ID |
|---|---|---|
| SS | 2 | c7497.graph_c0, c14961.graph_c0 |
| SE | 6 | c16422.graph_c0, c15729.graph_c0, |
| c24729.graph_c0, c20215.graph_c0 | ||
| c61.graph_c0, c19680.graph_c0 | ||
| AACT | 4 | c8971.graph_c0, c16837.graph_c0 |
| c7417.graph_c0, c18532.graph_c0 | ||
| HMGR | 3 | c12846.graph_c0, c17445.graph_c0 |
| c18037.graph_c0, | ||
| FPPS | 2 | c3736.graph_c0, c67.graph_c0 |
| FPS | 2 | c67.graph_c1, c10924.graph_c0 |
| PMK | 2 | c13594.graph_c0, c22627.graph_c0 |
| SM | 1 | c25144.graph_c0 |
| HMGS | 5 | c10205.graph_c0, c16013.graph_c0 |
| c26810.graph_c0, c20761.graph_c0 | ||
| c27696.graph_c0 | ||
| DMD | 1 | c9873.graph_c0 |
| FT | 1 | c15553.graph_c0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, Y.; Liu, D.; Yang, Q.; Sun, C.; Wang, Z.; Li, D. Transcriptional Responses for Biosynthesis of Triterpenoids in Exogenous Inducers Treated Inonotus Hispidus Using RNA-Seq. Molecules 2022, 27, 8541. https://doi.org/10.3390/molecules27238541
Huo Y, Liu D, Yang Q, Sun C, Wang Z, Li D. Transcriptional Responses for Biosynthesis of Triterpenoids in Exogenous Inducers Treated Inonotus Hispidus Using RNA-Seq. Molecules. 2022; 27(23):8541. https://doi.org/10.3390/molecules27238541
Chicago/Turabian StyleHuo, Yonghong, Dongchao Liu, Qin Yang, Changyan Sun, Zhanbin Wang, and Dehai Li. 2022. "Transcriptional Responses for Biosynthesis of Triterpenoids in Exogenous Inducers Treated Inonotus Hispidus Using RNA-Seq" Molecules 27, no. 23: 8541. https://doi.org/10.3390/molecules27238541
APA StyleHuo, Y., Liu, D., Yang, Q., Sun, C., Wang, Z., & Li, D. (2022). Transcriptional Responses for Biosynthesis of Triterpenoids in Exogenous Inducers Treated Inonotus Hispidus Using RNA-Seq. Molecules, 27(23), 8541. https://doi.org/10.3390/molecules27238541