
LC-HR/MS Analysis of Lipophilic Extracts from Calendula arvensis (Vaill.) L. Organs: An Unexplored Source in Cosmeceuticals


 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
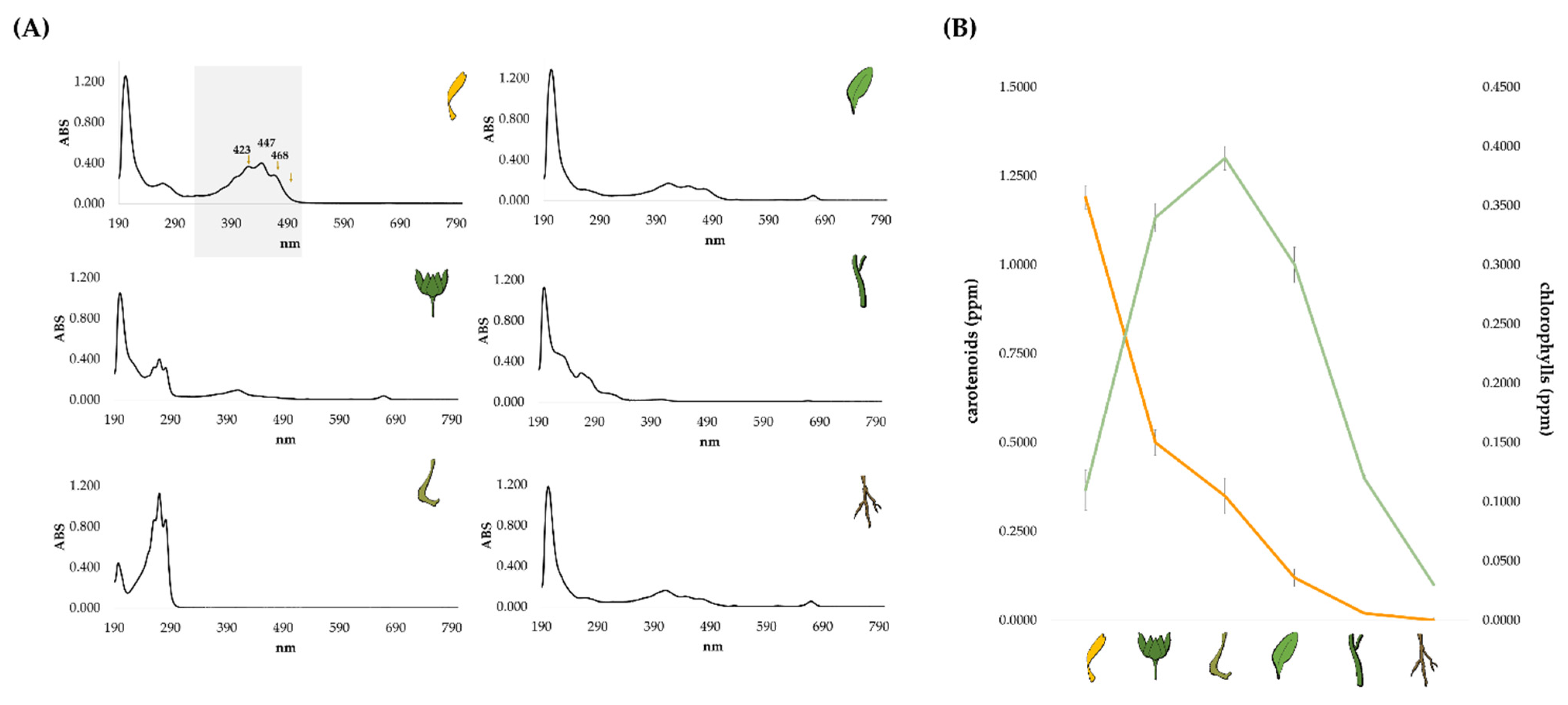
2.1. Lipid Profile of the Different Calendula arvensis Organs
2.2. Cytotoxicity Screening of Apolar Extracts from Calendula arvensis Organs
2.3. Cytotoxicity of Cosmeceutical Formulations Based on Calendula arvensis Apolar Extracts
3. Materials and Methods
3.1. Plant Collection and Extraction
3.2. UHPLC-ESI-QqTOF-MS/MS Analyses
3.3. HPLC-UV-DAD Analyses
3.4. Chlorophyll and Carotenoids Content
3.5. Aqueous Serum Formulation
3.6. Cell Culture and Cytotoxicity Assessment
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Piccolella, S.; Crescente, G.; Pacifico, F.; Pacifico, S. Wild aromatic plants bioactivity: A function of their (poly)phenol seasonality? A case study from Mediterranean area. Phytochem. Rev. 2018, 17, 785–799. [Google Scholar] [CrossRef]
- Alcalde, M.T. Natural and ecological cosmetics. Offarm 2008, 27, 96–104. (In Spanish) [Google Scholar]
- Burnett, C.L.; Fiume, M.M.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.; Marks, J.G., Jr.; Shank, R.C.; Slaga, T.J.; et al. Safety Assessment of Plant-Derived Fatty Acid Oils. Int. J. Toxicol. 2017, 36, 51S–129S. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vioque, R.; Herraiz-Peñalver, D.; Melero Bravo, E.; Ortiz de Elguea-Culebras, G.; Herrero, B.; Santiago, Y.; Bueno, M.; Pérez-Magarino, S.; del Carmen Asensio, S.; Manzanera, M. Variability of the essential oil composition of cultivated populations of Salvia lavandulifolia Vahl. Crop Sci. 2022, 62, 744–752. [Google Scholar] [CrossRef]
- Aburjai, T.; Natsheh, F.M. Plant Used in Cosmetics. Phytother. Res. 2003, 17, 987–1000. [Google Scholar] [CrossRef]
- Gilchrest, B.A.; Krutmann, J. Skin Aging, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 33–42. [Google Scholar]
- Bouftira, I.; Abdelly, C.; Sfar, S. Characterization of cosmetic cream with Mesembryanthemum crystallinum plant extract: Influence of formulation composition on physical stability and antioxidant activity. Int. J. Cosmet. Sci. 2008, 30, 443–452. [Google Scholar] [CrossRef]
- Auner, B.G.; Wirth, M.; Valenta, C. Antioxidative activity and cytotoxicity of four different flavonoids for dermal applications. J. Drug Deliv. Sci. Technol. 2005, 15, 227–232. [Google Scholar] [CrossRef]
- Siti Zulaikha, R.; Sharifah Norkhadijah, S.I.; Praveena, S.M. Hazardous Ingredients in Cosmetics and Personal Care Products and Health Concern: A Review. J. Public Health Res. 2015, 5, 7–15. [Google Scholar] [CrossRef]
- Draelos, Z.D. The Multifunctional Value of Sunscreen containing Cosmetics. Skin Ther. Lett. 2011, 16, 1–3. [Google Scholar]
- Spector, A.A.; Yorek, M.A. Membrane lipid composition and cellular function. J. Lipid Res. 1985, 26, 1015–1035. [Google Scholar] [CrossRef]
- Nicolaou, A. Eicosanoids in skin inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Wertz, P.W. Lipids and barrier function of the skin. Acta Derm. Venereol. Suppl. 2000, 208, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Bialek, A.; Bialek, M.; Jelinska, M.; Tokarz, A. Fatty acid profile of new promising unconventional plant oils for cosmetic use. Int. J. Cosmet. Sci. 2016, 38, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Tremblay, A.; Morin, S.; Martin, C.; Julien, P.; Fardette, J.; Flamand, N.; Pouliot, R. α-Linolenic acid and linoleic acid modulate the lipidome and the skin barrier of a tissue-engineered skin model. Acta Biomater. 2022, 140, 261–274. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Carotenoids and protection against solar UV radiation. Skin Pharmacol. Physiol. 2002, 15, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Darvin, M.; Patzelt, A.; Gehse, S.; Schanzer, S.; Benderoth, C.; Sterry, W.; Lademann, J. Cutaneous concentration of lycopene correlates significantly with the roughness of the skin. Eur. J. Pharm. Biopharm. 2008, 69, 943–947. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Marini, A.; Jaenicke, T.; Stahl, W.; Krutmann, J. Molecular evidence that oral supplementation with lycopene or lutein protects human skin against ultraviolet radiation: Results from a double-blinded, placebo-controlled, crossover study. Br. J. Dermatol. 2017, 176, 1231–1240. [Google Scholar] [CrossRef] [Green Version]
- Davinelli, S.; Nielsen, M.E.; Scapagnini, G. Astaxanthin in skin health, repair, and disease: A comprehensive review. Nutrients 2018, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Stinca, A.; Musarella, C.M.; Rosati, L.; Laface, V.L.A.; Licht, W.; Fanfarillo, E.; Wagensommer, R.P.; Galasso, G.; Fascetti, S.; Esposito, A.; et al. Italian Vascular Flora: New Findings, Updates and Exploration of Floristic Similarities between Regions. Diversity 2021, 13, 600. [Google Scholar] [CrossRef]
- Fiorentino, M.; Gravina, C.; Piccolella, S.; Pecoraro, M.T.; Formato, M.; Stinca, A.; Pacifico, S.; Esposito, A. Calendula arvensis (Vaill.) L.: A Systematic Plant Analysis of the Polar Extracts from Its Organs by UHPLC-HRMS. Foods 2022, 11, 247. [Google Scholar] [CrossRef]
- Messina, C.M.; Troia, A.; Arena, R.; Manuguerra, S.; Ioannou, T.; Curcuraci, E.; Renda, G.; Hellio, C.; Santulli, A. Species-Specific Antioxidant Power and Bioactive Properties of the Extracts Obtained from Wild Mediterranean Calendula Spp. (Asteraceae). Appl. Sci. 2019, 9, 4627. [Google Scholar] [CrossRef] [Green Version]
- Chakraborthy, G.S. Antimicrobial activity of the leaf extracts of Calendula officinalis (Linn.). J. Herb. Med. Toxicol. 2008, 2, 65–66. [Google Scholar]
- Arora, D.; Rani, A.; Sharma, A. A review on phytochemistry and ethnopharmacological aspects of genus Calendula. Pharmacogn. Rev. 2013, 7, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Passalacqua, N.G.; Guarrera, P.M.; De Fine, G. Contribution to the Knowledge of the Folk Plant Medicine in Calabria Region (Southern Italy). Fitoterapia 2007, 78, 52–68. [Google Scholar] [CrossRef]
- Addis, R.; Cruciani, S.; Santaniello, S.; Bellu, E.; Sarais, G.; Ventura, C.; Maioli, M.; Pintore, G. Fibroblast Proliferation and Migration in Wound Healing by Phytochemicals: Evidence for a Novel Synergic Outcome. Int. J. Med. Sci. 2020, 17, 1030–1042. [Google Scholar] [CrossRef] [Green Version]
- Abudunia, A.M.; Marmouzi, I.; Faouzi, M.E.A.; Ramli, Y.; Taoufik, J.; El Madani, N.; Essassi, E.M.; Salama, A.; Khedid, K.; Ansar, M.; et al. Anticandidal, Antibacterial, Cytotoxic and Antioxidant Activities of Calendula arvensis Flowers. J. Mycol. Med. 2017, 27, 90–97. [Google Scholar] [CrossRef]
- Abutaha, N.; Nasr, F.A.; Mohammed, A.Z.; Semlali, A.; Al-Mekhlafi, F.A.; Wadaan, M.A. Calendula arvensis L. as an Anti-Cancer Agent against Breast Cancer Cell Lines. Mol. Biol. Rep. 2019, 46, 2187–2196. [Google Scholar] [CrossRef]
- Ul’ Chenko, N.T.; Glushenkova, A.I.; Mukhamedova, K.S. Lipids of Calendula officinalis. Chem. Nat. Compd. 1998, 34, 272–274. [Google Scholar] [CrossRef]
- Dulf, F.V.; Pamfil, D.; Baciu, A.D.; Pintea, A. Fatty acid composition of lipids in pot marigold (Calendula officinalis L.) seed genotypes. Chem. Cent. J. 2013, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, M.J.; Hopkins, C.Y. Conjugated fatty acids of Tragopogon and Calendula seed oils. Can. J. Chem. 1960, 38, 2500–2507. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, K.; Hornung, E.; Peitzsch, N.; Renz, A.; Feussner, I. Isolation and characterization of a calendic acid producing (8,11)-linoleoyl desaturase. FEBS Lett. 1999, 462, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, D.W.; Savile, C.K.; Qiu, X.; Buist, P.H.; Covello, P.S. Mechanism of 1,4- dehydrogenation catalyzed by a fatty acid (1,4)-desaturase of Calendula officinalis. Eur. J. Biochem. 2002, 269, 5024–5029. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, J.L.; Wiens, A.M.; Ericsson, L.H. Identification of fatty acids by electrospray mass spectrometry and tandem mass spectrometry. J. Mass Spectrom. 1996, 31, 184–192. [Google Scholar] [CrossRef]
- Yang, K.; Zhao, Z.; Gross, R.W.; Han, X. Identification and quantitation of unsaturated fatty acid isomers by electrospray ionization tandem mass spectrometry: A shotgun lipidomics approach. Anal. Chem. 2011, 83, 4243–4250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuda, O.; Brezinova, M.; Rombaldova, M.; Slavikova, B.; Posta, M.; Beier, P.; Janovska, P.; Veleba, J.; Kopecky, J., Jr.; Kudova, E.; et al. Docosahexaenoic Acid-Derived Fatty Acid Esters of Hydroxy Fatty Acids (FAHFAs) With Anti-inflammatory Properties. Diabetes 2016, 65, 2580–2590. [Google Scholar] [CrossRef] [Green Version]
- Formato, M.; Piccolella, S.; Zidorn, C.; Vastolo, A.; Calabrò, S.; Cutrignelli, M.I.; Pacifico, S. UHPLC-ESI-QqTOF Analysis and In Vitro Rumen Fermentation for Exploiting Fagus sylvatica Leaf in Ruminant Diet. Molecules 2022, 27, 2217. [Google Scholar] [CrossRef]
- Grati, W.; Samet, S.; Bouzayani, B.; Ayachi, A.; Treilhou, M.; Téné, N.; Mezghani-Jarraya, R. HESI-MS/MS Analysis of Phenolic Compounds from Calendula aegyptiaca Fruits Extracts and Evaluation of Their Antioxidant Activities. Molecules 2022, 27, 2314. [Google Scholar] [CrossRef]
- Arena, K.; Rigano, F.; Mangraviti, D.; Cacciola, F.; Occhiuto, F.; Dugo, L.; Dugo, P.; Mondello, L. Exploration of Rapid Evaporative-Ionization Mass Spectrometry as a Shotgun Approach for the Comprehensive Characterization of Kigelia Africana (Lam) Benth. Fruit. Molecules 2020, 25, 962. [Google Scholar] [CrossRef] [Green Version]
- Vilchez, A.C.; Margutti, M.P.; Reyna, M.; Wilke, N.; Villasuso, A.L. Recovery from chilling modulates the acyl-editing of phosphatidic acid molecular species in barley roots (Hordeum vulgare L.). Plant Physiol. Biochem. 2021, 167, 862–873. [Google Scholar] [CrossRef]
- Testerink, C.; Munnik, T. Molecular, cellular, and physiological responses to phosphatidic acid formation in plants. J. Exp. Bot. 2011, 62, 2349–2361. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Devaiah, S.P.; Zhang, W.; Welti, R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006, 45, 250–278. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Lee, B.D.; Han, J.M.; Kim, J.H.; Kim, Y.; Suh, P.-G.; Ryu, S.H. Phospholipase D2 activity suppresses hydrogen peroxide-induced apoptosis in PC12 cells. J. Neurochem. 2000, 75, 1053–1059. [Google Scholar] [CrossRef] [Green Version]
- Villanueva-Bermejo, D.; Vázquez, E.; Villalva, M.; Santoyo, S.; Fornari, T.; Reglero, G.; Rodriguez García-Risco, M. Simultaneous Supercritical Fluid Extraction of Heather (Calluna vulgaris L.) and Marigold (Calendula officinalis L.) and Anti-Inflammatory Activity of the Extracts. Appl. Sci. 2019, 9, 2245. [Google Scholar] [CrossRef]
- Gudoityte, E.; Arandarcikaite, O.; Mazeikiene, I.; Bendokas, V.; Liobikas, J. Ursolic and Oleanolic Acids: Plant Metabolites with Neuroprotective Potential. Int. J. Mol. Sci. 2021, 22, 4599. [Google Scholar] [CrossRef]
- Jesus, J.A.; Lago, J.H.G.; Laurenti, M.D.; Yamamoto, E.S.; Passero, L.F.D. Antimicrobial Activity of Oleanolic and Ursolic Acids: An Update. Evid. Based Complement. Altern. Med. 2015, 2015, 620472. [Google Scholar] [CrossRef] [Green Version]
- Raal, A.; Kirsipuu, K.; Must, R.; Tenno, S. Content of total carotenoids in Calendula officinalis L. from different countries cultivated in Estonia. Natl. Prod. Comm. 2009, 4, 35–38. [Google Scholar] [CrossRef] [Green Version]
- Khalid, K.A.; Teixeira da Silva, J.A. Biology of Calendula officinalis L.: Focus on Pharmacology, Biological activities and Agronomic Practices. Med. Aromat. Plant Sci. Biotechnol. 2012, 6, 12–27. [Google Scholar]
- Sausserde, R.; Kampuss, K. Composition of carotenoids in Calendula (Calendula officinalis L.) flowers. In Proceedings of the FOODBALT 2014: 9th Baltic Conference on Food Science and Technology “Food for Consumer Well-Being”; LLU: Jelgava, Latvia, 2014; pp. 8–12. [Google Scholar]
- Kishimoto, S.; Maoka, T.; Sumitomo, K.; Ohmiya, A. Analysis of carotenoid composition in petals of calendula (Calenula officinalis L.). Biosc. Biotechnol. Biochem. 2005, 69, 2122–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, S.; Sumitomo, K.; Yagi, M.; Nakayama, M.; Ohmiya, A. Three routes to orange petal color via carotenoids components in 9 Compositae species. J. Jpn. Soc. Hortic. Sci. 2007, 76, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, S.; Ohmiya, A. Studies in carotenoids in the petals of composite plants. J. Jpn. Soc. Hortic. Sci. 2009, 78, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Pintea, A.; Constantin, B.; Andrei, S.; Socaciu, C. HPLC analysis of carotenoids in four varieties of Calendula officinalis L. flowers. Acta Biol. Szeged. 2003, 47, 37–40. [Google Scholar]
- Bakó, E.; Deli, J.; Tóth, G. HPLC study on the carotenoid composition of Calendula products. J. Biochem. Biophys. Methods 2002, 53, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Chuyen, H.V.; Nguyen, M.H.; Roach, P.D.; Golding, J.B.; Parks, S.E. Microwave-assisted extraction and ultrasound-assisted extraction for recovering carotenoids from Gac peel and their effects on antioxidant capacity of the extracts. Food Sci. Nutr. 2017, 6, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-deLeón, E.; Bah, M.; Jiménez-Halla, J.O.C.; Bonilla-Cruz, J.; Estévez, M.; Báez, J.E. Synthesis and characterization of segmented poly (ester-urethane)s (PEUs) containing carotenoids. Polym. Chem. 2019, 10, 6580. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Matozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Fouché, M.; Willers, C.; Hamman, S.; Malhere, C.; Steenekamp, J. Wound Healing Effects of Aloe muth-muth: In Vitro Investigations Using Immortalized Human Keratinocytes (HaCaT). Biology 2020, 9, 350. [Google Scholar] [CrossRef]
- Razak, N.A.; Abu, N.; Ho, W.Y.; Zamberi, N.R.; Tan, S.W.; Alitheen, N.B.; Long, K.; Yeap, S.K. Cytotoxicity of eupatorin in MCF-7 and MDA-MB-231 human breast cancer cells via cell cycle arrest, anti-angiogenesis and induction of apoptosis. Sci. Rep. 2019, 9, 1514. [Google Scholar] [CrossRef]
- Vale, N.; Silva, S.; Duarte, D.; Crista, D.M.A.; da Silva, L.P.; Joaquim, C.G.; da Silva, E. Normal breast epithelial MCF-10A cells to evaluate the safety of carbon dots. RSC Med. Chem. 2021, 12, 245. [Google Scholar] [CrossRef]
- Khanna, S.; Darbre, P.D. Parabens enable suspension growth of MCF-10A immortalized, non-transformed human breast epithelial cells. J. Appl. Toxicol. 2013, 33, 378–382. [Google Scholar] [CrossRef]
- Farasani, A.; Darbre, P.D. Exposure to cyclic volatile methylsiloxanes (cVMS) causes anchorage-independent growth and reduction of BRCA1 in non-transformed human breast epithelial cells. J. Appl. Toxicol. 2017, 37, 454–461. [Google Scholar] [CrossRef]
- Wei, Z.; Zhou, T.; Xia, Z.; Liu, S.; Li, M.; Zhang, G.; Tian, Y.; Li, B.; Wang, L.; Liu, S. Absolute Configuration and Biological Evaluation of Novel Triterpenes as Possible Anti-Inflammatory or Anti-Tumor Agents. Molecules 2022, 27, 6641. [Google Scholar] [CrossRef] [PubMed]
- Stelling-Férez, J.; Gabaldón, J.A.; Nicolás, F.J. Oleanolic acid stimulation of cell migration involves a biphasic signaling mechanism. Sci Rep. 2022, 12, 15065. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zeng, X.; Wu, X. Effect of Oleanolic Acid on Apoptosis and Autophagy of SMMC-7721 Hepatoma Cells. Med. Sci. Monit. 2020, 26, e921606. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bishayee, K.; Khuda-Bukhsh, A.R. Oleanolic acid isolated from ethanolic extract of Phytolacca decandra induces apoptosis in A375 skin melanoma cells: Drug-DNA interaction and signaling cascade. J. Integr. Med. 2014, 12, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Neimkhum, W.; Anuchapreeda, S.; Lin, W.-C.; Lue, S.-C.; Lee, K.-H.; Chaiyana, W. Effects of Carissa carandas Linn. Fruit, Pulp, Leaf, and Seed on Oxidation, Inflammation, Tyrosinase, Matrix Metalloproteinase, Elastase, and Hyaluronidase Inhibition. Antioxidants 2021, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.K.; Choi, Y.A.; Park, W.; Lee, T.; Ryu, S.H.; Kim, S.Y.; Kim, J.R.; Kim, J.H.; Baek, S.H. Phosphatidic acid regulates systemic inflammatory responses by modulating the Akt-mammalian target of rapamycin-p70 S6 kinase 1 pathway. J. Biol. Chem. 2003, 278, 45117–45127. [Google Scholar] [CrossRef] [Green Version]
- Fluhr, J.W.; Darlenski, R.; Surber, C. Glycerol and the skin: Holistic approach to its origin and functions. Br. J. Dermatol. 2008, 159, 23–34. [Google Scholar] [CrossRef]
- Mayser, P.; Grimm, H.; Grimminger, F. n-3 fatty acids in psoriasis. Br. J. Nutr. 2002, 87, S77–S82. [Google Scholar] [CrossRef] [Green Version]
- Delmastro-Greenwood, M.; Freeman, B.A.; Wendell, S.G. Redox-dependent anti-inflammatory signaling actions of unsaturated fatty acids. Annu. Rev. Physiol. 2014, 76, 79–105. [Google Scholar] [CrossRef] [Green Version]
- Rinkevich, Y.; Matranga, V.; Baruch, R. Stem cells in aquatic invertebrates: Common premises and emerging unique themes. In Stem Cells in Marine Organisms; Rinkevich, B., Matranga, V., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 61–103. [Google Scholar] [CrossRef]
- Re, T.A.; Mooney, D.; Antignac, E.; Dufour, E.; Bark, I.; Srinivasan, V.; Nohynek, G. Application of the threshold of toxicological concern approach for the safety evaluation of calendula flower (Calendula officinalis) petals and extracts used in cosmetic and personal care products. Food Chem. Toxicol. 2009, 47, 1246–1254. [Google Scholar] [CrossRef]
- Kodiyan, J.; Amber, K.T. A Review of the Use of Topical Calendula in the Prevention and Treatment of Radiotherapy-Induced Skin Reactions. Antioxidants 2015, 4, 293–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnuqaydan, A.M.; Lenehan, C.E.; Hughes, R.R.; Sanderson, B.J. Calendula officinalis Extracts Protect against H2O2 Induced Chromosome Damage on HacaT Human Skin Cells. J. Carcinog. Mutagen. 2015, 6, 245. [Google Scholar] [CrossRef] [Green Version]
- Alnuqaydan, A.M.; Sanderson, B.J. Calendula officinalis Protection Against Cytotoxicity Effects of Personal Care Products on HaCaT Human Skin Cells. J. Clin. Toxicol. 2016, 6, 316. [Google Scholar] [CrossRef] [Green Version]
- Sasidharan, S.; Joseph, P.; Junise. Formulation and evaluation of fairness serum using polyherbal extracts. Int. J. Pharm. 2014, 4, 105–112. [Google Scholar]
- Balestrin, L.A.; Kreutz, T.; Fachel, F.N.S.; Bidone, J.; Gelsleichter, N.E.; Koester, L.S.; Bassani, V.L.; Braganhol, E.; Dora, C.L.; Teixeira, H.F. Achyrocline satureyoides (Lam.) DC (Asteraceae) Extract-Loaded Nanoemulsions as a Promising Topical Wound Healing Delivery System: In Vitro Assessments in Human Keratinocytes (HaCaT) and HET-CAM Irritant Potential. Pharmaceutics 2021, 13, 1241. [Google Scholar] [CrossRef]
- Lin, Y.H.; Nien, C.J.; Chen, L.G.; Lee, S.Y.; Chang, W.J.; Pan, Y.H.; Hsieh, S.C.; Huang, M. Sapindus mukorossi Seed Oil Changes Tyrosinase Activity of α-MSH-Induced B16F10 Cells Via the Antimelanogenesic Effect of Eicosenoic Acid. Nat. Prod. Comm. 2020, 15, 1–7. [Google Scholar] [CrossRef]
- Duval, R.E.; Harmand, P.O.; Jayat-Vignoles, C.; Cook-Moreau, J.; Pinon, A.; Delage, C.; Simon, A. Differential involvement of mitochondria during ursolic acid-induced apoptotic process in HaCaT and M4Beu cells. Oncol. Rep. 2008, 19, 145–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegemann, S. Patient centric drug product design in modern drug delivery as an opportunity to increase safety and effectiveness. Expert Opin. Drug Deliv. 2018, 15, 619–627. [Google Scholar] [CrossRef]
- Andersen, F.A.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G., Jr.; Shank, R.C.; Slaga, T.J.; Snyder, P.W. Final report of the cosmetic ingredient review expert panel amended safety assessment of Calendula officinalis-derived cosmetic ingredients. Int. J. Toxicol. 2010, 29, 221–243. [Google Scholar] [CrossRef]
- Dumlao, D.S.; Buczynski, M.W.; Norris, P.C.; Harkewicz, R.; Dennis, E.A. High-throughput lipidomic analysis of fatty acid derived eicosanoids and N-acylethanolamines. Biochim. Biophys. Acta 2011, 1811, 724–736. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Aachary, A.A.; Hydamaka, A.; Eskin, N.A.; Eck, P.; Thiyam-Holländer, U. Reduction of chlorophyll in cold-pressed hemp (Cannabis sativa) seed oil by ultrasonic bleaching and enhancement of oxidative stability. Eur. J. Lipid Sci. Technol. 2018, 120, 1700349. [Google Scholar] [CrossRef]
- Aladić, K.; Jarni, K.; Barbir, T.; Vidović, S.; Vladić, J.; Bilić, M.; Jokić, S. Supercritical CO2 extraction of hemp (Cannabis sativa L.) seed oil. Ind. Crops Prod. 2015, 76, 472–478. [Google Scholar] [CrossRef]
- Piccolella, S.; Bianco, A.; Crescente, G.; Santillo, A.; Chieffi Baccari, G.; Pacifico, S. Recovering Cucurbita pepo cv. ‘Lungo Fiorentino’ Wastes: UHPLC-HRMS/MS metabolic profile, the basis for establishing their nutra- and cosmeceutical valorisation. Molecules 2019, 24, 1479. [Google Scholar] [CrossRef] [Green Version]
- ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Available online: https://biit.cs.ut.ee/clustvis/ (accessed on 20 October 2021).
- Podani, J. SYN-TAX 2000: Computer Programs for Data Analysis in Ecology and Systematics; User’s Manual; Scientia: Budapest, Hungary, 2001; p. 452. [Google Scholar]
- Ohsne, S.; Marques, M.B.; Silveira, P.C.; Válega, M.S.G.A.; Granato, D.; Silva, A.M.S.; Pinto, D.C.G.A. Inter-Individual versus Inter-Population Variability of Calendula suffruticosa subsp. algarbiensis Hexane Extracts. Chem. Biodivers. 2021, 18, e2100120. [Google Scholar] [CrossRef]
- Wiłkomirsko, B.; Kasprzyk, Z. Free and ester-bound triterpene alcohols and sterols in cellular subfractions of Calendula officinalis flowers. Phytochemistry 1979, 18, 253–255. [Google Scholar] [CrossRef]
- Kırmızıbekmez, H.; Bassarello, C.; Piacente, S.; Pizza, C.; Çalı, I. Triterpene Saponins from Calendula arvensis. Z. Naturforsch. 2006, 61, 1170–1173. [Google Scholar] [CrossRef]
- Muley, B.P.; Khadabadi, S.S.; Banarase, N.B. Phytochemical Constituents and Pharmacological Activities of Calendula officinalis Linn (Asteraceae): A Review. Trop. J. Pharm. 2009, 8, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Faustino, M.V.; Seca, A.M.L.; Silveira, P.; Silva, A.M.S.; Pinto, D.C.G.A. Gas chromatography–mass spectrometry profile of four Calendula L. taxa: A comparative analysis. Ind. Crops Prod. 2017, 104, 91–98. [Google Scholar] [CrossRef]
- Ismahene, S.; Seridi, R.; Miguel, C.M.D.M.; Nuria, C. Phytochemical Composition and Evaluation of the Antioxidant Activity of the Ethanolic Extract of Calendula suffruticosa subsp. suffruticosa Vahl. Pharmacogn. J. 2017, 10, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R. Studies of Selected Plant Raw Materials as Alternative Sources of Triterpenes of Oleanolic and Ursolic Acid Types. J. Agric. Food Chem. 2007, 55, 656–662. [Google Scholar] [CrossRef]
- Szakiel, D.; Ruszkowski, D.; Grudniak, A.; Kurek, A.; Wolska, K.I.; Doligalska, M.; Janiszowska, W. Antibacterial and Antiparasitic Activity of Oleanolic Acid and its Glycosides isolated from Marigold (Calendula officinalis). Plant. Med. 2008, 74, 1709–1715. [Google Scholar] [CrossRef] [PubMed]
- Lehbili, M.; Magid, A.A.; Kabouche, A.; Voutquenne-Nazabadioko, L.; Abedini, A.; Morjani, H.; Sarazin, T.; Gangloff, S.C.; Kabouche, Z. Oleanane-type triterpene saponins from Calendula stellata. Phytochemistry 2017, 144, 33–42. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | [M-H]− Calc. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 15 | 9.701 | α-Linolenic acid | C18H30O2 | 277.2177 | 277.2173 | 1.4 | 4.0 | 277.2165 (100); 233.2303 |
| 16 | 9.840 | Calendic acid (1) | C18H30O2 | 277.2177 | 277.2173 | 1.4 | 4.0 | 277.2173 (100) |
| 17 | 10.209 | Calendic acid (2) | C18H30O2 | 277.2177 | 277.2173 | 1.4 | 4.0 | 277.2169 (100) |
| 18 | 10.232 | Palmitoleic acid | C16H30O2 | 253.2177 | 253.2173 | 1.6 | 2.0 | 235.1975;153.2051 (100) |
| 19 | 10.427 | Linoleic acid | C18H32O2 | 279.2336 | 279.2330 | 2.7 | 3.0 | 279.2201 (100); 261.2237 |
| 22 | 10.679 | Linoelaidicic acid | C18H32O2 | 279.2332 | 279.2330 | 0.2 | 3.0 | 279.2201 (100) |
| 24 | 10.872 | Palmitic acid | C16H32O2 | 255.2331 | 255.2330 | 1.0 | 1.0 | 255.2219 (100); 237.2237 |
| 28 | 11.027 | Oleic acid | C18H34O2 | 281.2490 | 281.2486 | 1.4 | 2.0 | 281.2482 (100) |
| 29 | 11.451 | Stearic acid | C18H36O2 | 283.2643 | 283.2643 | 0.2 | 1.0 | 283.2646 (100); 265.2593 |
| Peak | Rt (min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | [M-H]− Calc. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 3 | 7.993 | 13-oxooctadeca-9,11,15-trienoic acid | C18H28O3 | 291.1968 | 291.1966 | 0.8 | 5.0 | 291.1974 (100); 247.2065; 223.1685; 195.1400; 111.0821 |
| 5 | 8.185 | 13-hydroxy-9,11-octadecadienoic acid | C18H32O3 | 295.2284 | 295.2279 | 1.8 | 3.0 | 295.2282 (100); 277.2175; 195.1395; 179.1431; 183.1390 |
| 7 | 8.426 | 9-oxo-10,12-octadecadienoic acid (1) | C18H30O3 | 293.2122 | 293.2123 | 0.3 | 4.0 | 249.2215; 185.1188; 96.9607 (100) |
| 9 | 8.551 | 9-oxo-10,12-octadecadienoic acid (2) | C18H30O3 | 293.2122 | 293.2118 | −1.4 | 4.0 | 197.1177; 185.1177; 96.9553 (100) |
| 10 | 8.879 | 9-hydroxy-10,12-octadecadienoic acid | C18H32O3 | 295.2280 | 295.2279 | 0.4 | 3.0 | 277.2154 (100); 171.1026 |
| Peak | Rt (min) | Tentative Assignment | Formula | [M-H]− Found (m/z) | [M-H]− Calc. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 1 | 7.823 | octadecatrienoyl-sn-glycero-3-phosphate (1) | C21H37O7P | 431.2208 | 431.2204 | 0.9 | 4.0 | 431.2208; 277.2172; 152.9964 (100); 96.9696 |
| 4 | 8.011 | octadecatrienoyl-sn-glycero-3-phosphate (2) | C21H37O7P | 431.2207 | 431.2204 | 0.7 | 4.0 | 431.2206; 277.2171; 152.9964 (100); 96.9696 |
| 6 | 8.424 | octadecadienoyl-sn-glycero-3-phosphate (1) | C21H39O7P | 433.2364 | 433.2361 | 0.8 | 3.0 | 433.2362; 279.2308; 152.9959 (100) |
| 8 | 8.520 | octadecadienoyl-sn-glycero-3-phosphate (2) | C21H39O7P | 433.2368 | 433.2361 | 1.7 | 3.0 | 433.2368; 279.2306; 171.0065; 152.9966 (100) |
| 11 | 9.041 | PI(16:0/18:2) (1) | C43H78O13P | 833.5191 | 833.5186 | 0.7 | 5.0 | 833.5213 (100); 553.2815; 391.2266; 279.2318; 255.2325 |
| 12 | 9.104 | palmitoyl-sn-glycero-3-phosphate | C19H39O7P | 409.2366 | 409.2361 | 1.3 | 1.0 | 409.2363; 255.2337; 152.9963 (100); 96.9702 |
| 20 | 10.620 | PA(18:3/18:3) | C39H65O8P | 691.4349 | 691.4344 | 0.7 | 8.0 | 691.4387; 413.2115; 277.2174 (100); 152.9958 |
| 21 | 10.677 | PI(16:0/18:2) (2) | C43H78O13P | 833.5192 | 833.5186 | 0.8 | 5.0 | 833.5224 (100); 553.2807; 391.2259; 255.2325; 223.0004 |
| 23 | 10.766 | PA(16:0/18:3) (1) | C37H67O8P | 669.4516 | 669.4501 | 2.3 | 5.0 | 669.4531; 409.2344; 391.2271; 277.2171; 255.2328 (100); 152.9953 |
| 25 | 10.889 | PA(16:0/18:3) (2) | C37H67O8P | 669.4520 | 669.4501 | 2.9 | 5.0 | 669.4533; 409.2342; 391.2252; 277.2169; 255.2326 (100); 152.9954 |
| 26 | 11.007 | PA(16:0/18:2) (2) | C37H69O8P | 671.4672 | 671.4657 | 2.2 | 4.0 | 671.4704; 391.2271; 279.2333; 255.2335 (100); 152.9962 |
| 27 | 11.007 | PA(18:2/18:2) | C39H68O8P | 695.4657 | 695.4672 | 2.1 | 6.0 | 695.4682; 433.2357; 415.2255; 279.2326 (100); 152.9952 |
| Peak | Rt (min) | Tentative Assignment | Formula | [M-H]− Calc. (m/z) | [M-H]− Found (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 2 | 7.992 | Acetyl oleanolic acid glucuronide | C38H58O10 | 673.3957 | 673.3976 | 2.8 | 10.0 | 673.3998 (100); 631.3903; 569.3872; 497.3671; 483.3508; 455.3545; 113.0245 |
| 13 | 9.493 | Ursolic acid | C30H48O3 | 455.3536 | 455.3531 | 1.2 | 7.0 | 455.3550 (100) |
| 14 | 9.665 | Oleanolic acid | C30H48O3 | 455.3537 | 455.3531 | 1.4 | 7.0 | 455.3551 (100) |
| FLORETS | BRACTS | STEMS | ROOTS | FRUITS | LEAVES | AERIAL PARTS | Co | Csa | Ca | Css | Cs | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oxo Fatty Acids | ||||||||||||
| 13-hydroxy-9,11-octadecadienoic acid | ● | ● | ● | ● | ● | ● | ||||||
| 9-hydroxy-10,12-octadecadienoic acid | ● | ● | ● | ● | ||||||||
| 13-oxo-9,11,15-octadecatrienoic acid | ● | ● | ● | ● | ● | ● | ||||||
| 9-oxo-10,12-octadecadienoic acid (1) | ● | ● | ● | ● | ● | |||||||
| 9-oxo-10,12-octadecadienoic acid (2) | ● | ● | ● | ● | ● | |||||||
| Phosphatidic Acids | ||||||||||||
| Octadecatrienoyl-sn-glycero-3-phosphate (1) | ● | ● | ||||||||||
| Octadecatrienoyl-sn-glycero-3-phosphate (2) | ● | ● | ● | |||||||||
| Octadecadienoyl-sn-glycero-3-phosphate (1) | ● | ● | ● | ● | ● | ● | ||||||
| Octadecadienoyl-sn-glycero-3-phosphate (2) | ● | ● | ● | ● | ● | ● | ||||||
| PI (16:0/18:2) (1) | ● | ● | ||||||||||
| PI (16:0/18:2) (2) | ● | ● | ● | |||||||||
| Palmitoyl-sn-glycero-3-phosphate | ● | ● | ● | ● | ● | ● | ||||||
| PA (18:3/18:3) | ● | |||||||||||
| PA (16:0/18:3) (1) | ● | ● | ||||||||||
| PA (16:0/18:3) (2) | ● | ● | ||||||||||
| PA (16:0/18:2) | ● | ● | ||||||||||
| PA (18:2/18:2) | ● | ● | ||||||||||
| Saturated Fatty Acids | ||||||||||||
| Palmitic acid | ● | ● | ● | ● | ⦿ | ⦿ | ○ | [29,89,90,91,92] | [88,90] | [27,90] | [93] | |
| Stearic acid | ● | ● | ● | ● | ⦿ | ⦿ | ○ | [28,29,89,91,92,94] | [88,90] | [27,90] | ||
| Mono Unsaturated Fatty Acids | ||||||||||||
| Palmitoleic acid | ● | ● | ● | ● | ⦿ | ⦿ | ○ | [29,89] | [27] | |||
| Oleic acid | ● | ● | ● | ● | ⦿ | ⦿ | ○ | [29,89,91,92] | [27] | |||
| Poly Unsaturated Fatty Acids | ||||||||||||
| α-linolenic acid | ● | ● | ● | ● | ⦿ | ⦿ | ○ | [29,90,91,92] | [88,90] | [27,90] | [93] | |
| Linoleic acid | ● | ● | ● | ● | ⦿ | ⦿ | ○ | [28,29,89,90,91,92] | [88,90] | [27] | [93] | |
| Linoelaidicic acid | ● | ● | ● | ⦿ | ● | ○ | [29] | |||||
| Conjugated Linoleic Acids | ||||||||||||
| Calendic acid (1) | ● | ● | ● | ● | ● | ● | ||||||
| Calendic acid (2) | ● | ● | ○ | ○ | [28,29] | |||||||
| Triterpenes | ||||||||||||
| Hexuronyl acetyl OA | ● | |||||||||||
| Ursolic acid | ● | ● | ● | ● | ● | ○ | ○ | [43,94] | ||||
| Oleanolic acid | ● | ● | ● | ● | ⦿ | ○ | [43,94,95,96] | [97] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gravina, C.; Fiorentino, M.; Formato, M.; Pecoraro, M.T.; Piccolella, S.; Stinca, A.; Pacifico, S.; Esposito, A. LC-HR/MS Analysis of Lipophilic Extracts from Calendula arvensis (Vaill.) L. Organs: An Unexplored Source in Cosmeceuticals. Molecules 2022, 27, 8905. https://doi.org/10.3390/molecules27248905
Gravina C, Fiorentino M, Formato M, Pecoraro MT, Piccolella S, Stinca A, Pacifico S, Esposito A. LC-HR/MS Analysis of Lipophilic Extracts from Calendula arvensis (Vaill.) L. Organs: An Unexplored Source in Cosmeceuticals. Molecules. 2022; 27(24):8905. https://doi.org/10.3390/molecules27248905
Chicago/Turabian StyleGravina, Claudia, Marika Fiorentino, Marialuisa Formato, Maria Tommasina Pecoraro, Simona Piccolella, Adriano Stinca, Severina Pacifico, and Assunta Esposito. 2022. "LC-HR/MS Analysis of Lipophilic Extracts from Calendula arvensis (Vaill.) L. Organs: An Unexplored Source in Cosmeceuticals" Molecules 27, no. 24: 8905. https://doi.org/10.3390/molecules27248905
APA StyleGravina, C., Fiorentino, M., Formato, M., Pecoraro, M. T., Piccolella, S., Stinca, A., Pacifico, S., & Esposito, A. (2022). LC-HR/MS Analysis of Lipophilic Extracts from Calendula arvensis (Vaill.) L. Organs: An Unexplored Source in Cosmeceuticals. Molecules, 27(24), 8905. https://doi.org/10.3390/molecules27248905