
Antifungal Effect and Inhibition of the Virulence Mechanism of D-Limonene against Candida parapsilosis

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Antifungal Activity
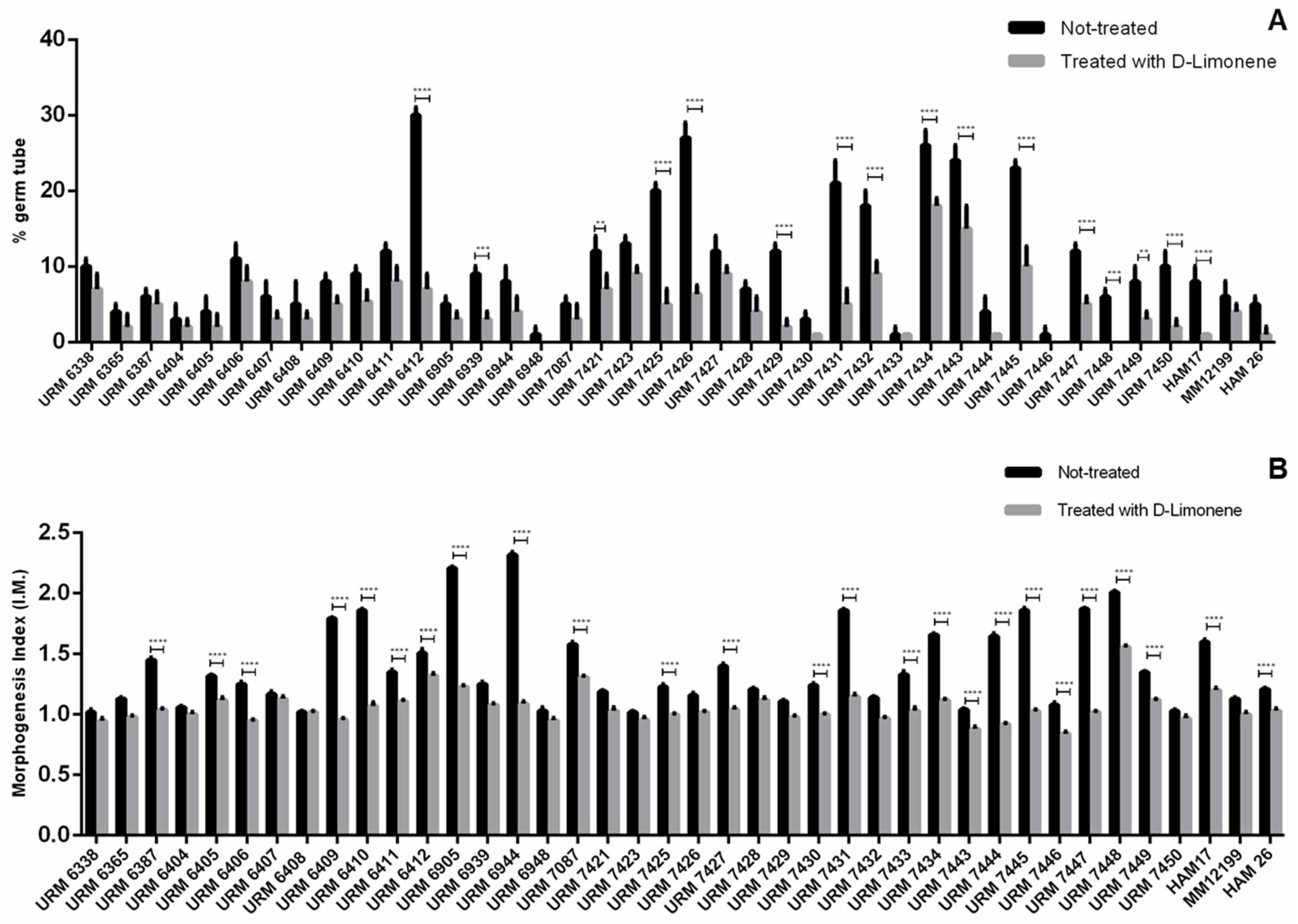
2.2. Morphogenesis Assay
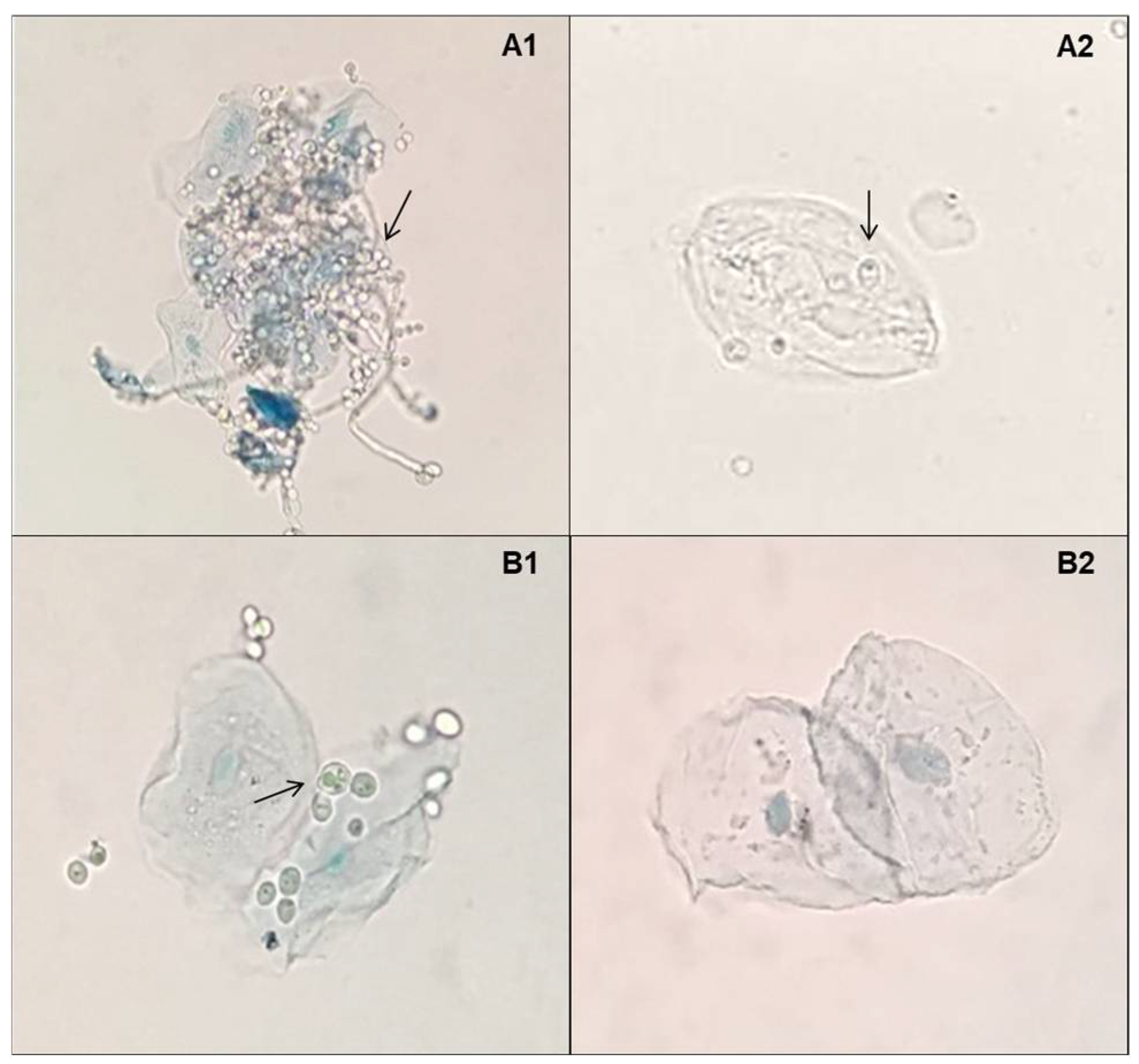
2.3. Adherence Capacity Assay of Yeast Cells to Epithelial Cells
2.4. Evaluation of the D-Limonene Mechanism of Action
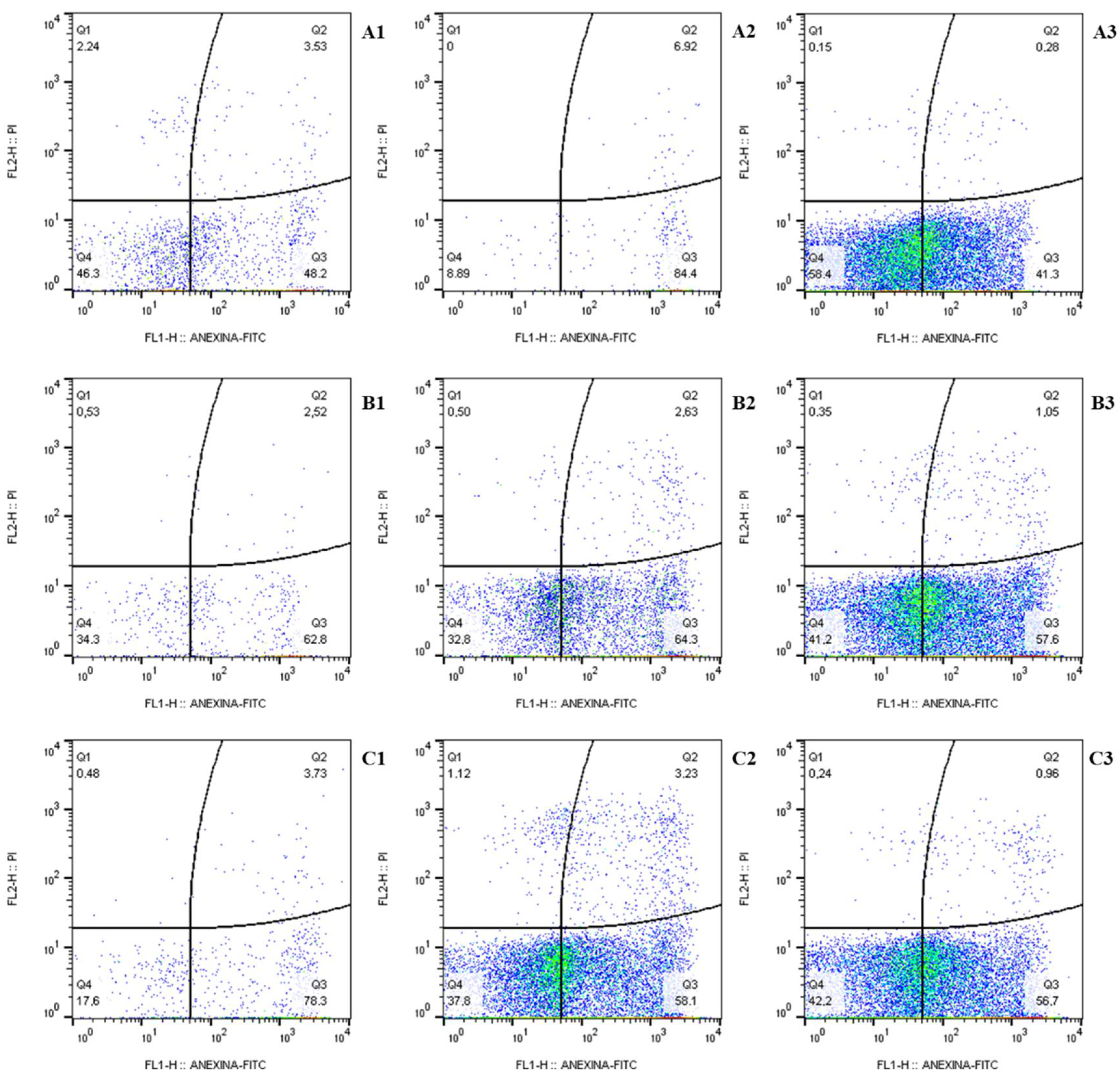
2.4.1. Cell Death Profile
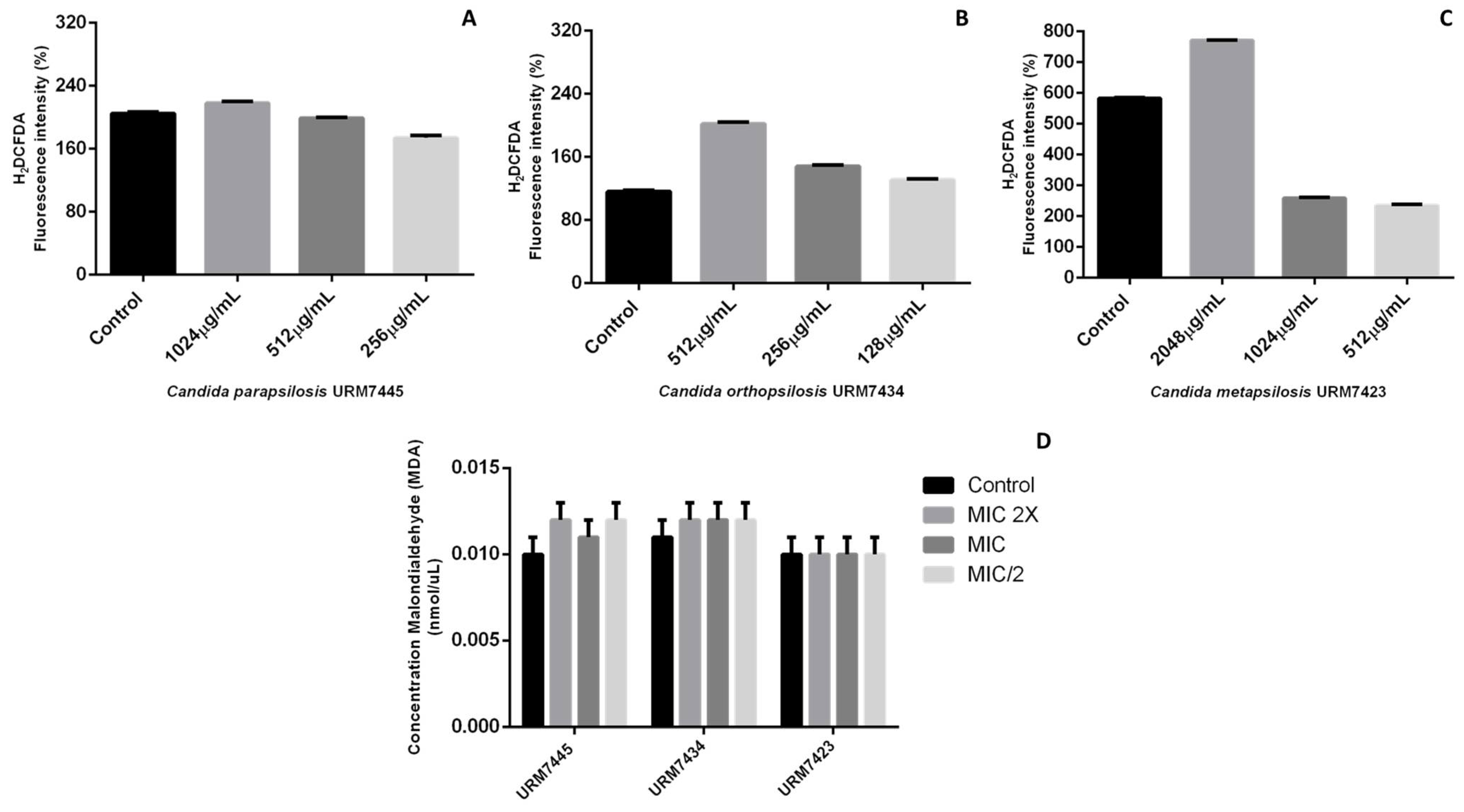
2.4.2. Quantification of Reactive Oxygen Species (ROS) and Lipid Peroxidation
2.4.3. Mitochondrial Membrane Potential Evaluation
3. Discussion
4. Materials and Methods
4.1. Candida Strains and Culture Conditions
4.2. Antifungal Activity
4.3. Virulence Characterization of Yeast Isolates from the C. parapsilosis Complex and D-Limonene Antivirulence Activity
4.3.1. Morphogenesis Assay
4.3.2. Adherence Capacity Assay of Yeast Cells to Epithelial Cells
4.4. Evaluation of the D-Limonene Mechanism of Action
4.4.1. Cell Death Profile
4.4.2. Reactive Oxygen Species (ROS) Quantification
4.4.3. Malondialdehyde Assay (MDA)
4.4.4. Mitochondrial Membrane Potential Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Netea, M.G.; Joosten, L.A.; Van der Meer, J.W.; Kullberg, B.J.; Van de Veerdonk, F.L. Immune defence against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Govender, N.P.; Patel, J.; Magobo, R.E.; Naicker, S.; Wadula, J.; Whitelaw, A.; Coovadia, Y.; Kularatne, R.; Govind, C.; Lockhart, S.R.; et al. Emergence of azole-resistant Candida parapsilosis causing bloodstream infection: Results from laboratory-based sentinel surveillance in South Africa. J. Antimicrob. Chemother. 2016, 71, 1994–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meletiadis, J.; Curfs-Breuker, I.; Meis, J.F.; Mouton, J.W. In vitro antifungal susceptibility testing of Candida isolates with the EUCAST methodology, a new method for ECOFF determination. Antimicrob. Agents Chemother. 2017, 61, e02372-16. [Google Scholar] [CrossRef] [Green Version]
- Tóth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D. Candida parapsilosis: From Genes to the Bedside. Clin. Microbiol. Rev. 2019, 32, e00111-18. [Google Scholar] [CrossRef] [Green Version]
- Thakre, A.; Zore, G.; Kodgire, S.; Kazi, S.; Mulange, S.; Patil, R.; Shelar, A.; Santhakumari, B.; Kulkarni, M.; Kharat, K.; et al. Limonene inhibits Candida albicans growth by inducing apoptosis. Med. Mycol. 2018, 56, 565–578. [Google Scholar] [PubMed]
- Weber, L.D.; Pinto, F.G.S.; Scur, M.C.; Souza, J.G.L.; Costa, W.F.; Leite, C.W. Chemical composition and antimicrobial and antioxidant activity of essential oil and various plant extracts from Prunus myrtifolia. Afr. J. Agric. Res. 2014, 9, 846–853. [Google Scholar]
- Marasinib, B.P.; Baral, P.; Aryal, P.; Ghimire, K.R.; Neupane, S.; Dahal, N.; Singh, A.; Ghimire, L.; Shrestha, K. Evaluation of antibacterial activity of some traditionally used medicinal plants against human pathogenic bacteria. BioMed. Res. Int. 2015, 2015, 265425. [Google Scholar]
- Belletti, N.; Ndagijimana, M.; Sisto, C.; Guerzoni, M.E.; Lanciotti, R.; Gardini, F. Evaluation of the antimicrobial activity of citrus essences on Saccharomyces cerevisiae. J. Agric. Food Chem. 2004, 52, 6932–6938. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Shishodia, S.; Sandur, S.K.; Pandey, M.K.; Sethi, G. Inflammation and câncer: How hot is the link? Biochem. Pharmacol. 2006, 72, 1605–1621. [Google Scholar] [CrossRef]
- Prates, H.T.; Santos, J.P. Óleos essenciais no controle de pragas de grãos armazenados. In Armazenagem de Grãos; Lorini, I., Miike, L.H., Senssel, V.M., Eds.; Instituto Bio Geneziz: Campinas, Brazil, 2002; pp. 443–461. [Google Scholar]
- Rosa, M.S.S.; Mendonça-Filho, R.R.; Bizzo, H.R.; Soares, R.M.A.; Souto-Padrón, T.; Alviano, C.S.; Lopes, A.H.C.S. Antileishmanial activity of a linalool-rich essential oil from Croton cajucara. Antimicrob. Agents Chemother. 2003, 47, 1895–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, J.P.; Fyfe, L.; Smith, H. Plant active components—A resource for antiparasitic agents? Trends Parasitol. 2005, 21, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, J.E.; Rossi, D.C.P.; Jabes, D.L.; Barbosa, D.A.; Cunha, F.F.M.; Nunes, L.R.; Arruda, D.C.; Pelleschi Taborda, C. Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp. J. Fungi 2020, 6, 183. [Google Scholar] [CrossRef] [PubMed]
- Ahmedi, S.; Pant, P.; Raj, N.; Manzoor, N. Limonene inhibits virulence associated traits in Candida albicans: In-vitro and in-silico studies. Phytomedicine 2022, 2, 100285. [Google Scholar] [CrossRef]
- Ravichandran, C.; Badgujar, P.C.; Gundev, P.; Upadhyay, A. Review of toxicological assessment of d-limonene, a food and cosmetics additive. Food Chem. Toxicol. 2018, 120, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Chiotos, K.; Vendetti, N.; Zaoutis, T.E.; Baddley, J.; Ostrosky-Zeichner, L.; Pappas, P.; Fisher, B.T. Comparative effectiveness of echinocandins versus fluconazole therapy for the treatment of adult candidaemia due to Candida parapsilosis: A retrospective observational cohort study of the Mycoses Study Group (MSG-12). J. Antimicrob. Chemother. 2016, 71, 3536–3539. [Google Scholar] [CrossRef] [Green Version]
- Kontoyiannis, D.P.; Bassetti, M.; Nucci, M.; Capparella, M.R.; Yan, L.J.; Aram, J.; Hogan, P.A. Anidulafungin for the treatment of candidaemia caused by Candida parapsilosis: Analysis of pooled data from six prospective clinical studies. Mycoses 2017, 60, 663–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilhante, R.S.; de Jesus, S.R.T.; de Souza, C.M.C.B.D.; Teixeira, C.E.; Brito, M.R.; Bandeira, S.P.; Alencar, L.P.; Monteiro, A.J.; Cordeiro, R.S.; Bandeira, T.J.P.G.; et al. Antifungal susceptibility and virulence attributes of animal-derived isolates of Candida parapsilosis complex. J. Med. Microbiol. 2014, 63, 1568–1572. [Google Scholar] [CrossRef]
- Ning, Y.; Xiao, M.; Perlin, D.S.; Zhao, Y.; Lu, M.; Li, I.Y.; Luo, Z.; Dai, R.; Li, S.; Xu, J.; et al. Decreased echinocandin susceptibility in Candida parapsilosis causing candidemia and emergence of a pan-echinocandin resistant case in China. Emerg. Microbes Infect. 2022. [Google Scholar] [CrossRef]
- Mahzoon, T.; Koraei, P.; Tavakol, O.; Gholami, M.; Barfi, A. Effect of Limonene, Camphor and Menthol on Cariogenic Oral Pathogens. PJMHS 2022, 16, 706–709. [Google Scholar] [CrossRef]
- Lamoth, F.; Lockhart, S.R.; Berkow, E.L.; Calandra, T. Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemother. 2018, 1 (Suppl. S1), i4–i13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Rocha, W.P.; Lemos, V.L.B.; Ferreira, M.R.A.; Soares, L.A.L.; Svidzisnki, T.I.E.; Milan, E.P.; Chaves, G.M. Effect of the crude extract of Eugenia uniflora in morphogenesis and secretion of hydrolytic enzymes in Candida albicans from the oral cavity of kidney transplant recipientes. BMC Complement Altern. Med. 2015, 15, 6. [Google Scholar] [CrossRef]
- Zambonelli, A.; D’aurelio, A.Z.; Bianchi, A.; Albasin, A. Effects of essential oils on phytopathogenic fungi in vitro. J. Phytopathol. 1996, 144, 491–494. [Google Scholar] [CrossRef]
- De Billerbeck, V.G.; Roques, C.G.; Bessière, J.; Fonvieille J-Dargent, R. Effects of Cymbopogon nardus (L.) W. Watson essential oil on the growth and morphogenesis of Aspergillus niger. Can. J. Microbiol. 2001, 47, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, Y.; Du, G.; Zhou, J.; Chen, J. Response of Saccharomyces cerevisiae to D-limonene-induced oxidative stress. Appl. Microbiol. Biotechnol. 2013, 97, 6467–6475. [Google Scholar] [CrossRef]
- Brennan, T.C.R.; Kromer, J.O.; Nielsen, L.K. Physiological and transcriptional responses of Saccharomyces cerevisiae to D-Limonene show changes to the cell wall but not to the plasma membrane. Appl. Environ. Microbiol. 2014, 79, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Modrzewska, B.; Kurnatowski, P.; Modrezewka, B.; Kurnatowski, P. Adherence of Candida sp. to host tissues and cells as one of its pathogenicity features. Ann. Parasitol. 2015, 61, 3–9. [Google Scholar]
- Leite-Andrade, M.C.; Soares-Oliveira, M.A.; Santos, F.A.G.; Ximenes Vilela, P.B.; Silva, M.N.; Macêdo, D.P.C.; Neto, R.G.L.; Neves, H.J.P.; Brandão, I.S.L.; Chaves, G.M.; et al. A new approach by optical coherence tomography for elucidating biofilm formation by emergent Candida species. PLoS ONE 2017, 12, e0188020. [Google Scholar] [CrossRef] [Green Version]
- Thaweboon, B.; Thaweboon, S. Effect of Phyllanthus emblica Linn. On Candida adhesion to oral epithelium and denture acrylic. Asian Pac. J. Trop Med. 2011, 4, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.B.F.C.; Silva-Rocha, W.P.; Ferreira, M.R.A.; Soares, L.A.L.; Svidzinski, T.I.E.; Milan, E.P.; Pires, R.H.; Almeida, A.M.F.; Mendes-Giannini, M.J.S.; Chaves, G.M. Influence of Eugenia uniflora Extract on Adhesion to Human Buccal Epithelial Cells, Biofilm Formation, and Cell Surface Hydrophobicity of Candida spp. from the Oral Cavity of Kidney Transplant Recipients. Molecules 2018, 23, 2418. [Google Scholar] [CrossRef] [Green Version]
- Sandes, J.M.; Fontes, A.; Regis-da-Silva, C.G.; Castro, M.C.A.B.; Lima-Junior, C.G.; Silva, F.P.L.; Vasconcellos, M.L.A.A.; Figueiredo, R.C.B.Q. Trypanosoma cruzi Cell Death Induced by the Morita-Baylis-Hillman Adduct 3-Hydroxy-2 Methylene-3(4-Nitrophenylpropanenitrile). PLoS ONE 2014, 9, e93936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, F.; Lewis, R.E.; Kontoyiannis, D.P. Micafungin induced apoptosis in Candida parapsilosis independent of its susceptibility to micafungin. Microbial. Cell 2015, 2, 445–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosek, J.; Holesova, Z.; Kosa, P.; Gacser, A.; Tomaska, L. Biology and genetics of the pathogenic yeast Candida parapsilosis. Curr. Genet. 2009, 55, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Lee, D.G. Reactive oxygen species modulate itraconazole induced apoptosis via mitochondrial disruption in Candida albicans. Free Radic. Res. 2018, 52, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Jarmuszkiewicz, W.; Milani, G.; Fortes, F.; Schreiber, A.Z.; Sluse, F.E.; Vercesi, A.E. First evidence and characterization of an uncoupling protein in fungi kingdom: CpUCP of Candida parapsilosis. FEBS Lett. 2000, 467, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeast; Approved Standard—Third Edition; CLSI Document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Fourth Informational Supplement; CLSI Document M27-S4; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Chaves, G.M.; Bates, S.; Maccallum, D.M.; Odds, F.C. Candida albicans GRX2, encoding a putative glutaredoxin, is required for virulence in a murine model. Genetic Mol. Res. 2007, 6, 1051–1063. [Google Scholar]
- Kearns, M.J.; Davies, P.; Smith, H. Variability of the adherence of Candida albicans strains to human buccal epithelial cells: Inconsistency of differences between strains related to virulence. Sabouraudia 1983, 21, 93–98. [Google Scholar] [CrossRef]
- Bates, D.D.; Mims, J.W. Invasive fungal sinusitis caused by Pseudallescheria boydii: Case report and literature review. Ear. Nose Throat. J. 2006, 85, 729–737. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | AmB (µg/mL) | FLU (µg/mL) | CFG (µg/mL) | AFG (µg/mL) | MFG (µg/mL) | D-Limonene (µg/mL) |
|---|---|---|---|---|---|---|
| URM6338 | 1 | 1 | 1 | 4 | 2 | 512 |
| URM6365 | 0.25 | 0.25 | 0.125 | 8 | 1 | 1024 |
| URM6387 | 1 | 0.25 | 1 | 4 | 2 | 1024 |
| URM 6404 | 1 | 0.5 | 0.25 | 2 | 0.125 | 256 |
| URM6405 | 0.125 | 0.25 | 0.5 | 2 | 0.125 | 512 |
| URM6406 | 0.5 | 0.125 | 2 | 2 | 2 | 512 |
| URM6407 | 1 | 0.5 | 1 | 4 | 1 | 1024 |
| URM6408 | 0.06 | 0.25 | 1 | 2 | 0.25 | 256 |
| URM6409 | 0.5 | 0.125 | 2 | 2 | 2 | 512 |
| URM6410 | 1 | 0.25 | 2 | 4 | 1 | 1024 |
| URM6411 | 0.5 | 1 | 0.5 | 2 | 0.25 | 256 |
| URM6412 | 0.125 | 0.125 | 4 | 4 | 2 | >1024 |
| URM6905 | 1 | 0.5 | 0.25 | 84 | 4 | 512 |
| URM6939 | 1 | 0.5 | 2 | 4 | 0.06 | >1024 |
| URM6944 | 0.06 | 0.25 | 0.5 | 2 | 0.5 | 512 |
| URM6948 | 0.25 | 1 | 4 | 4 | 0.5 | >1024 |
| URM 7087 | 0.5 | 0.5 | 0.25 | 2 | 0.125 | 256 |
| URM 7421 | 1 | 0.25 | 0.25 | 8 | 4 | 512 |
| URM7423 | 0.5 | 0.25 | 2 | 8 | 4 | >1024 |
| URM7425 | 0.125 | 0.25 | 2 | 8 | 2 | 1024 |
| URM 7426 | 0.5 | 0.5 | 1 | 8 | 4 | 1024 |
| URM 7427 | 0.125 | 1 | 0.125 | 4 | 1 | 512 |
| URM7428 | 1 | 0.25 | 2 | 8 | 4 | 1024 |
| URM 7429 | 0.25 | 0.25 | 0.25 | 4 | 2 | 1024 |
| URM 7430 | 0.5 | 0.25 | 1 | 8 | 2 | 256 |
| URM 7431 | 0.5 | 0.25 | 0.25 | 2 | 0.25 | 1024 |
| URM7432 | 1 | 0.06 | 2 | 4 | 1 | 256 |
| URM 7433 | 0.25 | 0.25 | 0.125 | 8 | 1 | 1024 |
| URM 7434 | 0.125 | 1 | 0.125 | 4 | 1 | 512 |
| URM7443 | 0.25 | 0.5 | 4 | 4 | 0.5 | 512 |
| URM7444 | 0.06 | 0.125 | 0.25 | 2 | 0.5 | >1024 |
| URM 7445 | 0.5 | 1 | 0.125 | 4 | 1 | 256 |
| URM 7446 | 0.5 | 1 | 1 | 8 | 4 | 1024 |
| URM7447 | 1 | 0.06 | 2 | 4 | 1 | 512 |
| URM7448 | 0.25 | 4 | 4 | 8 | 2 | 256 |
| URM7449 | 0.5 | 0.5 | 2 | 8 | 2 | 1024 |
| URM7450 | 0.25 | 0.06 | 2 | 4 | 0.5 | >1024 |
| HAM17 | 0.125 | 0.5 | 4 | 4 | 0.5 | 1024 |
| MM12199 | 1 | 0.25 | 1 | 8 | 2 | >1024 |
| HAM26 | 0.125 | 0.25 | 2 | 8 | 0.5 | 512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite-Andrade, M.C.; de Araújo Neto, L.N.; Buonafina-Paz, M.D.S.; de Assis Graciano dos Santos, F.; da Silva Alves, A.I.; de Castro, M.C.A.B.; Mori, E.; de Lacerda, B.C.G.V.; Araújo, I.M.; Coutinho, H.D.M.; et al. Antifungal Effect and Inhibition of the Virulence Mechanism of D-Limonene against Candida parapsilosis. Molecules 2022, 27, 8884. https://doi.org/10.3390/molecules27248884
Leite-Andrade MC, de Araújo Neto LN, Buonafina-Paz MDS, de Assis Graciano dos Santos F, da Silva Alves AI, de Castro MCAB, Mori E, de Lacerda BCGV, Araújo IM, Coutinho HDM, et al. Antifungal Effect and Inhibition of the Virulence Mechanism of D-Limonene against Candida parapsilosis. Molecules. 2022; 27(24):8884. https://doi.org/10.3390/molecules27248884
Chicago/Turabian StyleLeite-Andrade, Melyna Chaves, Luiz Nascimento de Araújo Neto, Maria Daniela Silva Buonafina-Paz, Franz de Assis Graciano dos Santos, Adryelle Idalina da Silva Alves, Maria Carolina Accioly Brelaz de Castro, Edna Mori, Bruna Caroline Gonçalves Vasconcelos de Lacerda, Isaac Moura Araújo, Henrique Douglas Melo Coutinho, and et al. 2022. "Antifungal Effect and Inhibition of the Virulence Mechanism of D-Limonene against Candida parapsilosis" Molecules 27, no. 24: 8884. https://doi.org/10.3390/molecules27248884
APA StyleLeite-Andrade, M. C., de Araújo Neto, L. N., Buonafina-Paz, M. D. S., de Assis Graciano dos Santos, F., da Silva Alves, A. I., de Castro, M. C. A. B., Mori, E., de Lacerda, B. C. G. V., Araújo, I. M., Coutinho, H. D. M., Kowalska, G., Kowalski, R., Baj, T., & Neves, R. P. (2022). Antifungal Effect and Inhibition of the Virulence Mechanism of D-Limonene against Candida parapsilosis. Molecules, 27(24), 8884. https://doi.org/10.3390/molecules27248884