

Metabolite Profiling of Gardenia jasminoides Ellis In Vitro Cultures with Different Levels of Differentiation





Abstract
:1. Introduction
2. Results and Discussion
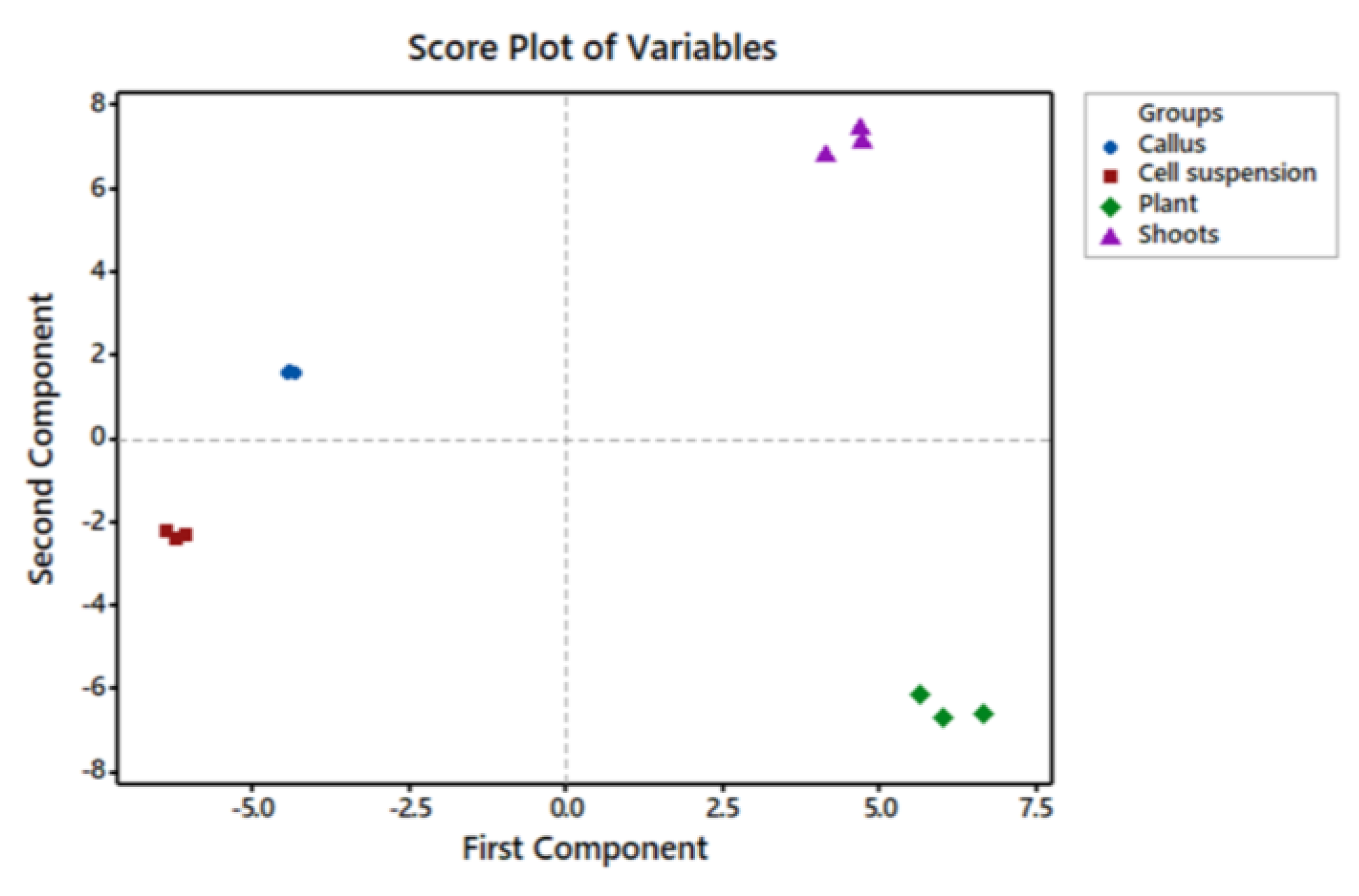
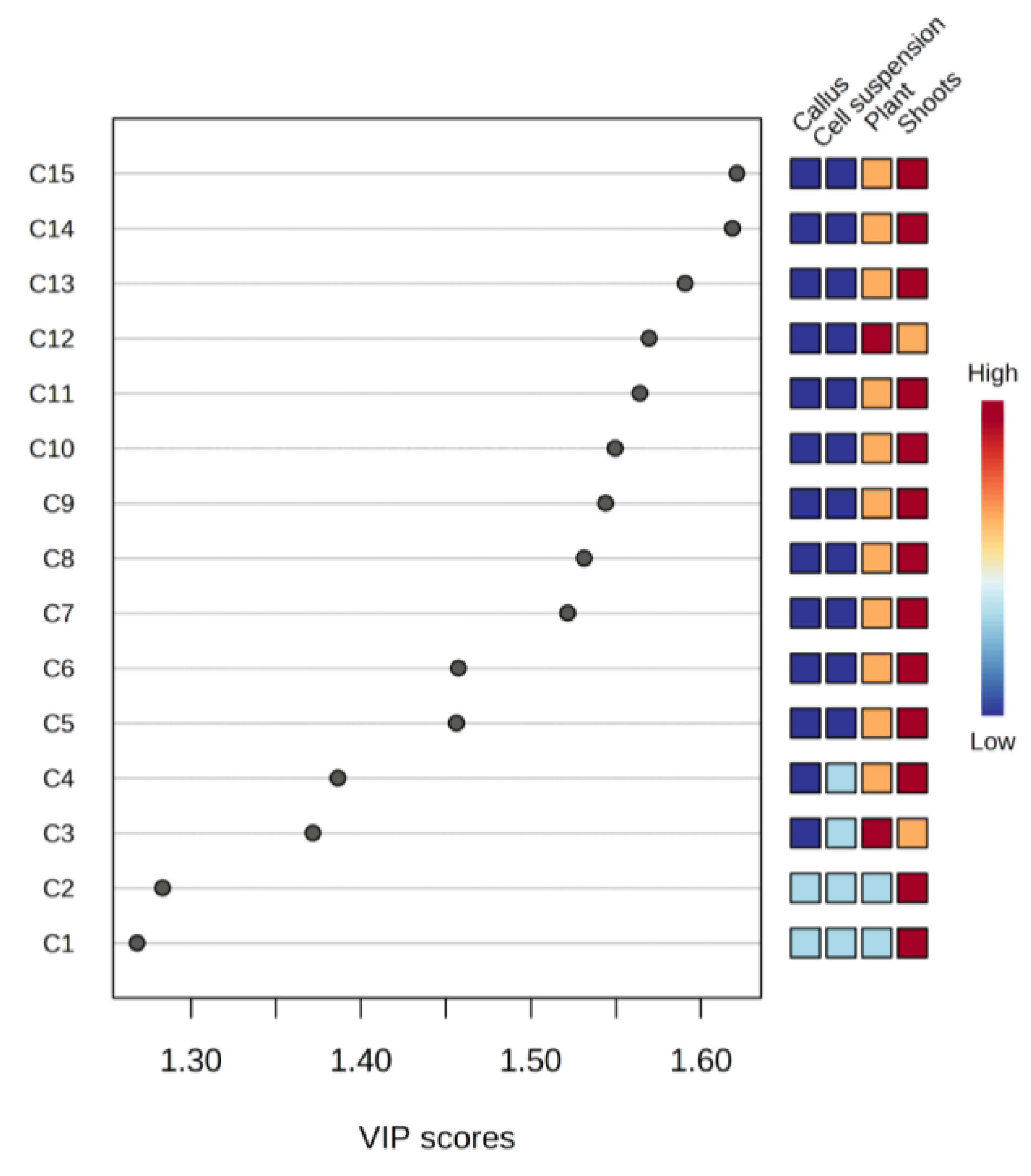
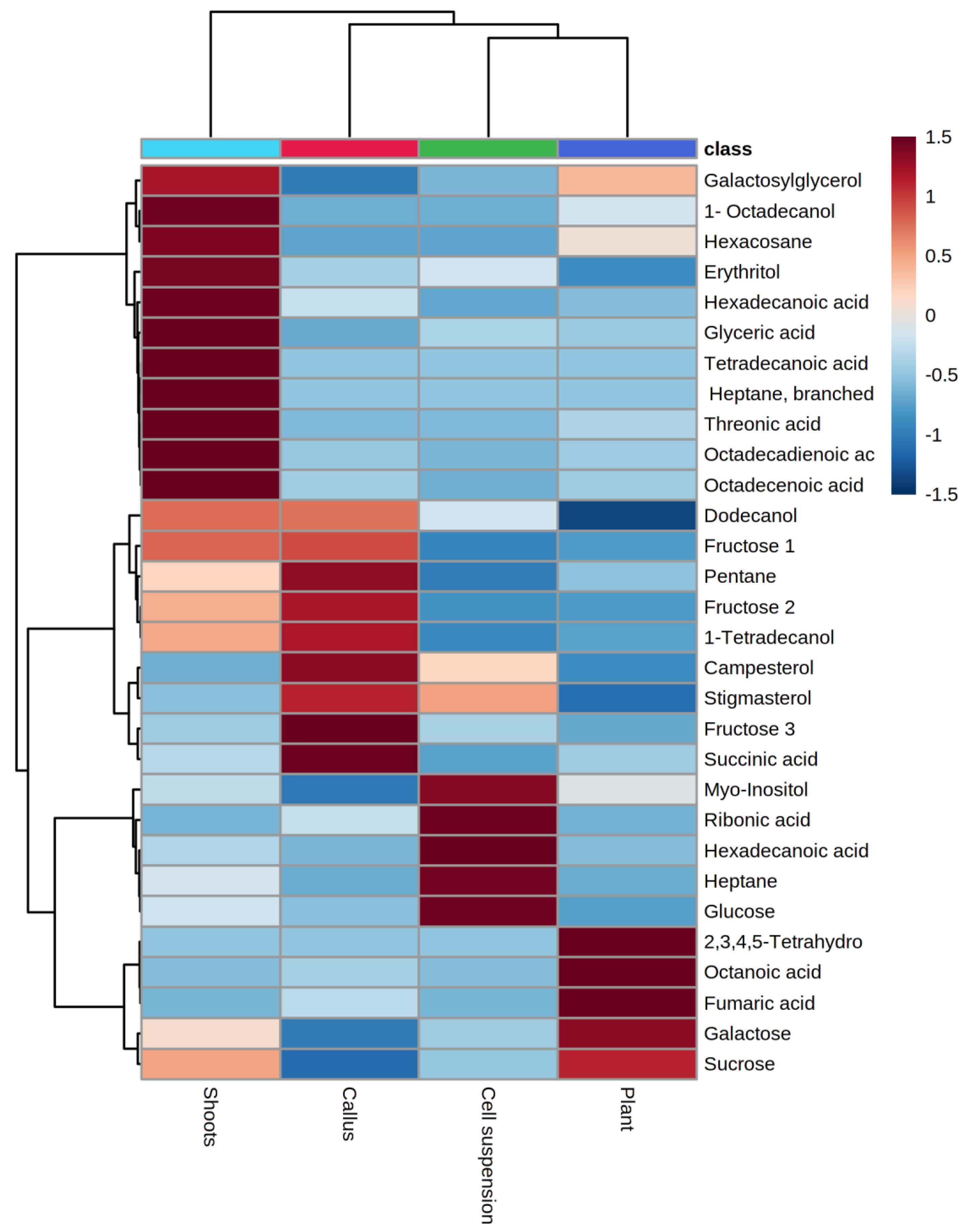
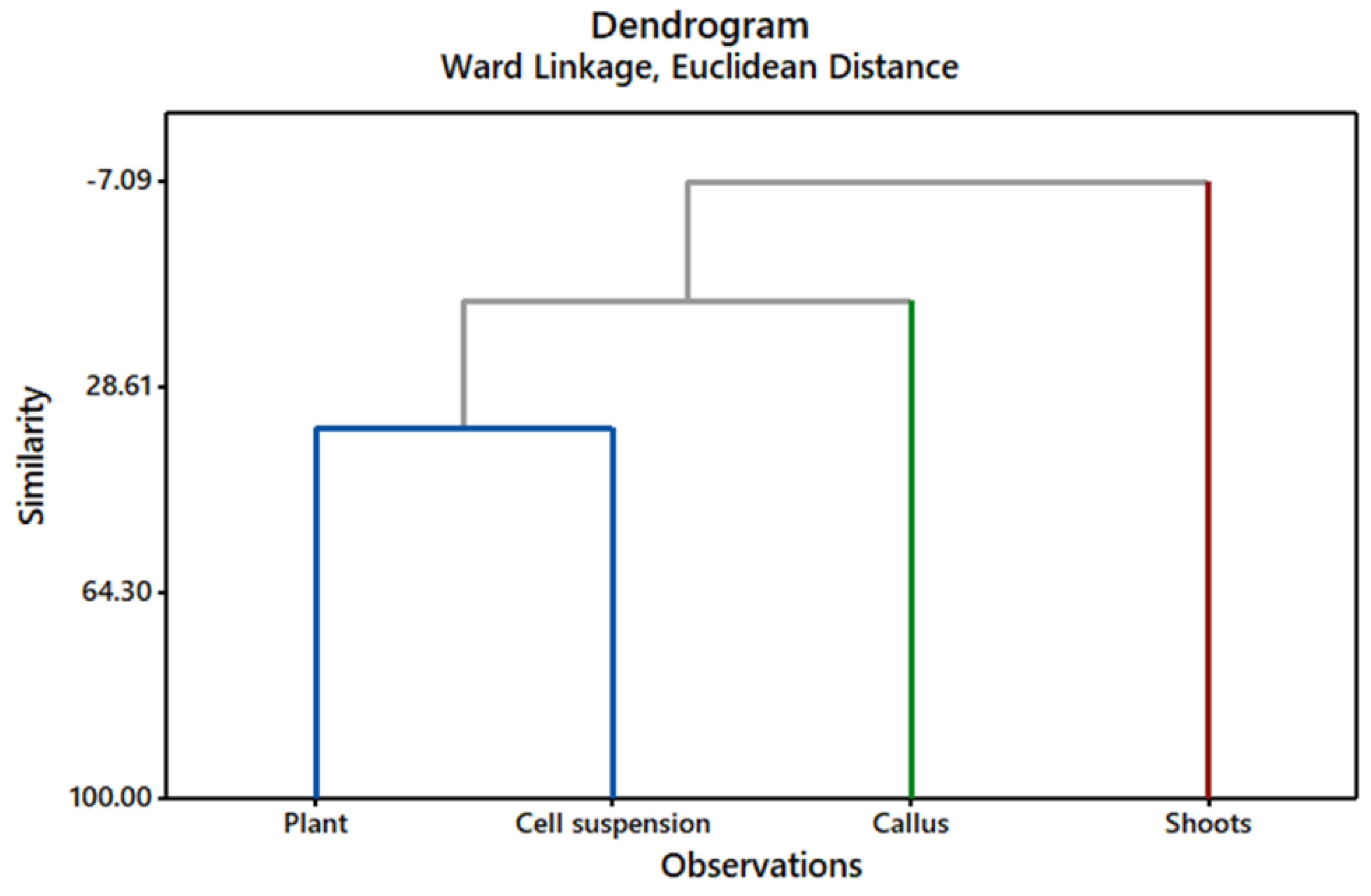
2.1. GC/MS Profiling of Gardenia Plant Leaves and In Vitro Cultures
2.2. HPLC Quantification of Phenolic Compounds in Gardenia Plant Leaves and In Vitro Cultures
2.3. Antioxidant Activities of Phenolic Extracts from Gardenia Plant Leaves and In Vitro Cultures
2.4. Somaclonal Variation in Gardenia In Vitro Cultures
3. Materials and Methods
3.1. Plant Material
3.2. GC/MS Analyses
3.3. HPLC Analyses
3.4. Antioxidant Activity Analyses
3.4.1. DPPH Assay
3.4.2. TEAC Assay
3.4.3. FRAP Assay
3.4.4. CUPRAC Assay
3.4.5. Total Phenolic Assay
3.5. Analyses of Somaclonal Variation
3.5.1. DNA Extraction
3.5.2. SSR Analysis
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Chen, L.; Li, M.; Yang, Z.; Tao, W.; Wang, P.; Tian, X.; Li, X.; Wang, W. Gardenia jasminoides Ellis: Ethnopharmacology, phytochemistry, and pharmacological and industrial applications of an important traditional chinese medicine. J. Ethnopharmacol. 2020, 257, 112829. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Jin, B.; Yan, Y.; Yuan, Q.; Wen, H.; Tao, W.; Cui, X.; Shan, C.; Yu, S. Anti-depressant effect of zhi-zi-chi decoction on cums mice and elucidation of its signaling pathway. J. Ethnopharmacol. 2021, 266, 113283. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Y.-F.; Li, F.; Zhang, H.-Y. Fructus gardenia (Gardenia jasminoides J. Ellis) phytochemistry, pharmacology of cardiovascular, and safety with the perspective of new drugs development. J. Asian Nat. Prod. Res. 2013, 15, 94–110. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Liu, J.-h. Research and application progress of Gardenia jasminoides. Chin. Herb. Med. 2018, 10, 362–370. [Google Scholar] [CrossRef]
- Xiao, W.; Li, S.; Wang, S.; Ho, C.-T. Chemistry and bioactivity of Gardenia jasminoides. J. Food Drug Anal. 2017, 25, 43–61. [Google Scholar] [CrossRef] [Green Version]
- Park, E.-H.; Joo, M.-H.; Kim, S.-H.; Lim, C.-J. Antiangiogenic activity of Gardenia jasminoides fruit. Phytother. Res. 2003, 17, 961–962. [Google Scholar] [CrossRef]
- Wang, L.; Yang, C.; Song, F.; Liu, Z.; Liu, S. Therapeutic effectiveness of Gardenia jasminoides on type 2 diabetic rats: Mass spectrometry-based metabolomics approach. J. Agric. Food Chem. 2020, 68, 9673–9682. [Google Scholar] [CrossRef]
- Ni, Y.; Li, L.; Zhang, W.; Lu, D.; Zang, C.; Zhang, D.; Yu, Y.; Yao, X. Discovery and LC-MS characterization of new crocins in Gardeniae Fructus and their neuroprotective potential. J. Agric. Food Chem. 2017, 65, 2936–2946. [Google Scholar] [CrossRef]
- Ma, W.-W.; Tao, Y.; Wang, Y.-Y.; Peng, I.F. Effects of Gardenia jasminoides extracts on cognition and innate immune response in an adult drosophila model of alzheimer’s disease. Chin. J. Nat. Med. 2017, 15, 899–904. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Park, S.; Sathiyaseelan, A.; Kim, K.-N.; Cho, S.-H.; Mariadoss, A.V.A.; Wang, M.-H. Metabolite Profiling of Methanolic Extract of Gardenia jaminoides by LC-MS/MS and GC-MS and Its Anti-Diabetic, and Anti-Oxidant Activities. Pharmaceuticals 2021, 14, 102. [Google Scholar] [CrossRef]
- Wang, L.; Liu, S.; Zhang, X.; Xing, J.; Liu, Z.; Song, F. A strategy for identification and structural characterization of compounds from Gardenia jasminoides by integrating macroporous resin column chromatography and liquid chromatography-tandem mass spectrometry combined with ion-mobility spectrometry. J. Chromatogr. A 2016, 1452, 47–57. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, X.; Xu, H.; Gong, Y.; Yuan, F.; Gao, Y. On-line HPLC-ABTS screening and HPLC-DAD-MS/MS identification of free radical scavengers in gardenia (Gardenia jasminoides Ellis) fruit extracts. Food Chem. 2010, 123, 521–528. [Google Scholar] [CrossRef]
- Yu, Y.; Feng, X.-L.; Gao, H.; Xie, Z.-L.; Dai, Y.; Huang, X.-J.; Kurihara, H.; Ye, W.-C.; Zhong, Y.; Yao, X.-S. Chemical constituents from the fruits of Gardenia jasminoides Ellis. Fitoterapia 2012, 83, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Yang, L.; Zhao, S.; Chen, L.; Zhao, F.; Qiu, F. Chemical constituents from the fruit of Gardenia jasminoides and their inhibitory effects on nitric oxide production. Bioorg. Med. Chem. Lett. 2013, 23, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-G.; Ren, Y.-J.; Liu, Y.-L.; Wang, M.-N.; He, C.; Chen, X.; Fan, X.-L.; Zhang, Y.-L.; Hao, Z.-Y.; Li, H.-W.; et al. Iridoid glycosides and lignans from the fruits of Gardenia jasminoides Eills. Phytochemistry 2021, 190, 112893. [Google Scholar] [CrossRef] [PubMed]
- Akihisa, T.; Watanabe, K.; Yamamoto, A.; Zhang, J.; Matsumoto, M.; Fukatsu, M. Melanogenesis inhibitory activity of monoterpene glycosides from Gardeniae Fructus. Chem. Biodivers. 2012, 9, 1490–1499. [Google Scholar] [CrossRef]
- Lelono, R.; Tachibana, S.; Itoh, K. Isolation of antifungal compounds from Gardenia jasminoides. Pak. J. Biol. Sci. PJBS 2009, 12, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, R.; Saha, M.R.; Subhan, N.; Hossain, H.; Jahan, I.A.; Akter, R.; Alam, A. HPLC-analysis of polyphenolic compounds in Gardenia jasminoides and determination of antioxidant activity by using free radical scavenging assays. Adv. Pharm. Bull. 2014, 4, 273–281. [Google Scholar]
- Debnath, T.; Park, P.-J.; Deb Nath, N.C.; Samad, N.B.; Park, H.W.; Lim, B.O. Antioxidant activity of Gardenia jasminoides Ellis fruit extracts. Food Chem. 2011, 128, 697–703. [Google Scholar] [CrossRef]
- Liu, Z.-B.; Chen, J.-G.; Yin, Z.-P.; Shangguan, X.-C.; Peng, D.-Y.; Lu, T.; Lin, P. Methyl jasmonate and salicylic acid elicitation increase content and yield of chlorogenic acid and its derivatives in Gardenia jasminoides cell suspension cultures. Plant Cell Tissue Organ Cult. 2018, 134, 79–93. [Google Scholar] [CrossRef]
- Georgiev, V.; Slavov, A.; Vasileva, I.; Pavlov, A. Plant cell culture as emerging technology for production of active cosmetic ingredients. Eng. Life Sci. 2018, 18, 779–798. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, G.; Georgiev, V.; Pavlov, A. Recent applications of plant cell culture technology in cosmetics and foods. Eng. Life Sci. 2021, 21, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Amer, E.; Fetouh, M.; Rasha, S.E. Micropropagation and acclimatization of Gardenia jasminoides Ellis. Environ. Sci. 2019, 14, 107–120. [Google Scholar]
- Salim, S.A.A.-R.; Hamza, S.Y. An efficient protocol for micro propagation of Gardenia jasminoides Ellis. Biosci. Biotechnol. Res. Asia 2017, 14, 757. [Google Scholar] [CrossRef]
- Dumanois, C.; Godin, B.; Bigot, C. Multiplication végétative in vitro de Gardenia jasminoïdes Ellis: In Vitro vegetative multiplication of Gardenia jasminoïdes Ellis. J. Plant Physiol. 1984, 116, 389–407. [Google Scholar] [CrossRef]
- MS Duhoky, M. Micropropagation of gardenia Gardenia jasminoides by using single nodes. Mesop. J. Agric. 2009, 37, 21–33. [Google Scholar]
- Gaber, M.K.; Barakat, A.A. Micropropagation and somatic embryogenesis induction of Gardenia jasminoides plants. Alex. Sci. Exch. J. 2019, 40, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Hatzilazarou, S.P.; Syros, T.D.; Yupsanis, T.A.; Bosabalidis, A.M.; Economou, A.S. Peroxidases, lignin and anatomy during in vitro and ex vitro rooting of gardenia (Gardenia jasminoides Ellis) microshoots. J. Plant Physiol. 2006, 163, 827–836. [Google Scholar] [CrossRef]
- Economou, A.S.; Spanoudaki, M.J. In vitro propagation of gardenia. HortScience 1985, 20, 213. [Google Scholar] [CrossRef]
- George, P.S.; Ravishankar, G.A.; Venkataraman, L.V. Clonal multiplication of Gardenia jasminoides Ellis through axillary bud culture. Plant Cell Rep. 1993, 13, 59–62. [Google Scholar] [CrossRef]
- El-Sayeda, I.M.; El-Ziatb, R.A.; Muradc, S.A.; Tahaa, L.S.; Mahgouba, M.H. Optimization of micropropagation protocol and secondary metabolites of Gardenia jasminoides plant. Plant Arch. 2020, 20, 9183–9189. [Google Scholar]
- Serret, M.D.; Trillas, M.I.; Araus, J.L. The effect of in vitro culture conditions on the pattern of photoinhibition during acclimation of gardenia plantlets to ex vitro conditions. Photosynthetica 2001, 39, 67–73. [Google Scholar] [CrossRef]
- Serret, M.D.; Trillas, M.I. Effects of light and sucrose levels on the anatomy, ultrastructure, and photosynthesis of Gardenia jasminoides Ellis leaflets cultured in vitro. Int. J. Plant Sci. 2000, 161, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Kozak, D. The influence of light quality and ba on in vitro growth and development of Gardenia jasminoides Ellis. Acta Sci. Pol.-Hortorum Cultus 2011, 10, 65–73. [Google Scholar]
- Chuenboonngarm, N.; Charoonsote, S.; Bhamarapravati, S. Effect of BA and 2iP on shoot proliferation and somaclonal variation of Gardenia jasminoides Ellis in vitro culture. Sci. Asia 2001, 27, 137–141. [Google Scholar] [CrossRef]
- Ebrahim, M.A.; Mohamed Esmaiel, N.; Ibrahim Abido, A.; Kadry Gaber, M. Regeneration, in vitro mutation and evaluation of genetic stability of gardenia somaclones via ssr markers. Alex. Sci. Exch. J. 2022, 43, 261–269. [Google Scholar] [CrossRef]
- Tawfik, E.; Fathy, M. Chemical mutagens affecting in vitro behavior of Gardenia jasminoides. Plant Tissue Cult. Biotechnol. 2020, 30, 209–218. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Kostas, S.; Nendou, T.; Economou, A. Conservation, Regeneration and Genetic Stability of Regenerants from Alginate-Encapsulated Shoot Explants of Gardenia jasminoides Ellis. Polymers 2021, 13, 1666. [Google Scholar] [CrossRef]
- Berkov, S.; Pavlov, A.; Georgiev, V.; Weber, J.; Bley, T.; Viladomat, F.; Bastida, J.; Codina, C. Changes in apolar metabolites during in vitro organogenesis of Pancratium maritimum. Plant Physiol. Biochem. 2010, 48, 827–835. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A brief review of phenolic compounds identified from plants: Their extraction, analysis, and biological activity. Nat. Prod. Commun. 2022, 17, 1934578X211069721. [Google Scholar] [CrossRef]
- Yoga, I.; Suprapta, D.; Jawi, I.; Permana, I. A study on the antioxidant and active compounds of Gardenia jasminoides Ellis (GJE) leaves extract. J. Agric. Sci. 2022, 17, 445–457. [Google Scholar] [CrossRef]
- Xu, Y.-Q.; Wei, G.-Y.; Zhou, Y.I.N.; Ge, F.E.I.; Luo, G.-M. Isolation and characterization of twenty-two polymorphic microsatellite markers from Gardenia jasminoides (Rubiaceae). J. Genet. 2015, 94, 22–24. [Google Scholar] [CrossRef]
- Deng, S.-Y.; Wang, X.-R.; Zhu, P.-L.; Wen, Q.; Yang, C.-X. Development of polymorphic microsatellite markers in the medicinal plant Gardenia jasminoides (Rubiaceae). Biochem. Syst. Ecol. 2015, 58, 149–155. [Google Scholar] [CrossRef]
- Steingroewer, J.; Bley, T.; Georgiev, V.; Ivanov, I.; Lenk, F.; Marchev, A.; Pavlov, A. Bioprocessing of differentiated plant in vitro systems. Eng. Life Sci. 2013, 13, 26–38. [Google Scholar] [CrossRef]
- Nikolova, M.; Aneva, I.; Zhelev, P.; Berkov, S. GC/MS based metabolite profiling and antioxidant activity of balkan and bulgarian endemic plants. Agric. Conspec. Sci. 2019, 84, 59–65. [Google Scholar]
- Berkov, S.; Georgieva, L.; Sidjimova, B.; Nikolova, M. Metabolite profiling of in vitro plant systems. In Bioprocessing of Plant In Vitro Systems; Pavlov, A., Bley, T., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 67–83. [Google Scholar]
- Mladenova, T.; Stoyanov, P.; Denev, P.; Dimitrova, S.; Katsarova, M.; Teneva, D.; Todorov, K.; Bivolarska, A. Phytochemical Composition, Antioxidant and Antimicrobial Activity of the Balkan Endemic Micromeria frivaldszkyana (Degen) Velen. (Lamiaceae). Plants 2021, 10, 710. [Google Scholar] [CrossRef]
- Georgiev, V.; Ananga, A.; Dincheva, I.; Badjakov, I.; Gochev, V.; Tsolova, V. Chemical Composition, In Vitro Antioxidant Potential, and Antimicrobial Activities of Essential Oils and Hydrosols from Native American Muscadine Grapes. Molecules 2019, 24, 3355. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Plant, µg/g DW | Shoots, µg/g DW | Callus, µg/g DW | Cell Suspension, µg/g DW |
|---|---|---|---|---|
| Protocatechuic acid | ND b,* | 8.65 ± 0.41 a | ND b | ND b |
| (+)-Catechin | ND c | ND c | 28.24 ± 0.99 b | 47.13 ± 3.40 a |
| Chlorogenic acid | 173.02 ± 9.67 b | 411.47 ± 11.67 a | 10.21 ± 0.77 c | 12.08 ± 0.27 c |
| Vanillic acid | ND c | 375.02 ± 19.34 a | 16.21 ± 1.03 b | 17.48 ± 1.38 b |
| Caffeic acid | 84.34 ± 9.16 a | 97.57 ± 7.12 a | 28.32 ± 0.95 b | ND c |
| Syringic acid | 84.68 ± 1.34 a | 26.85 ± 1.58 b | 16.98 ± 0.01 c | 14.01 ± 2.96 c |
| (−)-Epicatechin | 386.83 ± 10.90 a | 177.86 ± 4.53 b | 48.44 ± 1.84 c | 53.31 ± 2.86 c |
| p-Coumaric acid | ND b | 15.15 ± 2.52 a | ND b | ND b |
| Ferulic acid | 83.38 ± 9.93 a | 32.64 ± 3.28 b | 21.76 ± 2.40 bc | 13.74 ± 1.32 c |
| Salicylic acid | 1344.97 ± 100.11 a | 1262.82 ± 193.99 a | 10.67 ± 3.37 b | 9.78 ± 1.09 b |
| Rutin | 40.75 ± 1.50 a | 50.35 ± 9.59 a | 3.18 ± 0.40 b | 3.06 ± 0.07 b |
| Hesperidin | 309.32 ± 13.40 a | 233.42 ± 46.31 b | ND c | ND c |
| Rosmarinic acid | 1636.22 ± 135.45 a | 419.72 ± 101.73 b | 31.27 ± 6.50 c | 32.30 ± 1.98 c |
| Quercetin | 39.33 ± 2.45 b | 284.28 ± 22.85 a | 0.12 ± 0.02 c | ND c |
| Kaempferol | 4.15 ± 1.63 b | 286.05 ± 24.56 a | ND c | ND c |
| Assay | Plant | Shoots | Callus | Cell Suspension |
|---|---|---|---|---|
| Total Phenolics, mg GAE/gDW | 4.05 ± 0.36 a | 3.03 ± 0.20 b,* | 0.18 ± 0.02 c,* | 0.27 ± 0.01 c,* |
| DPPH, µM TE/g DW | 368.41 ± 41.77 b | 647.21 ± 33.29 a,* | 8.73 ± 1.50 c,* | 18.29 ± 2.69 c,* |
| TEAC, µM TE/g DW | 2162.79 ± 62.62 a | 1394.90 ± 15.00 b,* | 12.50 ± 1.72 c,* | 23.57 ± 2.09 c,* |
| FRAP, µM TE/g DW | 251.68 ± 16.45 a | 234.56 ± 15.59 a,* | 5.47 ± 0.65 b,* | 10.95 ± 0.50 b,* |
| CUPRAC, µM TE/g DW | 349.74 ± 39.39 a | 281.28 ± 17.34 b,* | 49.56 ± 8.50 c,* | 53.45 ± 8.41 c,* |
| SSR Locus | Na | Ne | Ho | He | PIC |
|---|---|---|---|---|---|
| GJ02 | 3 | 2 | 1 | 0.833 | 0.555 |
| GJ03 | 5 | 3 | 1 | 0.933 | 0.744 |
| GJ04 | 4 | 2 | 1 | 1 | 0.703 |
| GJ08 | 2 | 1 | 1 | 1 | 0.375 |
| GJ09 | 8 | 4 | 1 | 1 | 0.861 |
| GJ10 | 8 | 4 | 1 | 1 | 0.861 |
| GJ16 | 7 | 4 | 1 | 0.964 | 0.825 |
| GJ17 | 4 | 2 | 1 | 1 | 0.703 |
| eGJ010 | 7 | 4 | 1 | 0.964 | 0.825 |
| eGJ118 | 8 | 4 | 1 | 1 | 0.861 |
| eGJ144 | 6 | 3 | 1 | 1 | 0.810 |
| mean ± SD | 5.636 ± 2.157 | 3.0 ± 1.095 | 1.0 ± 0.0 | 0.972 ± 0.051 | 0.738 ± 0.153 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krasteva, G.; Berkov, S.; Pavlov, A.; Georgiev, V. Metabolite Profiling of Gardenia jasminoides Ellis In Vitro Cultures with Different Levels of Differentiation. Molecules 2022, 27, 8906. https://doi.org/10.3390/molecules27248906
Krasteva G, Berkov S, Pavlov A, Georgiev V. Metabolite Profiling of Gardenia jasminoides Ellis In Vitro Cultures with Different Levels of Differentiation. Molecules. 2022; 27(24):8906. https://doi.org/10.3390/molecules27248906
Chicago/Turabian StyleKrasteva, Gergana, Strahil Berkov, Atanas Pavlov, and Vasil Georgiev. 2022. "Metabolite Profiling of Gardenia jasminoides Ellis In Vitro Cultures with Different Levels of Differentiation" Molecules 27, no. 24: 8906. https://doi.org/10.3390/molecules27248906
APA StyleKrasteva, G., Berkov, S., Pavlov, A., & Georgiev, V. (2022). Metabolite Profiling of Gardenia jasminoides Ellis In Vitro Cultures with Different Levels of Differentiation. Molecules, 27(24), 8906. https://doi.org/10.3390/molecules27248906