

The Potential Role of Fisetin, a Flavonoid in Cancer Prevention and Treatment
 ,
,  , and
, and
Abstract
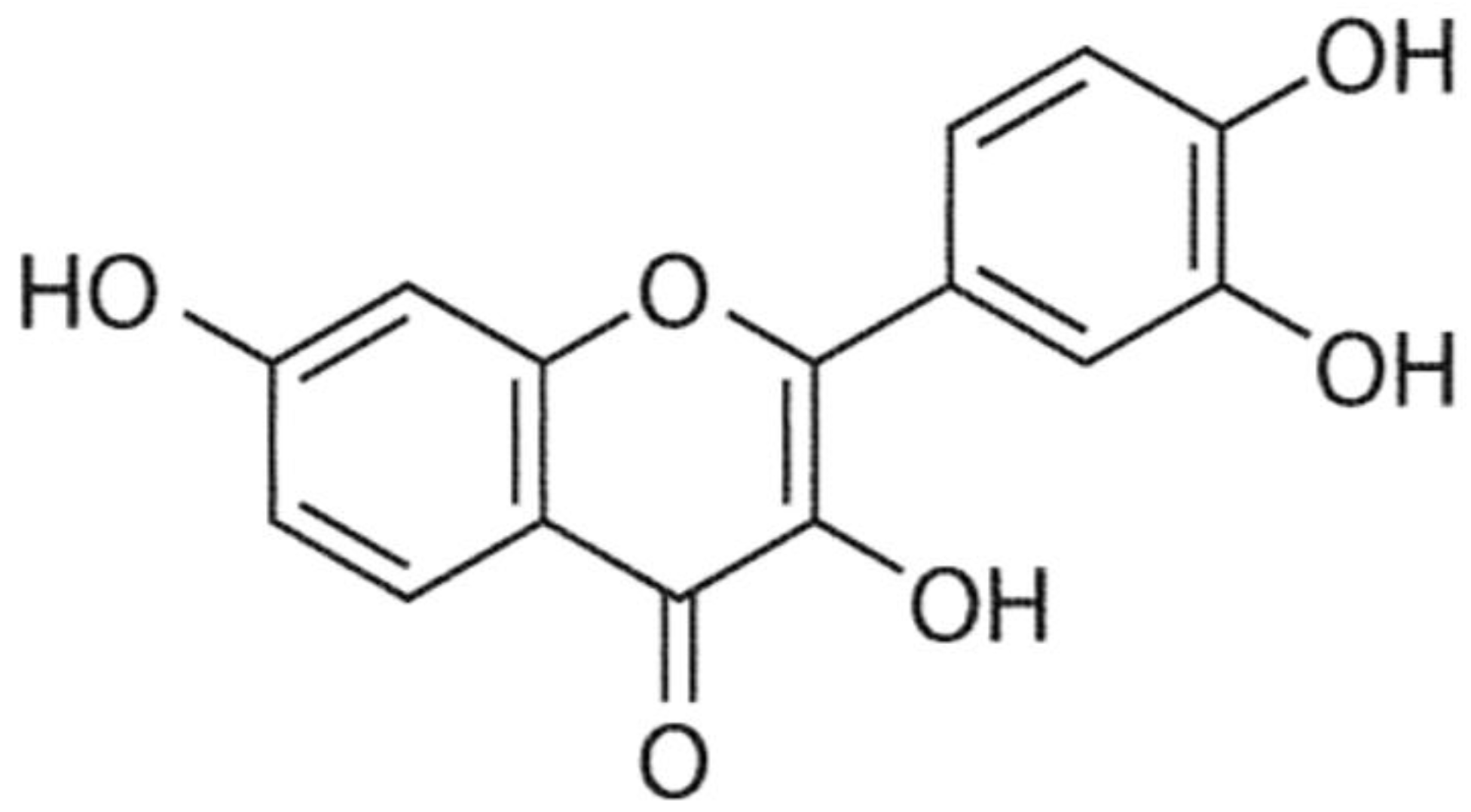
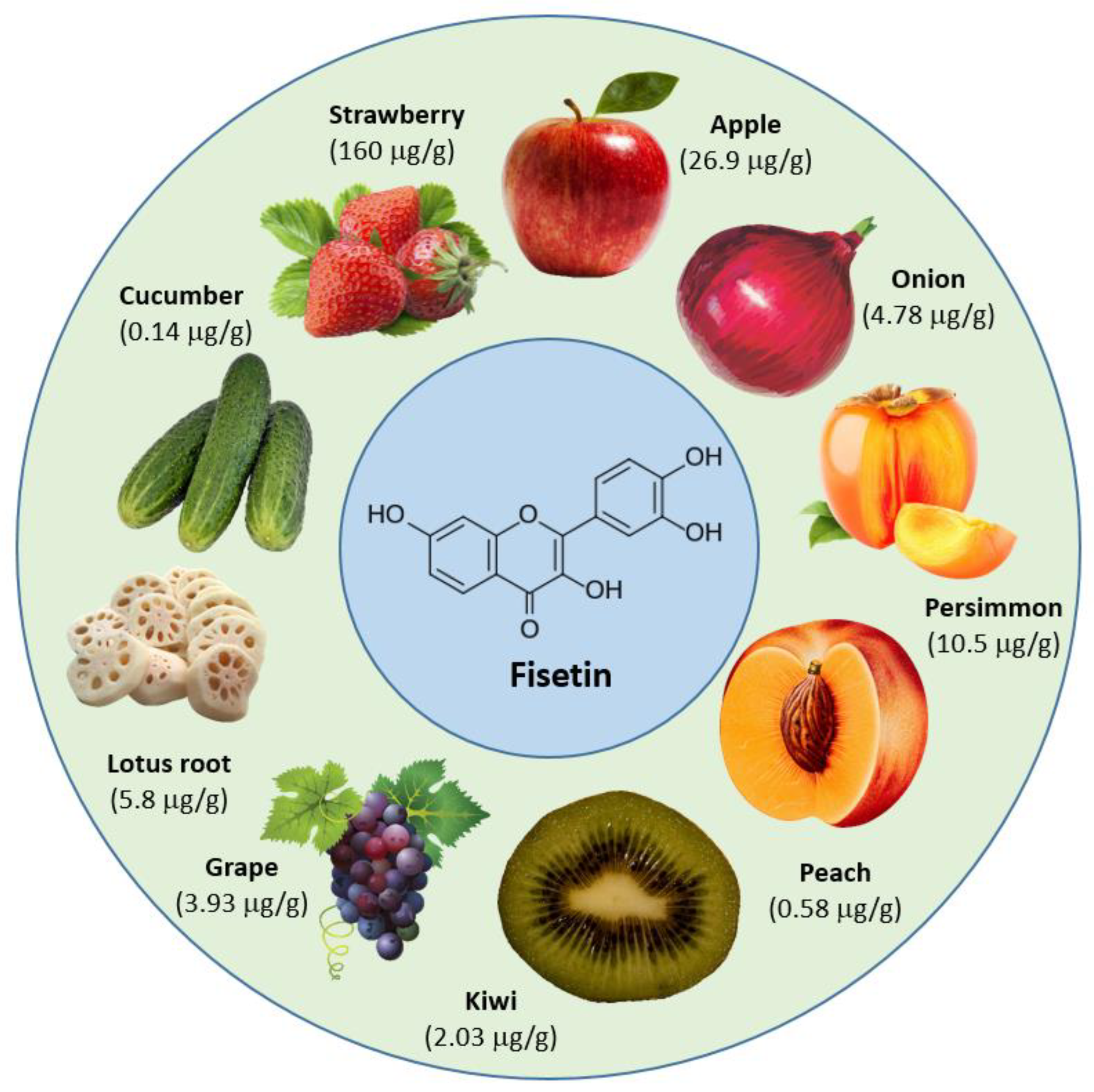
:1. Introduction
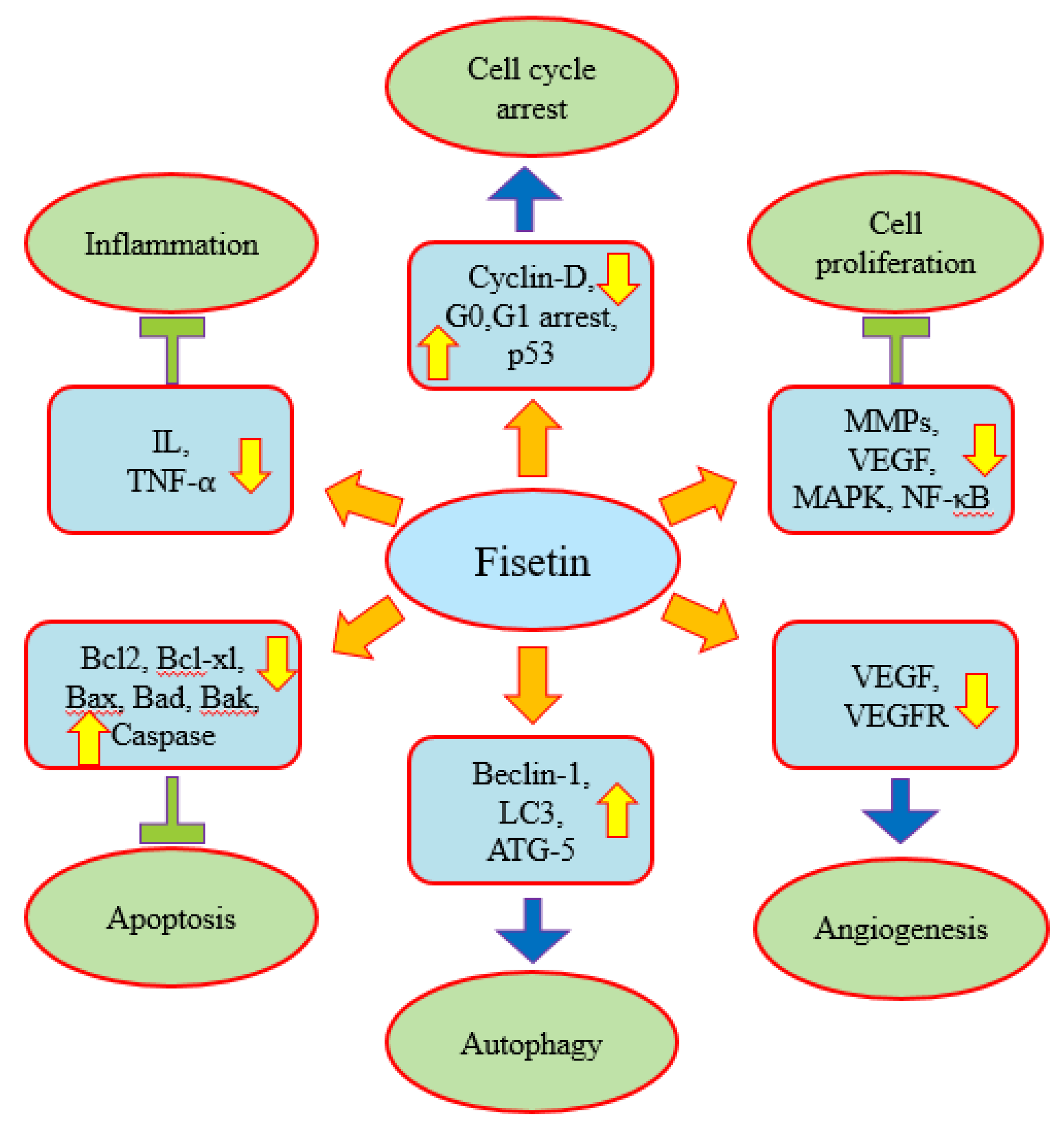
2. Anticancer Molecular Target and Mechanism-of-Action of Fisetin
2.1. Inflammation
2.2. Apoptosis
2.3. Autophagy
2.4. Angiogenesis
2.5. Cell Cycle Arrest
2.6. PI3K/AKT/mTOR Pathway
2.7. Signal Transducer and Activator of Transcription 3 (STAT3)
2.8. Wnt/β-Catenin Signaling Pathway
2.9. NF-κB Pathway
2.10. Nrf2 Pathway
2.11. AP-1 pathway
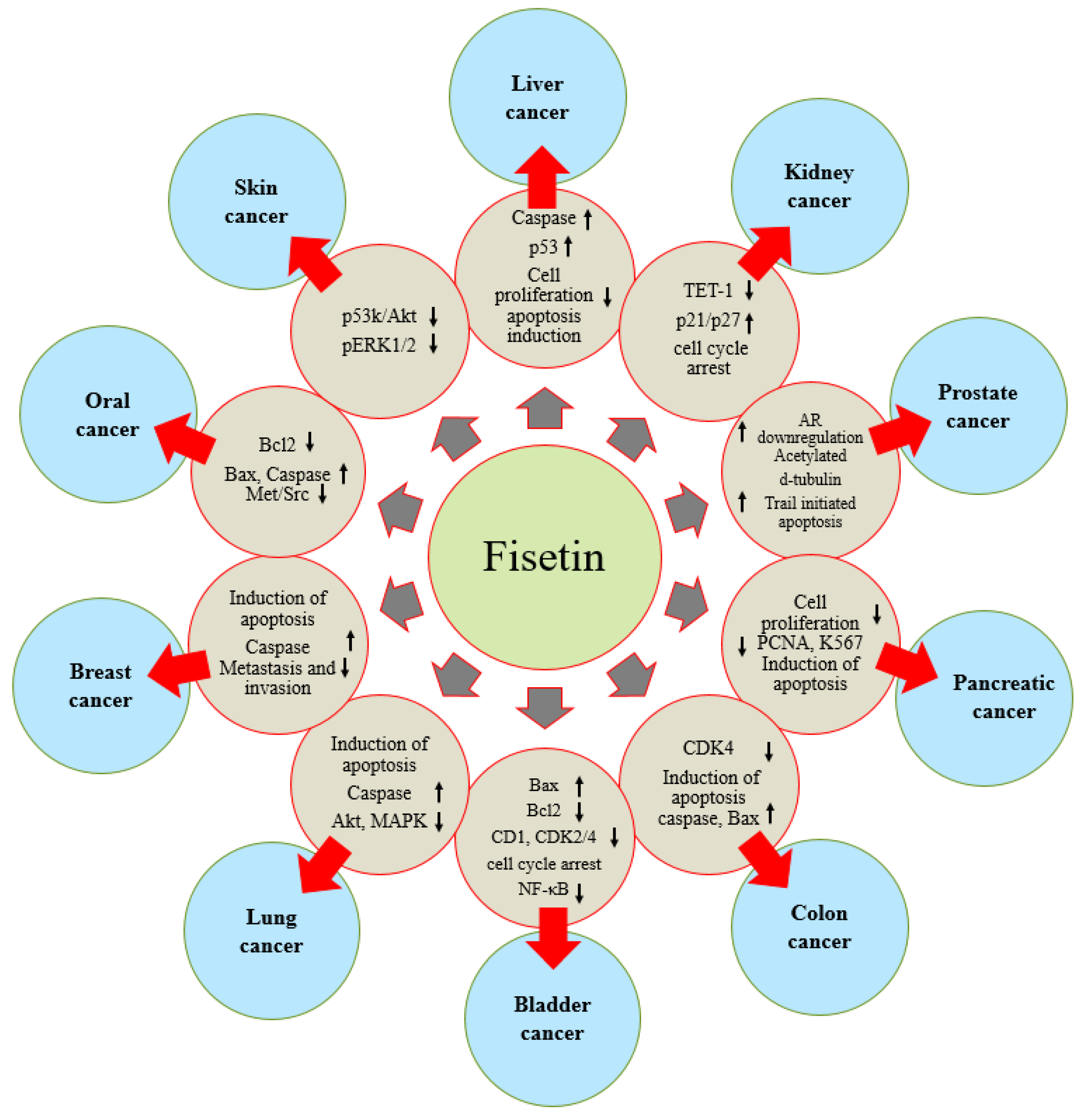
3. Fisetin: Role in Various Types of Cancer
3.1. Prostate Cancer
3.2. Kidney Cancer
3.3. Liver Cancer
3.4. Colon Cancer
3.5. Gastric Cancer
3.6. Pancreatic Cancer
3.7. Bile Duct Cancer
3.8. Bladder Cancer
3.9. Lung Cancer
3.10. Skin Cancer
3.11. Oral Cancer
3.12. Leukemia
3.13. Breast Cancer
3.14. Ovarian Cancer
3.15. Cervix Cancer
3.16. Endometrial Cancer
3.17. Osteosarcoma
3.18. Brain Cancer
3.19. Retinoblastoma
3.20. Lymphoma
3.21. Thyroid Cancer
4. Synergistic Effect of Fisetin with Anticancer Drugs
5. Pharmacokinetics of Fisetin and Strategies to Improve its Bioavailability
6. Clinical Trials and Patents on Fisetin
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devi, K.P.; Rajavel, T.; Nabavi, S.F.; Setzer, W.N.; Ahmadi, A.; Mansouri, K. Hesperidin: A promising anticancer agent from nature. Ind. Crop. Prod. 2015, 76, 582–589. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Tompa, D.R.; Immanuel, A.; Srikanth, S.; Kadhirvel, S. Trends and strategies to combat viral infections: A review on FDA approved antiviral drugs. Int. J. Biol. Macromol. 2021, 172, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic interactions of phytochemicals with antimicrobial agents: Potential strategy to counteract drug resistance. Chem. Interact. 2019, 308, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Ayaz, A.; Saqib, S.; Zaman, W.; Butt, M.A.; Ullah, A. Silene conoidea L.: A Review on its Systematic, Ethnobotany and Phytochemical profile. Plant Sci. Today 2019, 6, 373–382. [Google Scholar] [CrossRef]
- Liskova, A.; Koklesova, L.; Samec, M.; Smejkal, K.; Samuel, S.M.; Varghese, E.; Abotaleb, M.; Biringer, K.; Kudela, E.; Danko, J.; et al. Flavonoids in Cancer Metastasis. Cancers 2020, 12, 1498. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Almatroudi, A.; Alsahli, M.A.; Khan, A.A.; Rahmani, A.H. Thymoquinone, an Active Compound of Nigella sativa: Role in Prevention and Treatment of Cancer. Curr. Pharm. Biotechnol. 2020, 21, 1028–1041. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydhi, F.A.; Alsahli, M.A.; Rahmani, A.H. Potential Therapeutic Targets of Epigallocatechin Gallate (EGCG), the Most Abundant Catechin in Green Tea, and its Role in the Therapy of Various Types of Cancer. Molecules 2020, 25, 3146. [Google Scholar] [CrossRef]
- Syed, M.A.; Rahmani, A.H. Potential Therapeutic Targets of Curcumin, Most Abundant Active Compound of Turmeric Spice: Role in the Management of Various Types of Cancer. Recent Patents Anti-Cancer Drug Discov. 2021, 16, 3–29. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Almatroudi, A.; Khan, A.A.; Babiker, A.Y.; Alanezi, M.; Allemailem, K.S. The Multifaceted Role of Baicalein in Cancer Management through Modulation of Cell Signalling Pathways. Molecules 2022, 27, 8023. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Delarami Far, A.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G.; et al. Anticancer and apoptosis-inducing effects of quercetin in vitro and in vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almatroodi, S.A.; Alsahli, M.A.; Rahmani, A.H. Berberine: An Important Emphasis on Its Anticancer Effects through Modulation of Various Cell Signaling Pathways. Molecules 2022, 27, 5889. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Alsahli, M.A.; Almatroudi, A.; Almogbel, M.A.; Khan, A.A.; Anwar, S.; Almatroodi, S.A. The Potential Role of Apigenin in Cancer Prevention and Treatment. Molecules 2022, 27, 6051. [Google Scholar] [CrossRef] [PubMed]
- Almatroudi, A.; Alsahli, M.A.; Alrumaihi, F.; Allemailem, K.S.; Rahmani, A.H. Ginger: A Novel Strategy to Battle Cancer through Modulating Cell Signalling Pathways: A Review. Curr. Pharm. Biotechnol. 2019, 20, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Kubina, R.; Iriti, M.; Kabała-Dzik, A. Anticancer Potential of Selected Flavonols: Fisetin, Kaempferol, and Quercetin on Head and Neck Cancers. Nutrients 2021, 13, 845. [Google Scholar] [CrossRef] [PubMed]
- Kimira, M.; Arai, Y.; Shimoi, K.; Watanabe, S. Japanese intake of flavonoids and isoflavonoids from foods. J. Epidemiol. 1998, 8, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, A.K.; Garg, G.; Rizvi, S.I. Fisetin as a caloric restriction mimetic protects rat brain against aging induced oxidative stress, apoptosis and neurodegeneration. Life Sci. 2018, 193, 171–179. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Zhou, Y.; Zhu, Y.; Fei, M. Fisetin alleviates oxidative stress after traumatic brain injury via the Nrf2-ARE pathway. Neurochem. Int. 2018, 118, 304–313. [Google Scholar] [CrossRef]
- Wu, P.-Y.; Lyu, J.-L.; Liu, Y.-J.; Chien, T.-Y.; Hsu, H.-C.; Wen, K.-C.; Chiang, H.-M. Fisetin Regulates Nrf2 Expression and the Inflammation-Related Signaling Pathway to Prevent UVB-Induced Skin Damage in Hairless Mice. Int. J. Mol. Sci. 2017, 18, 2118. [Google Scholar] [CrossRef]
- Althunibat, O.Y.; Al Hroob, A.M.; Abukhalil, M.H.; Germoush, M.O.; Bin-Jumah, M.; Mahmoud, A.M. Fisetin ameliorates oxidative stress, inflammation and apoptosis in diabetic cardiomyopathy. Life Sci. 2019, 221, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A Dietary Antioxidant for Health Promotion. Antioxidants Redox Signal. 2013, 19, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Saeed, F.; Gilani, S.A.; Shariati, M.A.; Imran, A.; Afzaal, M.; Atif, M.; Tufail, T.; Anjum, F.M. Fisetin: An anticancer perspective. Food Sci. Nutr. 2021, 9, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, A.; O’Hagan, H. Interplay Between Inflammation and Epigenetic Changes in Cancer. Prog. Mol. Biol. Transl. Sci. 2016, 144, 69–117. [Google Scholar] [CrossRef] [PubMed]
- Germolec, D.R.; Shipkowski, K.A.; Frawley, R.P.; Evans, E. Markers of Inflammation. Methods Mol. Biol. 2018, 1803, 57–79. [Google Scholar]
- Kay, J.; Thadhani, E.; Samson, L.; Engelward, B. Inflammation-induced DNA damage, mutations and cancer. DNA Repair 2019, 83, 102673. [Google Scholar] [CrossRef]
- Korniluk, A.; Koper, O.; Kemona, H.; Dymicka-Piekarska, V. From inflammation to cancer. Ir. J. Med. Sci. 2017, 186, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Karin, M.; Sun, B. Targeting cancer-promoting inflammation-have anti-inflammatory therapies come of age? Nat. Rev. Clin. Oncol. 2021, 18, 261–279. [Google Scholar] [CrossRef]
- Alsahli, M.A.; Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Anwar, S.; Almutary, A.G.; Alrumaihi, F.; Rahmani, A.H. 6-Gingerol, a major ingredient of ginger attenuates Diethylnitrosamine-induced liver injury in rats through the modulation of oxidative stress and anti-inflammatory activity. Mediat. Inflamm. 2021, 2021, 6661937. [Google Scholar] [CrossRef]
- Alzohairy, M.A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. Protective Effects of Thymoquinone, Active Compound of Nigella sativa, on Rats with Benzo(a)pyrene-Induced Lung Injury through Regulation of Oxidative Stress and Inflammation. Molecules 2021, 26, 3218. [Google Scholar] [CrossRef]
- Sun, Y.; Qin, H.; Zhang, H.; Feng, X.; Yang, L.; Hou, D.-X.; Chen, J. Fisetin inhibits inflammation and induces autophagy by mediating PI3K/AKT/mTOR signaling in LPS-induced RAW264.7 cells. Food Nutr. Res. 2021, 65, 6355. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.; Alnuqaydan, A.; Alsahli, M.; Khan, A.; Rahmani, A. Thymoquinone, the Most Prominent Constituent of Nigella Sativa, Attenuates Liver Damage in Streptozotocin-Induced Diabetic Rats via Regulation of Oxidative Stress, Inflammation and Cyclooxygenase-2 Protein Expression. Appl. Sci. 2021, 11, 3223. [Google Scholar] [CrossRef]
- Jiang, K.; Yang, J.; Xue, G.; Dai, A.; Wu, H. Fisetin Ameliorates the Inflammation and Oxidative Stress in Lipopolysaccharide-Induced Endometritis. J. Inflamm. Res. 2021, 14, 2963–2978. [Google Scholar] [CrossRef] [PubMed]
- Farsad-Naeimi, A.; Alizadeh, M.; Esfahani, A.; Darvish Aminabad, E. Effect of fisetin supplementation on inflammatory factors and matrix metalloproteinase enzymes in colorectal cancer patients. Food Funct. 2018, 9, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Arbiser, J.L.; Bonner, M.Y.; Gilbert, L.C. Targeting the duality of cancer. NPJ Precis. Oncol. 2017, 1, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Jing, L.; Wang, Q.; Lin, C.-C.; Chen, X.; Diao, J.; Liu, Y.; Sun, X. Bas-PGAM5L-Drp1 complex is required for intrinsic apoptosis execution. Oncotarget 2015, 6, 30017–30034. [Google Scholar] [CrossRef] [Green Version]
- Won, D.H.; Chung, S.H.; Shin, J.A.; Hong, K.O.; Yang, I.H.; Yun, J.W.; Cho, S.D. Induction of sestrin 2 is associated with fisetin-mediated apoptosis in human head and neck cancer cell lines. J. Clin. Biochem. Nutr. 2019, 64, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Min, K.-J.; Nam, J.-O.; Kwon, T.K. Fisetin Induces Apoptosis Through p53-Mediated Up-Regulation of DR5 Expression in Human Renal Carcinoma Caki Cells. Molecules 2017, 22, 1285. [Google Scholar] [CrossRef] [Green Version]
- Park, B.-S.; Choi, N.-E.; Lee, J.H.; Kang, H.-M.; Yu, S.-B.; Kim, H.-J.; Kang, H.-K.; Kim, I.-R. Crosstalk between Fisetin-induced Apoptosis and Autophagy in Human Oral Squamous Cell Carcinoma. J. Cancer 2019, 10, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Afroze, N.; Pramodh, S.; Shafarin, J.; Bajbouj, K.; Hamad, M.; Sundaram, M.K.; Haque, S.; Hussain, A. Fisetin Deters Cell Proliferation, Induces Apoptosis, Alleviates Oxidative Stress and Inflammation in Human Cancer Cells, HeLa. Int. J. Mol. Sci. 2022, 23, 1707. [Google Scholar] [CrossRef]
- Wang, K.; Hu, D.-N.; Lin, H.-W.; Yang, W.-E.; Hsieh, Y.-H.; Chien, H.-W.; Yang, S.-F. Fisetin induces apoptosis through mitochondrial apoptosis pathway in human uveal melanoma cells. Environ. Toxicol. 2018, 33, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhao, L.; Kuang, M.; Zhang, B.; Liang, Z.; Yi, T.; Wei, Y.; Zhao, X. Autophagy in tumorigenesis and cancer therapy: Dr. Jekyll or Mr. Hyde? Cancer Lett. 2012, 323, 115–127. [Google Scholar] [CrossRef]
- White, E. Deconvoluting the context-dependent role for autophagy in cancer. Nat. Rev. Cancer 2012, 12, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, E.; DiPaola, R.S. The Double-Edged Sword of Autophagy Modulation in Cancer. Clin. Cancer Res. 2009, 15, 5308–5316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, S.; Xu, X.; Zhou, S.; Chen, Y.; Ding, G.; Cao, L. Fisetin induces autophagy in pancreatic cancer cells via endoplasmic reticulum stress- and mitochondrial stress-dependent pathways. Cell Death Dis. 2019, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and Therapeutic Aspects of Angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, N.; Cheng, H. Fisetin inhibits vascular endothelial growth factor-induced angiogenesis in retinoblastoma cells. Oncol. Lett. 2020, 20, 1239–1244. [Google Scholar] [CrossRef]
- Bhat, T.A.; Nambiar, D.; Pal, A.; Agarwal, R.; Singh, R.P. Fisetin inhibits various attributes of angiogenesis in vitro and in vivo--implications for angioprevention. Carcinogenesis 2012, 33, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Touil, Y.S.; Seguin, J.; Scherman, D.; Chabot, G.G. Improved antiangiogenic and antitumour activity of the combination of the natural flavonoid fisetin and cyclophosphamide in Lewis lung carcinoma-bearing mice. Cancer Chemother. Pharmacol. 2011, 68, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Jang, Y.-J.; Choi, Y.J.; Jang, J.W.; Kim, J.-H.; Rho, Y.-K.; Kim, I.J.; Kim, H.-J.; Leem, M.J.; Lee, S.-T. Fisetin Inhibits Matrix Metalloproteinases and Reduces Tumor Cell Invasiveness and Endothelial Cell Tube Formation. Nutr. Cancer 2013, 65, 1192–1199. [Google Scholar] [CrossRef]
- Li, J.; Cheng, Y.; Qu, W.; Sun, Y.; Wang, Z.; Wang, H.; Tian, B. Fisetin, a dietary flavonoid, induces cell cycle arrest and apoptosis through activation of p53 and inhibition of NF-kappa B pathways in bladder cancer cells. Basic Clin. Pharmacol. Toxicol. 2011, 108, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.H.; Lin, J.J.; Ma, Y.S.; Peng, S.F.; Huang, A.C.; Huang, Y.P.; Fan, M.J.; Lien, J.C.; Chung, J.G. Fisetin inhibits cell proliferation through the induction of G0/G1 phase arrest and caspase-3-mediated apoptosis in mouse leukemia cells. Am. J. Chin. Med. 2019, 47, 841–863. [Google Scholar] [CrossRef] [PubMed]
- Adan, A.; Baran, Y. The pleiotropic effects of fisetin and hesperetin on human acute promyelocytic leukemia cells are mediated through apoptosis, cell cycle arrest, and alterations in signaling networks. Tumor Biol. 2015, 36, 8973–8984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.J.; Jia, S.S. Fisetin inhibits laryngeal carcinoma through regulation of AKT/NF-κB/mTOR and ERK1/2 signaling pathways. Biomed Pharm. 2016, 83, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, Y.; Gao, Z.; Li, X.; Weng, M.; Shi, C.; Wang, C.; Sun, L. Fisetin inhibits the proliferation, migration and invasion of pancreatic cancer by targeting PI3K/AKT/mTOR signaling. Aging 2021, 13, 24753–24767. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Khusro, F.H.; Mustafa Adhami, V.; Suh, Y.; Mukhtar, H. Dual inhibition of phosphatidylinositol 3-kinase/Akt and mammalian target of rapamycin signaling in human nonsmall cell lung cancer cells by a dietary flavonoid fisetin. Int. J. Cancer 2012, 130, 1695–1705. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Ma, X.; Li, Q.; Yang, Y.; Xu, X.; Sun, J.; Yu, M.; Cao, K.; Yang, L.; Yang, G.; et al. Anti-cancer effects of fisetin on mammary carcinoma cells via regulation of the PI3K/Akt/mTOR pathway: In vitro and in vivo studies. Int. J. Mol. Med. 2018, 42, 811–820. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.J.; Leung, K.H.; Chan DS, H.; Wang, Y.T.; Ma, D.L.; Leung, C.H. Identification of a natural product-like STAT3 dimerization inhibitor by structure-based virtual screening. Cell Death Dis. 2014, 5, e1293. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Qu, Z.; Sun, F.; Han, L.; Li, L.; Yan, S.; Stabile, L.P.; Chen, L.F.; Siegfried, J.M.; Xiao, G. Myeloid STAT3 promotes lung tumorigenesis by transforming tumor immunosurveillance into tumor-promoting inflammation. Cancer Immunol. Res. 2017, 5, 257. [Google Scholar] [CrossRef] [Green Version]
- Song, T.L.; Nairismägi, M.-L.; Laurensia, Y.; Lim, J.-Q.; Tan, J.; Li, Z.-M.; Pang, W.-L.; Kizhakeyil, A.; Wijaya, G.-C.; Huang, D.; et al. Oncogenic activation of the STAT3 pathway drives PD-L1 expression in natural killer/T-cell lymphoma. Blood 2018, 132, 1146–1158. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 signalling in cancer: New and unexpected biological functions. Nat. Rev. Cancer 2014, 14, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Kong, D.; Zhang, Y.; Li, S.; Li, Y.; Ramamoorthy, A.; Ma, J. Fisetin Inhibits Cell Proliferation and Induces Apoptosis via JAK/STAT3 Signaling Pathways in Human Thyroid TPC 1 Cancer Cells. Biotechnol. Bioprocess Eng. 2020, 25, 197–205. [Google Scholar] [CrossRef]
- Pai, S.G.; Carneiro, B.A.; Mota, J.M.; Costa, R.; Leite, C.A.; Barroso-Sousa, R.; Kaplan, J.B.; Chae, Y.K.; Giles, F.J. Wnt/beta-catenin pathway: Modulating anticancer immune response. J. Hematol. Oncol. 2017, 10, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Zhou, X.; Zhang, W.; Qu, Y.; Ke, X. Is β-Catenin a Druggable Target for Cancer Therapy? Trends Biochem. Sci. 2018, 43, 623–634. [Google Scholar] [CrossRef]
- Qin, J.-J.; Wang, W.; Li, X.; Deokar, H.; Buolamwini, J.K.; Zhang, R. Inhibiting β-Catenin by β-Carboline-Type MDM2 Inhibitor for Pancreatic Cancer Therapy. Front. Pharmacol. 2018, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Xu, X.; Chen, D.; Zhao, F.; Wang, W. Therapeutic potential of targeting the Wnt/β-catenin signaling pathway in colorectal cancer. Biomed. Pharmacother. 2018, 110, 473–481. [Google Scholar] [CrossRef]
- Suh, Y.; Afaq, F.; Johnson, J.J.; Mukhtar, H. A plant flavonoid fisetin induces apoptosis in colon cancer cells by inhibition of COX2 and Wnt/EGFR/NF-kappaB-signaling pathways. Carcinogenesis 2009, 30, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Syed, D.N.; Afaq, F.; Maddodi, N.; Johnson, J.J.; Sarfaraz, S.; Ahmad, A.; Setaluri, V.; Mukhtar, H. Inhibition of Human Melanoma Cell Growth by the Dietary Flavonoid Fisetin Is Associated with Disruption of Wnt/β-Catenin Signaling and Decreased Mitf Levels. J. Investig. Dermatol. 2011, 131, 1291–1299. [Google Scholar] [CrossRef] [Green Version]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A. The Keap1-Nrf2pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Pal, H.C.; Sharma, S.; Strickland, L.R.; Katiyar, S.K.; Ballestas, M.E.; Athar, M.; Elmets, C.A.; Afaq, F. Fisetin Inhibits Human Melanoma Cell Invasion through Promotion of Mesenchymal to Epithelial Transition and by Targeting MAPK and NFκB Signaling Pathways. PLoS ONE 2014, 9, e86338. [Google Scholar] [CrossRef] [PubMed]
- Menegon, S.; Columbano, A.; Giordano, S. The Dual Roles of NRF2 in Cancer. Trends Mol. Med. 2016, 22, 578–593. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Namani, A.; Wu, S.; Wang, X. Nrf2: Bane or blessing in cancer? J. Cancer Res. Clin. Oncol. 2014, 140, 1251–1259. [Google Scholar] [CrossRef]
- Bai, X.; Chen, Y.; Hou, X.; Huang, M.; Jin, J. Emerging role of NRF2 in chemoresistance by regulating drug-metabolizing enzymes and efflux transporters. Drug Metab. Rev. 2016, 48, 541–567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, W.; Feng, X.; Yang, F.; Qin, H.; Wu, S.; Hou, D.-X.; Chen, J. Nrf2–ARE Signaling Acts as Master Pathway for the Cellular Antioxidant Activity of Fisetin. Molecules 2019, 24, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-F.; Chen, J.-H.; Chang, C.-N.; Lu, D.-Y.; Chang, P.-C.; Wang, S.-L.; Yeh, W.-L. Fisetin inhibits cell migration via inducing HO-1 and reducing MMPs expression in breast cancer cell lines. Food Chem. Toxicol. 2018, 120, 528–535. [Google Scholar] [CrossRef]
- Trop-Steinberg, S.; Azar, Y. AP-1 Expression and its Clinical Relevance in Immune Disorders and Cancer. Am. J. Med. Sci. 2017, 353, 474–483. [Google Scholar] [CrossRef]
- Kappelmann, M.; Bosserhoff, A.; Kuphal, S. AP-1/c-Jun transcription factors: Regulation and function in malignant melanoma. Eur. J. Cell Biol. 2014, 93, 76–81. [Google Scholar] [CrossRef]
- Fu, C.; Chen, M.; Tseng, Y.; Chen, M.; Zhou, Z.; Yang, J.; Lin, Y.; Viswanadha, V.P.; Wang, G.; Huang, C. Fisetin activates Hippo pathway and JNK/ERK/AP-1 signaling to inhibit proliferation and induce apoptosis of human osteosarcoma cells via ZAK overexpression. Environ. Toxicol. 2019, 34, 902–911. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Dai, C.; Heemers, H.; Sharifi, N. Androgen signaling in prostate cancer. Cold Spring Harb. Perspect. Med. 2017, 7, a030452. [Google Scholar] [CrossRef] [PubMed]
- Lall, R.K.; Syed, D.N.; Khan, M.I.; Adhami, V.M.; Gong, Y.; Lucey, J.A.; Mukhtar, H. Dietary flavonoid fisetin increases abundance of high-molecular-mass hyaluronan conferring resistance to prostate oncogenesis. Carcinogenesis 2016, 37, 918–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhtar, E.; Adhami, V.M.; Sechi, M.; Mukhtar, H. Dietary flavonoid fisetin binds to β-tubulin and disrupts microtubule dynamics in prostate cancer cells. Cancer Lett. 2015, 367, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szliszka, E.; Helewski, K.J.; Mizgala, E.; Krol, W. The dietary flavonol fisetin enhances the apoptosis-inducing potential of TRAIL in prostate cancer cells. Int. J. Oncol. 2011, 39, 771–779. [Google Scholar] [CrossRef]
- Suh, Y.; Afaq, F.; Khan, N.; Johnson, J.J.; Khusro, F.H.; Mukhtar, H. Fisetin induces autophagic cell death through suppression of mTOR signaling pathway in prostate cancer cells. Carcinog. 2010, 31, 1424–1433. [Google Scholar] [CrossRef] [Green Version]
- Escudier, B.; Porta, C.; Schmidinger, M.; Rioux-Leclercq, N.; Bex, A.; Khoo, V.; Grünwald, V.; Gillessen, S.; Horwich, A.; ESMO Guidelines Committee. Renal cell carcinoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2019, 30, 706–720. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, M.-H.; Tsai, J.-P.; Yang, S.-F.; Chiou, H.-L.; Lin, C.-L.; Chang, H.-R. Fisetin Suppresses the Proliferation and Metastasis of Renal Cell Carcinoma through Upregulation of MEK/ERK-Targeting CTSS and ADAM9. Cells 2019, 8, 948. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.; Liu, J.; Shen, H.; Zhang, C.; Wu, Y.; Huang, Y.; Gong, Z.; Xue, J.; Liu, T. Fisetin decreases TET1 activity and CCNY/CDK16 promoter 5hmC levels to inhibit the proliferation and invasion of renal cancer stem cell. J. Cell. Mol. Med. 2019, 23, 1095–1105. [Google Scholar] [CrossRef]
- Sundarraj, K.; Raghunath, A.; Panneerselvam, L.; Perumal, E. Fisetin Inhibits Autophagy in HepG2 Cells via PI3K/Akt/mTOR and AMPK Pathway. Nutr. Cancer 2021, 73, 2502–2514. [Google Scholar] [CrossRef]
- Liu, X.-F.; Long, H.-J.; Miao, X.-Y.; Liu, G.-L.; Yao, H.-L. Fisetin inhibits liver cancer growth in a mouse model: Relation to dopamine receptor. Oncol. Rep. 2017, 38, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Jung, J.I.; Cho, H.J.; Lim, D.Y.; Lee, H.S.; Chun, H.S.; Kwon, D.Y.; Park, J.H.Y. Fisetin Inhibits the Activities of Cyclin-Dependent Kinases Leading to Cell Cycle Arrest in HT-29 Human Colon Cancer Cells. J. Nutr. 2005, 135, 2884–2890. [Google Scholar] [CrossRef] [PubMed]
- Jeng, L.; Velmurugan, B.K.; Chen, M.; Hsu, H.; Ho, T.; Day, C.; Lin, Y.; Padma, V.V.; Tu, C.; Huang, C. Fisetin mediated apoptotic cell death in parental and Oxaliplatin/irinotecan resistant colorectal cancer cells in vitro and in vivo. J. Cell. Physiol. 2018, 233, 7134–7142. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.Y.; Park, J.H.Y. Induction of p53 contributes to apoptosis of HCT-116 human colon cancer cells induced by the dietary compound fisetin. Am. J. Physiol. Liver Physiol. 2009, 296, G1060–G1068. [Google Scholar] [CrossRef]
- Yu, S.-H.; Yang, P.-M.; Peng, C.-W.; Yu, Y.-C.; Chiu, S.-J. Securin depletion sensitizes human colon cancer cells to fisetin-induced apoptosis. Cancer Lett. 2011, 300, 96–104. [Google Scholar] [CrossRef]
- Sabarwal, A.; Agarwal, R.; Singh, R.P. Fisetin inhibits cellular proliferation and induces mitochondria-dependent apoptosis in human gastric cancer cells. Mol. Carcinog. 2017, 56, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Chen, S.; Zhao, Y.; Ye, X. Fisetin inhibits the proliferation of gastric cancer cells and induces apoptosis through suppression of ERK 1/2 activation. Oncol. Lett. 2018, 15, 8442–8446. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kang, M.-J.; Lee, S.H.; Son, J.H.; Lee, J.E.; Paik, W.H.; Ryu, J.K.; Kim, Y.-T. Fisetin Enhances the Cytotoxicity of Gemcitabine by Down-regulating ERK-MYC in MiaPaca-2 Human Pancreatic Cancer Cells. Anticancer. Res. 2018, 38, 3527–3533. [Google Scholar] [CrossRef]
- Huang, C.; Zhou, S.; Zhang, C.; Jin, Y.; Xu, G.; Zhou, L.; Ding, G.; Pang, T.; Jia, S.; Cao, L. ZC3H13-mediated N6-methyladenosine modification of PHF10 is impaired by fisetin which inhibits the DNA damage response in pancreatic cancer. Cancer Lett. 2022, 530, 16–28. [Google Scholar] [CrossRef]
- Murtaza, I.; Adhami, V.M.; Hafeez, B.B.; Saleem, M.; Mukhtar, H. Fisetin, a natural flavonoid, targets chemoresistant human pancreatic cancer AsPC-1 cells through DR3-mediated inhibition of NF-kappaB. Int. J. Cancer. 2009, 125, 2465–2473. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Lee, S.H.; Son, J.H.; Lee, J.M.; Kang, M.J.; Kim, B.H.; Lee, J.S.; Ryu, J.K.; Kim, Y.T. Fisetin Reduces Cell Viability Through Up-Regulation of Phosphorylation of ERK1/2 in Cholangiocarcinoma Cells. Anticancer. Res. 2016, 36, 6109–6116. [Google Scholar] [CrossRef]
- Li, J.; Qu, W.; Cheng, Y.; Sun, Y.; Jiang, Y.; Zou, T.; Wang, Z.; Xu, Y.; Zhao, H. The Inhibitory Effect of Intravesical Fisetin against Bladder Cancer by Induction of p53 and Down-Regulation of NF-kappa B Pathways in a Rat Bladder Carcinogenesis Model. Basic Clin. Pharmacol. Toxicol. 2014, 115, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T.; Thun, M.J. Cancer statistics, 2008. CA A Cancer J. Clin. 2008, 58, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, S. Fisetin inhibits the growth and migration in the A549 human lung cancer cell line via the ERK1/2 pathway. Exp. Ther. Med. 2018, 15, 2667–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimaszewska-Wiśniewska, A.; Hałas-Wiśniewska, M.; Grzanka, A.; Grzanka, D. Evaluation of Anti-Metastatic Potential of the Combination of Fisetin with Paclitaxel on A549 Non-Small Cell Lung Cancer Cells. Int. J. Mol. Sci. 2018, 19, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimaszewska-Wisniewska, A.; Halas-Wisniewska, M.; Tadrowski, T.; Gagat, M.; Grzanka, D.; Grzanka, A. Paclitaxel and the dietary flavonoid fisetin: A synergistic combination that induces mitotic catastrophe and autophagic cell death in A549 non-small cell lung cancer cells. Cancer Cell Int. 2016, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Huang, Y.; Zhuo, W.; Zhu, Y.; Zhu, B.; Chen, Z. Fisetin, a dietary phytochemical, overcomes Erlotinib-resistance of lung adenocarcinoma cells through inhibition of MAPK and AKT pathways. Am. J. Transl. Res. 2016, 8, 4857–4868. [Google Scholar]
- Kang, K.A.; Piao, M.J.; Madduma Hewage, S.R.; Ryu, Y.S.; Oh, M.C.; Kwon, T.K.; Chae, S.; Hyun, J.W. Fisetin induces apoptosis and endoplasmic reticulum stress in human non-small cell lung cancer through inhibition of the MAPK signaling pathway. Tumour Biol. 2016, 37, 9615–9624. [Google Scholar] [CrossRef]
- Islami, F.; Sauer, A.G.; Miller, K.D.; Fedewa, S.A.; Minihan, A.K.; Geller, A.C.; Lichtenfeld, J.L.; Jemal, A. Cutaneous Melanomas Attributable to Ultraviolet Radiation Exposure by State. Int. J. Cancer 2020, 147, 1385–1390. [Google Scholar] [CrossRef]
- Moolakkadath, T.; Aqil, M.; Ahad, A.; Imam, S.S.; Praveen, A.; Sultana, Y.; Mujeeb, M.; Iqbal, Z. Fisetin loaded binary ethosomes for management of skin cancer by dermal application on UV exposed mice. Int. J. Pharm. 2019, 560, 78–91. [Google Scholar] [CrossRef]
- Imtiyaz, K.; Rahmani, A.H.; Alsahli, M.A.; Almatroodi, S.A.; Alam Rizvi, M.M. Fisetin induces apoptosis in human skin cancer cells through downregulating MTH1. J. Biomol. Struct. Dyn. 2022, 1–15. [Google Scholar] [CrossRef]
- Pal, H.C.; Athar, M.; Elmets, C.A.; Afaq, F. Fisetin Inhibits UVB-induced Cutaneous Inflammation and Activation of PI3K/AKT/NFκB Signaling Pathways in SKH-1 Hairless Mice. Photochem. Photobiol. 2014, 91, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.-H.; Kuo, C.-L.; Lu, K.-W.; Yu, F.-S.; Ma, Y.-S.; Yang, J.-L.; Chu, Y.-L.; Chueh, F.-S.; Liu, K.-C.; Chung, J.-G. Fisetin-induced apoptosis of human oral cancer SCC-4 cells through reactive oxygen species production, endoplasmic reticulum stress, caspase-, and mitochondria-dependent signaling pathways. Environ. Toxicol. 2017, 32, 1725–1741. [Google Scholar] [CrossRef]
- Shih, Y.L.; Hung, F.M.; Lee, C.H.; Yeh, M.Y.; Lee, M.H.; Lu, H.F.; Chen, Y.L.; Liu, J.Y.; Chung, J.G. Fisetin Induces Apoptosis of HSC3 Human Oral Cancer Cells Through Endoplasmic Reticulum Stress and Dysfunction of Mitochondria-mediated Signaling Pathways. In Vivo 2017, 31, 1103–1114. [Google Scholar] [PubMed] [Green Version]
- Li, Y.-S.; Qin, X.-J.; Dai, W. Fisetin suppresses malignant proliferation in human oral squamous cell carcinoma through inhibition of Met/Src signaling pathways. Am. J. Transl. Res. 2017, 9, 5678–5683. [Google Scholar] [PubMed]
- Li, R.; Liang, H.Y.; Li, M.Y.; Lin, C.Y.; Shi, M.J.; Zhang, X.J. Interference of fisetin with targets of the nuclear factor-κB signal transduction pathway activated by Epstein-Barr virus encoded latent membrane protein 1. Asian Pac. J. Cancer Prev. 2014, 15, 9835–9839. [Google Scholar] [CrossRef] [Green Version]
- Girardi, T.; Vicente, C.; Cools, J.; De Keersmaecker, K. The genetics and molecular biology of T-ALL. Blood 2017, 129, 1113–1123. [Google Scholar] [CrossRef] [Green Version]
- Ash, D.; Subramanian, M.; Surolia, A.; Shaha, C. Nitric oxide is the key mediator of death induced by fisetin in human acute monocytic leukemia cells. Am. J. Cancer Res. 2015, 5, 481–497. [Google Scholar]
- Chiu, S.-J.; Yang, P.-M.; Tseng, H.-H.; Peng, C.-W.; Chen, W.-S. Dietary flavonoid fisetin targets caspase-3-deficient human breast cancer MCF-7 cells by induction of caspase-7-associated apoptosis and inhibition of autophagy. Int. J. Oncol. 2011, 40, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Noh, E.-M.; Park, Y.-J.; Kim, J.-M.; Kim, M.-S.; Kim, H.-R.; Song, H.-K.; Hong, O.-Y.; So, H.-S.; Yang, S.-H.; Kim, J.-S.; et al. Fisetin regulates TPA-induced breast cell invasion by suppressing matrix metalloproteinase-9 activation via the PKC/ROS/MAPK pathways. Eur. J. Pharmacol. 2015, 764, 79–86. [Google Scholar] [CrossRef]
- Smith, M.L.; Murphy, K.; Doucette, C.D.; Greenshields, A.L.; Hoskin, D.W. The Dietary Flavonoid Fisetin Causes Cell Cycle Arrest, Caspase-Dependent Apoptosis, and Enhanced Cytotoxicity of Chemotherapeutic Drugs in Triple-Negative Breast Cancer Cells. J. Cell. Biochem. 2016, 117, 1913–1925. [Google Scholar] [CrossRef]
- Guo, G.; Zhang, W.; Dang, M.; Yan, M.; Chen, Z. Fisetin induces apoptosis in breast cancer MDA-MB-453 cells through degradation of HER2/neu and via the PI3K/Akt pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22268. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Dong, X.; Shi, G. In vitro and in vivo Antitumor Effects of Fisetin and Fisetin Nanopartical on Ovarian Cancer. J. Sichuan Univ. Med. Sci. Ed. 2018, 49, 551–555. [Google Scholar]
- Meng, Y.-B.; Xiao, C.; Chen, X.-L.; Bai, P.; Yao, Y.; Wang, H.; Xiao, X. The Antitumor Effects of Fisetin on Ovarian Cancer in vitro and in vivo. J. Sichuan Univ. Med. Sci. Ed. 2016, 47, 830–836. [Google Scholar]
- Xiao, X.; Zou, J.; Fang, Y.; Meng, Y.; Xiao, C.; Fu, J.; Liu, S.; Bai, P.; Yao, Y. Fisetin and polymeric micelles encapsulating fisetin exhibit potent cytotoxic effects towards ovarian cancer cells. BMC Complement. Altern. Med. 2018, 18, 91. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.T.; Lin, C.L.; Lin, T.Y.; Cheng, C.W.; Yang, S.F.; Lin, C.L.; Wu, C.C.; Hsieh, Y.H.; Tsai, J.P. Synergistic effect of fisetin combined with sorafenib in human cervical cancer HeLa cells through activation of death receptor-5 mediated caspase-8/caspase-3 and the mitochondria-dependent apoptotic pathway. Tumour Biol. 2016, 37, 6987–6996. [Google Scholar] [CrossRef]
- Ying, T.H.; Yang, S.F.; Tsai, S.J.; Hsieh, S.C.; Huang, Y.C.; Bau, D.T.; Hsieh, Y.H. Fisetin induces apoptosis in human cervical cancer HeLa cells through ERK1/2-mediated activation of caspase-8-/caspase-3-dependent pathway. Arch Toxicol. 2012, 86, 263–273. [Google Scholar] [CrossRef]
- Chou, R.H.; Hsieh, S.C.; Yu, Y.L.; Huang, M.H.; Huang, Y.C.; Hsieh, Y.H. Fisetin inhibits migration and invasion of human cervical cancer cells by down-regulating urokinase plasminogen activator expression through suppressing the p38 MAPK-dependent NF-κB signaling pathway. PLoS ONE. 2013, 8, e71983. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, W.; Wang, S.; Wei, Z. Fisetin induces G2/M phase cell cycle arrest by inactivating cdc25C-cdc2 via ATM-Chk1/2 activation in human endometrial cancer cells. Bangladesh. J. Pharmacol. 2015, 10, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.K.; Peng, S.F.; Lai, K.C.; Liu, H.C.; Huang, Y.P.; Lin, C.C.; Huang, A.C.; Chueh, F.S.; Chung, J.G. Fistein Suppresses Human Osteosarcoma U-2 OS Cell Migration and Invasion via Affecting FAK, uPA and NF-ĸB Signaling Pathway In Vitro. In Vivo 2019, 33, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-M.; Li, W.-Y.; Huang, M.-Y.; Zhang, X.-Q. Fisetin, a dietary flavonoid induces apoptosis via modulating the MAPK and PI3K/Akt signalling pathways in human osteosarcoma (U-2 OS) cells. Bangladesh J. Pharmacol. 2015, 10, 820–829. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, J.M.P.F.; Pacheco, A.R.; Coutinho, L.; Oliveira, H.; Pinho, S.; Almeida, L.; Fernandes, E.; Santos, C. Combination of etoposide and fisetin results in anti-cancer efficiency against osteosarcoma cell models. Arch. Toxicol. 2018, 92, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.; Hsieh, Y.H.; Hwang, J.M.; Jan, H.J.; Hsieh, S.C.; Lin, S.H.; Lai, C.Y. Fisetin suppresses ADAM9 expression and inhibits invasion of glioma cancer cells through increased phosphorylation of ERK1/2. Tumour Biol. 2015, 36, 3407–3415. [Google Scholar] [CrossRef] [PubMed]
- Pak, F.; Oztopcu-Vatan, P. Fisetin effects on cell proliferation and apoptosis in glioma cells. Z. Nat. C J. Biosci. 2019, 74, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Kivelä, T. The epidemiological challenge of the most frequent eye cancer: Retinoblastoma, an issue of birth and death. Br. J. Ophthalmol. 2009, 93, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Lee, J.Y.; Byun, B.J.; Kim, S.H. Fisetin targets phosphatidylinositol-3-kinase and induces apoptosis of human B lymphoma Raji cells. Toxicol. Rep. 2015, 2, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics, 2014. CA Cancer J Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Kraehe, P.; Popper, B.; Shayan, P.; Goel, A.; Buhrmann, C. Curcumin potentiates antitumor activity of 5-fluorouracil in a 3D alginate tumor microenvironment of colorectal cancer. BMC Cancer 2015, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.B.; Shakibaei, M. Resveratrol Chemosensitizes TNF-β-Induced Survival of 5-FU-Treated Colorectal Cancer Cells. Nutrients 2018, 10, 888. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jiang, P.; Wang, P.; Yang, C.S.; Wang, X.; Feng, Q. EGCG Enhances Cisplatin Sensitivity by Regulating Expression of the Copper and Cisplatin Influx Transporter CTR1 in Ovary Cancer. PLoS ONE. 2015, 10, e0125402. [Google Scholar]
- Tripathi, R.; Samadder, T.; Gupta, S.; Surolia, A.; Shaha, C. Anticancer Activity of a Combination of Cisplatin and Fisetin in Embryonal Carcinoma Cells and Xenograft Tumors. Mol. Cancer Ther. 2011, 10, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Jafarzadeh, S.; Baharara, J.; Tehranipour, M. Apoptosis Induction with Combined Use of Cisplatin and Fisetin in Cisplatin- Resistant Ovarian Cancer Cells (A2780). Avicenna J. Med. Biotechnol. 2021, 13, 176. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Zhang, L.; Zhu, Y.; Zhu, B.; Chen, Z. Fisetin, a dietary bioflavonoid, reverses acquired Cisplatin-resistance of lung adenocarcinoma cells through MAPK/Survivin/Caspase pathway. Am. J. Transl. Res. 2015, 7, 2045–2052. [Google Scholar] [PubMed]
- Mukhtar, E.; Adhami, V.M.; Siddiqui, I.A.; Verma, A.K.; Mukhtar, H. Fisetin Enhances Chemotherapeutic Effect of Cabazitaxel against Human Prostate Cancer Cells. Mol. Cancer Ther. 2016, 15, 2863–2874. [Google Scholar] [CrossRef] [Green Version]
- Pal, H.C.; Diamond, A.C.; Strickland, L.R.; Kappes, J.C.; Katiyar, S.K.; Elmets, C.A.; Athar, M.; Afaq, F. Fisetin, a dietary flavonoid, augments the anti-invasive and anti-metastatic potential of sorafenib in melanoma. Oncotarget 2016, 7, 1227–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Jajeh, F.; Eberhardt, E.L.; Miller, D.D.; Albrecht, D.M.; Van Doorn, R.; Hruby, M.D.; Maresh, M.E.; Clipson, L.; Mukhtar, H.; et al. Fisetin and 5-fluorouracil: Effective combination for PIK3CA-mutant colorectal cancer. Int. J. Cancer 2019, 145, 3022–3032. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-S.; Lien, G.-S.; Shen, S.-C.; Yang, L.-Y.; Chen, Y.-C. HSP90 Inhibitors, Geldanamycin and Radicicol, Enhance Fisetin-Induced Cytotoxicity via Induction of Apoptosis in Human Colonic Cancer Cells. Evidence-Based Complement. Altern. Med. 2013, 2013, 987612. [Google Scholar] [CrossRef]
- de Oliveira, J.F.; Oliveira, H.; Pinho, S.; Pimentel, F.; Almeida, L.; Van Zoelen, E.; Santos, C. Cytotoxic and genotoxic activity of fisetin (3, 3′, 4′, 7-tetrahydroxyflavone) in an osteosarcoma in vitro model. Planta Med. 2014, 80, P1L4. [Google Scholar] [CrossRef]
- Seguin, J.; Brullé, L.; Boyer, R.; Lu, Y.M.; Ramos Romano, M.; Touil, Y.S.; Scherman, D.; Bessodes, M.; Mignet, N.; Chabot, G.G. Liposomal encapsulation of the natural flavonoid fisetin improves bioavailability and antitumor efficacy. Int. J. Pharm. 2013, 444, 146–154. [Google Scholar] [CrossRef]
- Bothiraja, C.; Yojana, B.D.; Pawar, A.P.; Shaikh, K.S.; Thorat, U.H. Fisetin-loaded nanocochleates: Formulation, characterisation, in vitro anticancer testing, bioavailability and biodistribution study. Expert Opin. Drug Deliv. 2014, 11, 17–29. [Google Scholar] [CrossRef]
- Touil, Y.S.; Auzeil, N.; Boulinguez, F.; Saighi, H.; Regazzetti, A.; Scherman, D.; Chabot, G.G. Fisetin disposition and metabolism in mice: Identification of geraldol as an active metabolite. Biochem. Pharmacol. 2011, 82, 1731–1739. [Google Scholar] [CrossRef] [Green Version]
- Shia, C.-S.; Tsai, S.-Y.; Kuo, S.-C.; Hou, Y.-C.; Chao, P.-D.L. Metabolism and Pharmacokinetics of 3,3′,4′,7-Tetrahydroxyflavone (Fisetin), 5-Hydroxyflavone, and 7-Hydroxyflavone and Antihemolysis Effects of Fisetin and Its Serum Metabolites. J. Agric. Food Chem. 2009, 57, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.-C.; Hsueh, T.Y.; Cheng, Y.-Y.; Lin, L.-C.; Tsai, T.-H. Pharmacokinetics and Biliary Excretion of Fisetin in Rats. J. Agric. Food Chem. 2018, 66, 6300–6307. [Google Scholar] [CrossRef] [PubMed]
- Sechi, M.; Syed, D.N.; Pala, N.; Mariani, A.; Marceddu, S.; Brunetti, A.; Mukhtar, H.; Sanna, V. Nanoencapsulation of dietary flavonoid fisetin: Formulation and in vitro antioxidant and α-glucosidase inhibition activities. Mater. Sci. Eng. C 2016, 68, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Kadari, A.; Gudem, S.; Kulhari, H.; Bhandi, M.M.; Borkar, R.M.; Kolapalli, V.R.M.; Sistla, R. Enhanced oral bioavailability and anticancer efficacy of fisetin by encapsulating as inclusion complex with HPβCD in polymeric nanoparticles. Drug Deliv. 2017, 24, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Ragelle, H.; Crauste-Manciet, S.; Seguin, J.; Brossard, D.; Scherman, D.; Arnaud, P.; Chabot, G.G. Nanoemulsion formulation of fisetin improves bioavailability and antitumour activity in mice. Int. J. Pharm. 2012, 427, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.X.; Ge, C.X.; Li, Q.; Lou, D.S.; Hu, L.F.; Sun, Y.; Xiong, M.X.; Lai, L.L.; Zhong, S.Y.; Yi, C.; et al. Fisetin nanoparticles protect against PM2. 5 exposure-induced neuroinflammation by down-regulation of astrocytes activation related NF-κB signaling pathway. J. Funct. Foods 2020, 65, 103716. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, K.H. Method for Preparing Rhus Verniciflua Stokes Extract Containing Increased Fisetin Content, and Metastasis-Inhibiting Anticancer Agent Composition Containing Same. US2021038667A1, 5 July 2022. [Google Scholar]
- Deguan, D.H.L.; Dong, Y.; Wang, M.; Zhang, Y.; Wu, J. Application of Fisetin and Salt Thereof to Preparation of Drugs Resisting Radiation Damages, 2020. CN111700888A, 25 September 2020. [Google Scholar]
- Hyung, J.M.; Jai, C.H.; Woo, L.J. Composition for Preventing or Treating Uterine Myoma Comprising Fisetin from Rhus Verniciflua Stokes Extract, 2020. KR20200027344A, 15 July 2020. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathways/Gene | Anticancer Mechanism | Refs. |
|---|---|---|
| Inflammation | Plasma levels of hs-CRP as well as interlukin-8 decreased, and the fisetin supplement reduced the values of MMP-7 levels. Fisetin improved the inflammatory status in cancer patients. | [34] |
| Apoptosis | Treatment with fisetin induced a sub-G1 population in a dose-dependent way. Additionally, the cleavage of poly(ADP-ribose) polymerase (PARP)—a substrate of caspase as well as a marker of apoptosis—was also enhanced. Fisetin caused morphological changes, followed by cell membrane blebbing and shrinkage. | [38] |
| Fisetin reduced the mitochondrial membrane potential and the proapoptotic members Bak and Bax and activated caspase-3 and PARP. | [39] | |
| Fisetin downregulated anti-apoptotic genes and upregulated pro-apoptotic genes. The expression of multiple receptors and ligands involved in extrinsic pathways increased. | [40] | |
| Fisetin inhibited antiapoptotic Bcl-2 family proteins as well as damaged the mitochondrial transmembrane potential. | [41] | |
| Autophagy | Fisetin induced autophagy, upregulated the autophagy marker LC3B and increased the autophagic flux in pancreatic cancer cells. | [45] |
| Fisetin treatment induces the development of autophagic vacuoles in oral cancer cells. Fisetin-induced autophagy in cancer cells was observed via numerous autophagy markers. | [39] | |
| Angiogenesis | Fisetin affected the expression of VEGFR, and this effect was in a dose-dependent manner. Consequently, fisetin downregulated the VEGFR expression. | [47] |
| Fisetin inhibited capillary-like tube formation, which was linked with a decreased expression of vascular endothelial growth factor and endothelial nitric oxide synthase. | [48] | |
| Fisetin treatment caused a dose-dependent decrease in Matrigel plug hemoglobin levels and a decrease in tumor angiogenesis. | [49] | |
| Cell cycle | Fisetin treatment played a role in the cell cycle arrest at the G0/G1 phase. The sub-G1 group meaningfully increased after the treatment of fisetin. | [51] |
| Fisetin decreased the total viable cells via G0/G1 phase arrest and induced the sub-G1 phase. | [52] | |
| Fisetin as well as hesperetin treatment caused a concentration- and time-dependent inhibition of proliferation and induced G2/M arrest. | [53] | |
| PI3K/AKT/ mTOR | Fisetin showed a potential role in the regulation of cancer via inducing apoptosis and regulated autophagy through AKT/NF-κB/mTOR signaling pathways. | [54] |
| Fisetin suppress the growth, invasion and migration of pancreatic cancer cells through reducing the PI3K/AKT/mTOR cascade. | [55] | |
| p-Akt and p-Akt/Akt, p-PI3K and p-PI3K/PI3K and p-mTOR decreased, upregulated Bax and downregulated Bcl-xL after fisetin treatment. | [57] | |
| STAT3 | Fisetin downregulated the JAK 1 and STAT3 expression in cancer cells, and fisetin induced apoptosis. | [63] |
| Wnt/ beta-catenin | Fisetin treatment reduced the levels of Wnt-target genes including cyclin D1 and MMP7. | [68] |
| Increasing doses of fisetin were linked with a decrease in cytosolic β-catenin, with a concomitant reduction in nuclear β-catenin. | [69] | |
| Nuclear factor-κB | Nuclear factor-κB was accumulated in the cytoplasm, and the protein expression of NF-κB in the nuclei was decreased by fisetin treatment. | [51] |
| Fisetin inhibited the enhancement of IKK, causing a reduction in the stimulation of the NF-κB signaling pathway. | [71] | |
| Nrf2 | Fisetin translocated Nrf2 into the nucleus, and the expression of the downstream HO-1 gene was upregulated via the inhibition of the Nrf2 degradation at the post-transcriptional level. | [75] |
| Fisetin-induced HO-1 protein expression was reduced through silencing Nrf2. | [76] | |
| JNK/ERK/AP-1 | Fisetin upregulated ZAK expression to induce the Hippo pathway and mediated the activation of JNK/ERK to trigger cell apoptosis in an AP-1-dependent manner | [79] |
| Cancer | Outcome of the Study | Refs. |
|---|---|---|
| Prostate cancer | Fisetin is a potent HA synthesis inhibitor which increases the abundance of antiangiogenic HMM-HA and could be used for the management of prostate cancer. | [82] |
| The treatment of cancer cells with fisetin showed a high expression of acetylated α-tubulin in a dose-dependent way. | [83] | |
| Fisetin sensitizes the tumor necrosis factor-related apoptosis-inducing ligand-resistant androgen-dependent LNCaP as well as the androgen-independent prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand-induced death. | [84] | |
| Fisetin acts as a dual inhibitor of mTORC1/2 signaling, leading to the induction of autophagic cell death and the inhibition of Cap-dependent translation in cancer cells. | [85] | |
| Kidney cancer | Fisetin induced a sub-G1 population in a dose-dependent fashion and caused cell shrinkage and membrane blebbing. Cancer cells were highly sensitive, compared to normal cells, to fisetin treatment. | [38] |
| Fisetin inhibited the migration and invasion of cancer cells via the downregulation of CTSS, metalloproteinase 9 and disintegrin. | [87] | |
| Liver cancer | Fisetin inhibited autophagy via the activation of PI3K/Akt/mTOR and the modulation of the AMPK signaling pathways. | [89] |
| Fisetin displayed a suggestively inhibitory role in cancer cell proliferation. Likewise, the death receptor 2 agonist of bromocriptine absolutely blocked cancer cell proliferation in a dose-dependent fashion. | [90] | |
| Colon cancer | Fisetin dose-dependently blocked both DNA synthesis and cell growth and disturbed cell cycle progression. | [91] |
| Fisetin treatment efficiently inhibited the cell viability and apoptosis of CPT11-LoVo cells compared to Oxaliplatin and parental LoVo cancer cells. Moreover, apoptosis was encouraged by fisetin treatment, endorsing Caspase-8 and Cytochrome-C expressions. | [92] | |
| Fisetin caused an increase in the levels of proapoptotic Bim and Bak and induced a reduction in the protein levels of antiapoptotic Bcl-2 and Bcl-xL. | [93] | |
| Securin reduction sensitizes human colon cancer cells to fisetin-induced apoptosis. | [94] | |
| Gastric cancer | Fisetin substantially decreases G1 phase cyclins and CDKs levels, and the levels of p53 increased. | [95] |
| Fisetin treatment with various concentrations suggestively decreased the proliferation rate of SGC7901 cells. | [96] | |
| Pancreatic cancer | Combination treatment with gemcitabine and fisetin inhibited the proliferation of cancer cells and played role in the induction of apoptosis, as indicated by caspase 3/7 activation. | [97] |
| PANC-1 cells were cultured with different concentrations of fisetin, and the findings exhibited that fisetin reduced PANC-1 cell viability in a dose- and time-dependent way. | [45] | |
| The transient downregulation of DR3 by RNA interference meaningfully increased fisetin-induced changes in cell proliferation, cell invasion and apoptosis. | [99] | |
| Bile duct cancer | Fisetin caused the inhibition of the survival of cancer cells. It also encouraged cellular apoptosis additively in combination with gemcitabine. | [100] |
| Bladder cancer | Fisetin-induced apoptosis in bladder cancer is initiated through the regulation of two associated pathways: the downregulation of the NF-κB pathway and the upregulation of p53 activity. | [101] |
| Fisetin inhibited the proliferation of EJ and T24 cells via blocking cell cycle progression in the G0/G1 phase and inducing apoptosis. | [51] | |
| Lung cancer | Fisetin suppressed proliferation, adhesion, migration and invasion. The induction of apoptosis was noticed after fisetin treatment via decreasing the expression of cyclin-D and c-myc. | [103] |
| Fisetin showed some protection against paclitaxel-mediated cytotoxicity. | [104] | |
| Fisetin efficiently enhanced the sensitivity of erlotinib-resistant cancer cells to erlotinib, possibly via inhibiting the abnormal enhancement of AKT as well as MAPK signaling pathways. | [106] | |
| Fisetin-treated cells demonstrated a dose-dependent prevention of the constituents of the mTOR signaling complex. | [56] | |
| Skin cancer | Mice previously treated with fisetin binary ethosomes gel showed a noticeable decrease in the levels of the pro-inflammatory marker and a lower percentage of tumor incidences. | [109] |
| Fisetin inhibited IKK activation, leading to a reduction in the enhancement of the Nuclear Factor-κB signaling pathway. | [71] | |
| The treatment of fisetin decreases cell proliferation markers and DNA damage, as demonstrated by the increased expression of p21 and p53 proteins. | [111] | |
| Oral cancer | Fisetin induced cell death, G2/M phase arrest and the induction of apoptosis and decreased the level of mitochondria membrane potential. | [112] |
| Fisetin induced apoptotic cell death via increased reactive oxygen species and Ca2+. Moreover, fisetin played a role in the increase in caspase-8, -9 and -3 activities as well as reduced the mitochondrial membrane potential. | [113] | |
| Fisetin expressively inhibits tumor cell proliferation and induces apoptosis in oral squamous cell carcinoma. | [114] | |
| Fisetin played a role in the inhibition of the enhancement of PI3K/AKT-controlled mTOR. | [54] | |
| Leukemia | Fisetin was able to kill THP-1 cells in vivo, resulting in tumor shrinkage in the xenograft mouse model. Death induction in vitro was facilitated by an increase in the nitric, causing the activation of both the extrinsic and the intrinsic apoptotic pathways. | [117] |
| Breast cancer | Fisetin-induced apoptosis was revoked by z-VAD-fmk, a pan-caspase inhibitor. Additionally, the inhibition of autophagy by using fisetin was revealed as an extra route to speedy anticancer activity in MCF-7 cells. | [118] |
| Fisetin suggestively decreased 2-O-tetradecanoylphorbol-13-acetate-caused cell invasion in breast cancer cells, and it was also found to inhibit the activation of the p38 MAPK. | [119] | |
| Fisetin inhibited breast cancer cells division and also played a role in the induction of apoptosis. | [120] | |
| Fisetin enhanced tumor cell apoptosis and decreased the growth of 4T1 cell-derived orthotopic breast tumors. | [57] | |
| Ovarian cancer | Fisetin as well as fisetin nanoparticles showed an inhibitory effect on ovarian cancer cells in a dose-dependent way. | [122] |
| In a rat model study, the tumor mass and tumor volume were meaningfully reduced under the effect of fisetin. A greater concentration of fisetin caused a substantial increase in Bax and a decrease in Bcl-2. | [123] | |
| Cell numbers were reduced after fisetin/fisetin micelles treatment in a time- and dose-dependent fashion. | [124] | |
| Cervix cancer | Synergistic apoptosis was induced by the combination of fisetin and sorafenib in cancer cells. | [125] |
| Fisetin induced the apoptosis of cervical cancer (HeLa) cells in a dose- and time-dependent fashion. | [126] | |
| SiHa cells treated with fisetin concentrations of 20 and 40 µM exhibited a decrease in motility of 46.0% and 81.3%, respectively, and similar findings were also seen in CaSki cells, with 62.1% and 90.2% of inhibition. | [127] | |
| Endometrium cancer | Fisetin (20–100 µM) efficiently decreased the viability of KLE and Hec1 A cells and possibly changed the cell population at the G2/M stage. | [128] |
| Bone cancer | Fisetin suppressed migration, cell mobility and invasion and inhibited MMP-2 activity in U-2 OS cells. | [129] |
| Fisetin was related to a reduction in colony formation in U2OS and Saos-2 cells, whereas it was not in MG-63 and MG-63 cells, and Saos-2 cells showed a reduced cell proliferation at concentrations of 40 and 20 µM of fisetin, respectively. | [130] | |
| The ERK1/2 and PI3K/Akt pathways were inhibited by fisetin, and fisetin enhanced the expressions of p-c-Jun, p-JNK and p-p38. Fisetin caused reactive oxygen species (ROS) generation as well as a reduction in mitochondrial membrane potential. | [131] | |
| Brain cancer | Fisetin could be a possible therapeutic agent to counter human glioma cells based on its ability to activate ERK1/2 and to prevent the expression of ADAM9. | [132] |
| Fisetin was found to have cytotoxicity at lower doses in T98G cells as compared to a positive control. | [133] | |
| Retina cancer | Fisetin inhibited the migration and invasion of Y79 cells in a dose-dependent way. | [47] |
| Cancer of the lymphatic system | Fisetin (30 μM) treatment played role in the decreased protein expression of cIAP-2. | [135] |
| Thyroid cancer | Fisetin enhanced the expression of the caspase (-3, -8, and -9) and decreased the JAK 1 and STAT3 expression in cancer cells. | [63] |
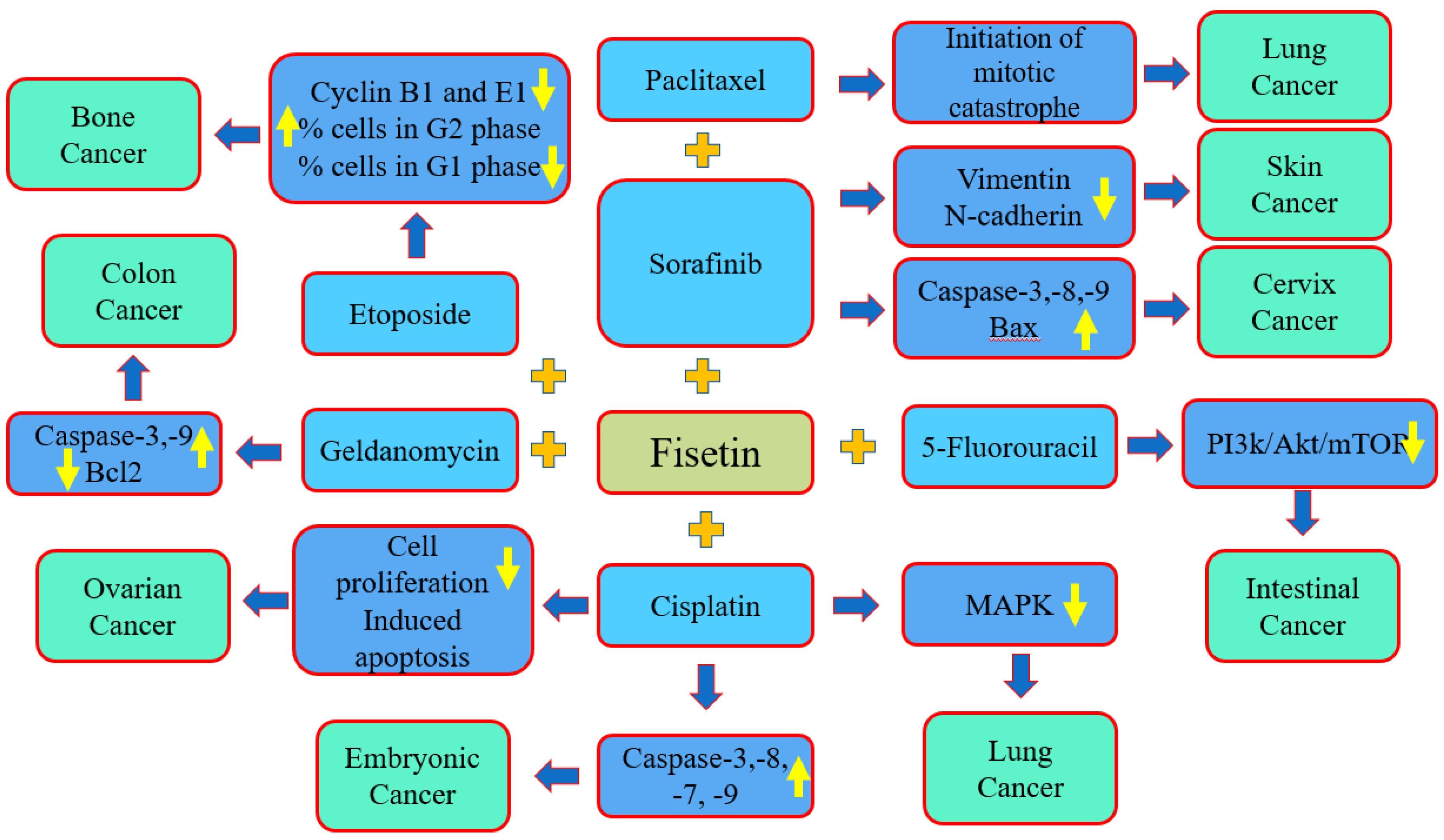
| Fisetin + Anticancer Drugs | Cancer | Finding of the Study | Refs. |
|---|---|---|---|
| Fisetin and sorafenib | Cervix cancer | The combination of sorafenib and fisetin synergistically caused the induction of apoptosis. The combined treatment of fisetin and sorafenib was obviously greater than sorafenib treatment alone based on the HeLa xenograft model. | [125] |
| Fisetin and cisplatin | Embryonal carcinoma | Fisetin enhanced the cytotoxicity of cisplatin. Fisetin and cisplatin increased cisplatin’s cytotoxicity at a dose that was 4 times lower than that needed by cisplatin monotherapy for the same effects of cytotoxicity. | [140] |
| Fisetin and cisplatin | Ovarian cancer | The combined treatment of Fisetin and cisplatin efficiently inhibits the proliferation of cancer cells, and the fragmentation of chromatin in cells occurred in the combination treatment. | [141] |
| Fisetin and cisplatin | Lung cancer | Fisetin efficiently increased the sensitivity of cancer cells to cisplatin, probably mediated via the inhibition of the abnormal activation of MAPK signaling pathways. | [142] |
| Fisetin and etoposide | Bone cancer | Fisetin:etoposide decreased the percentage of cells in the G1-phase and increased the percentage of cells in the G2-phase. | [131] |
| Fisetin and sorafenib | Skin cancer | The combination treatment (sorafenib + fisetin) more efficiently decreased the migration and invasion of BRAF-mutated melanoma cells. | [144] |
| Fisetin and 5-fluorouracil | Colon cancer | The treatment of PIK3CA-mutant cells with fisetin and 5-fluorouracil decreased the expression of PI3K, the phosphorylation of AKT and mTOR. Additionally, the combination of fisetin and 5-FU also decreased the total number of intestinal tumors. | [145] |
| Fisetin and geldanamycin | Colon cancer | Compared to fisetin treatment alone, geldanamycin and radicicol meaningfully increased the fisetin-induced cytotoxicity, produced a greater density of DNA ladder formation and enhanced the expression of cleaved caspase-3 as well as the PAPR protein | [146] |
| Fisetin and paclitaxel | Lung cancer | The switch from the cytoprotective autophagy to the autophagic cell death was concerned in the role of the synergistic action of both of the used compounds. | [105] |
| Fisetin and etoposide | Bone cancer | Cancer cells viability was decreased by 35% after exposure with fisetin. Fisetin showed synergistic cytotoxic effects with etoposide. | [147] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmani, A.H.; Almatroudi, A.; Allemailem, K.S.; Khan, A.A.; Almatroodi, S.A. The Potential Role of Fisetin, a Flavonoid in Cancer Prevention and Treatment. Molecules 2022, 27, 9009. https://doi.org/10.3390/molecules27249009
Rahmani AH, Almatroudi A, Allemailem KS, Khan AA, Almatroodi SA. The Potential Role of Fisetin, a Flavonoid in Cancer Prevention and Treatment. Molecules. 2022; 27(24):9009. https://doi.org/10.3390/molecules27249009
Chicago/Turabian StyleRahmani, Arshad Husain, Ahmad Almatroudi, Khaled S. Allemailem, Amjad Ali Khan, and Saleh A. Almatroodi. 2022. "The Potential Role of Fisetin, a Flavonoid in Cancer Prevention and Treatment" Molecules 27, no. 24: 9009. https://doi.org/10.3390/molecules27249009
APA StyleRahmani, A. H., Almatroudi, A., Allemailem, K. S., Khan, A. A., & Almatroodi, S. A. (2022). The Potential Role of Fisetin, a Flavonoid in Cancer Prevention and Treatment. Molecules, 27(24), 9009. https://doi.org/10.3390/molecules27249009