
Microplastics (Polystyrene) Exposure Induces Metabolic Changes in the Liver of Rare Minnow (Gobiocypris rarus)

Abstract
:1. Introduction
2. Results
2.1. Condition during This Study
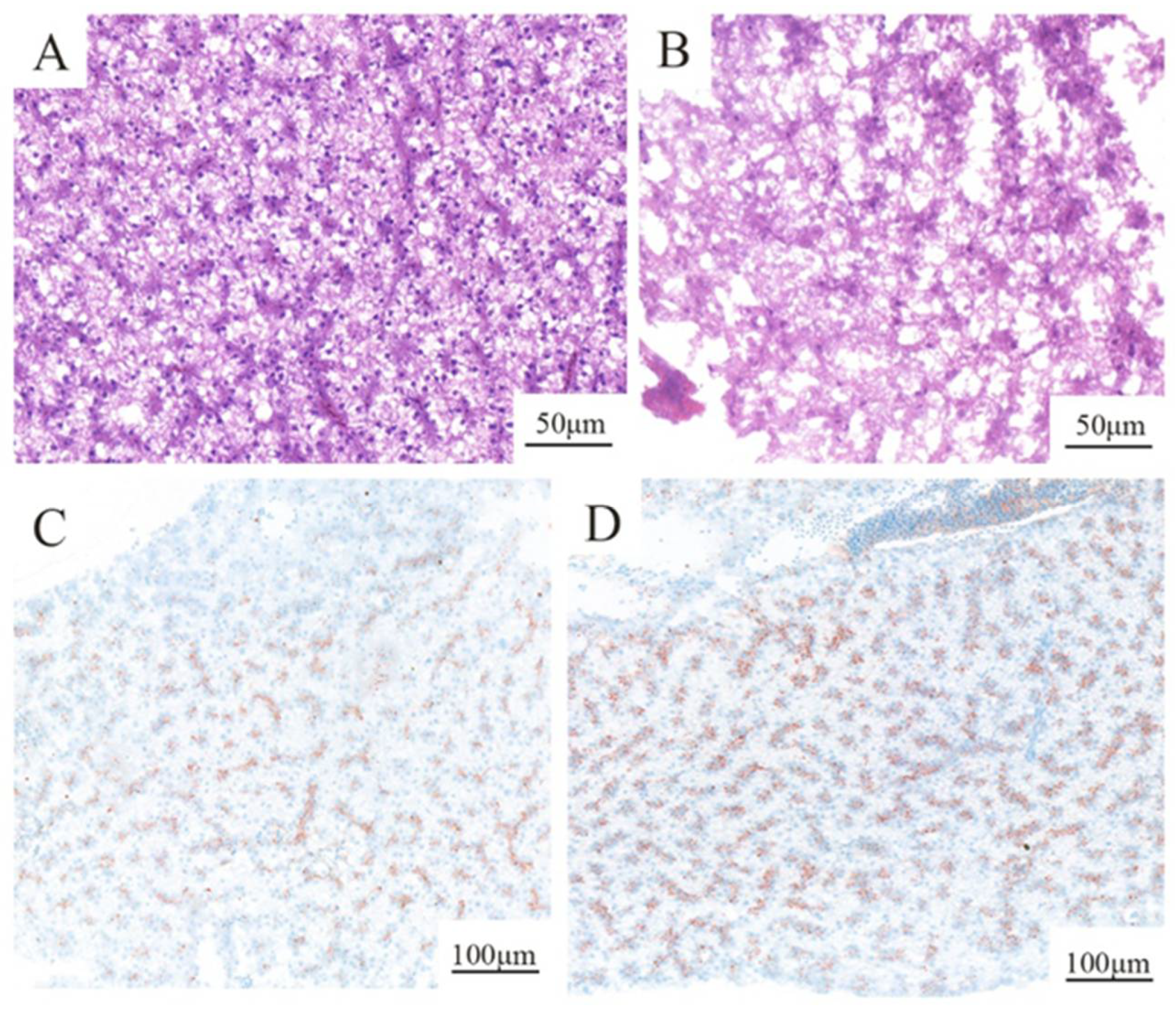
2.2. Histopathological Changes Induced by PS-MPs Treatment
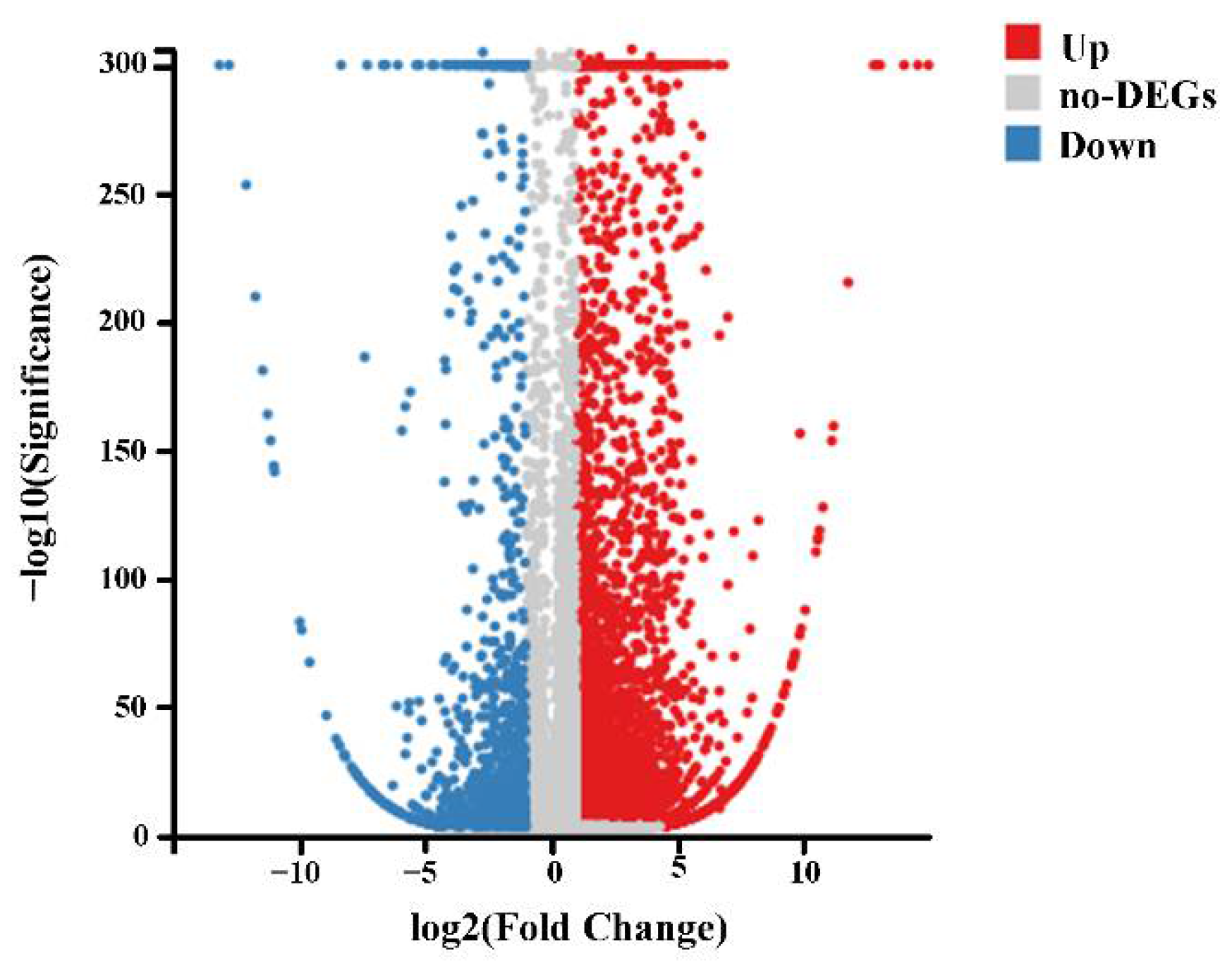
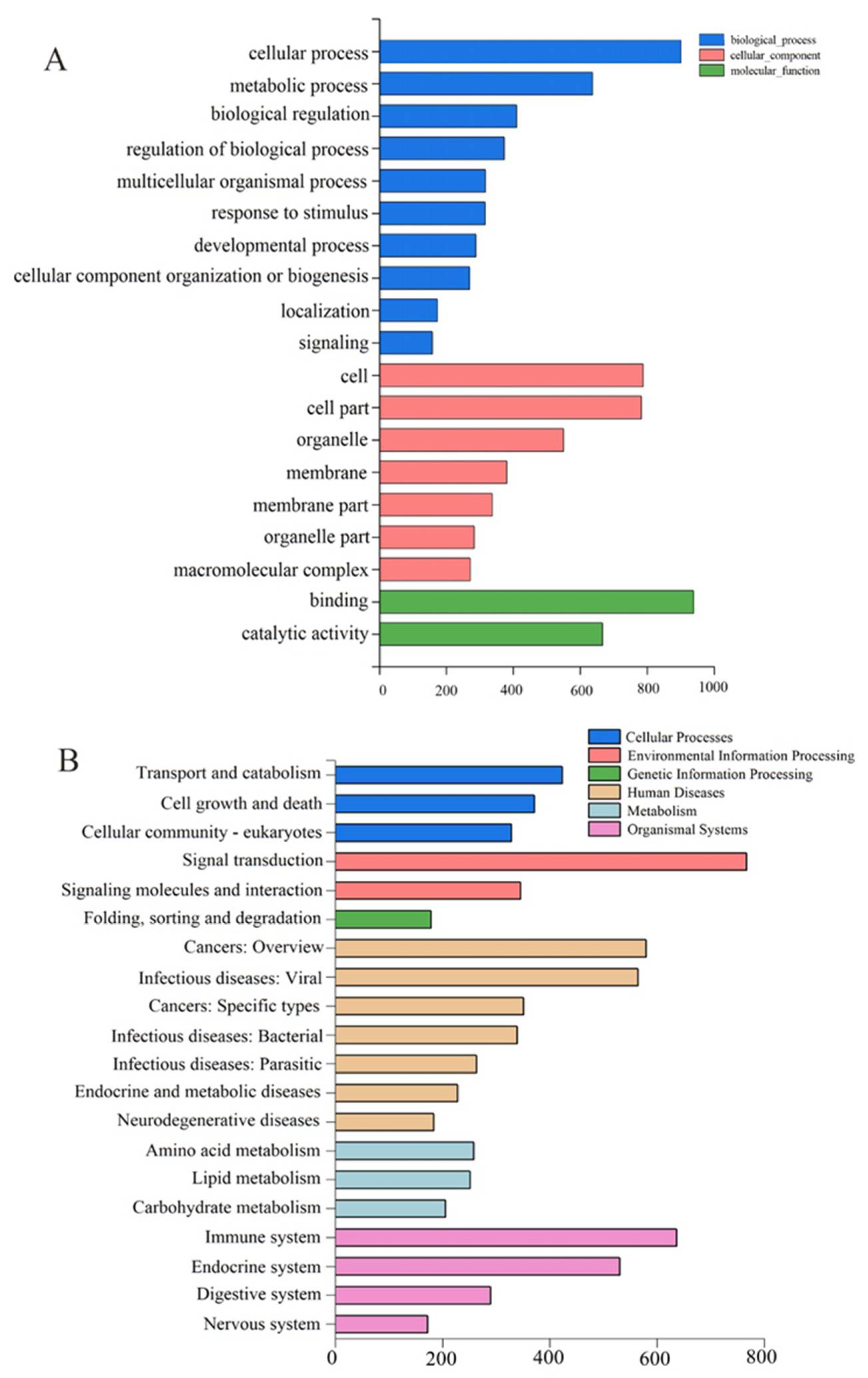
2.3. Transcriptome Analysis
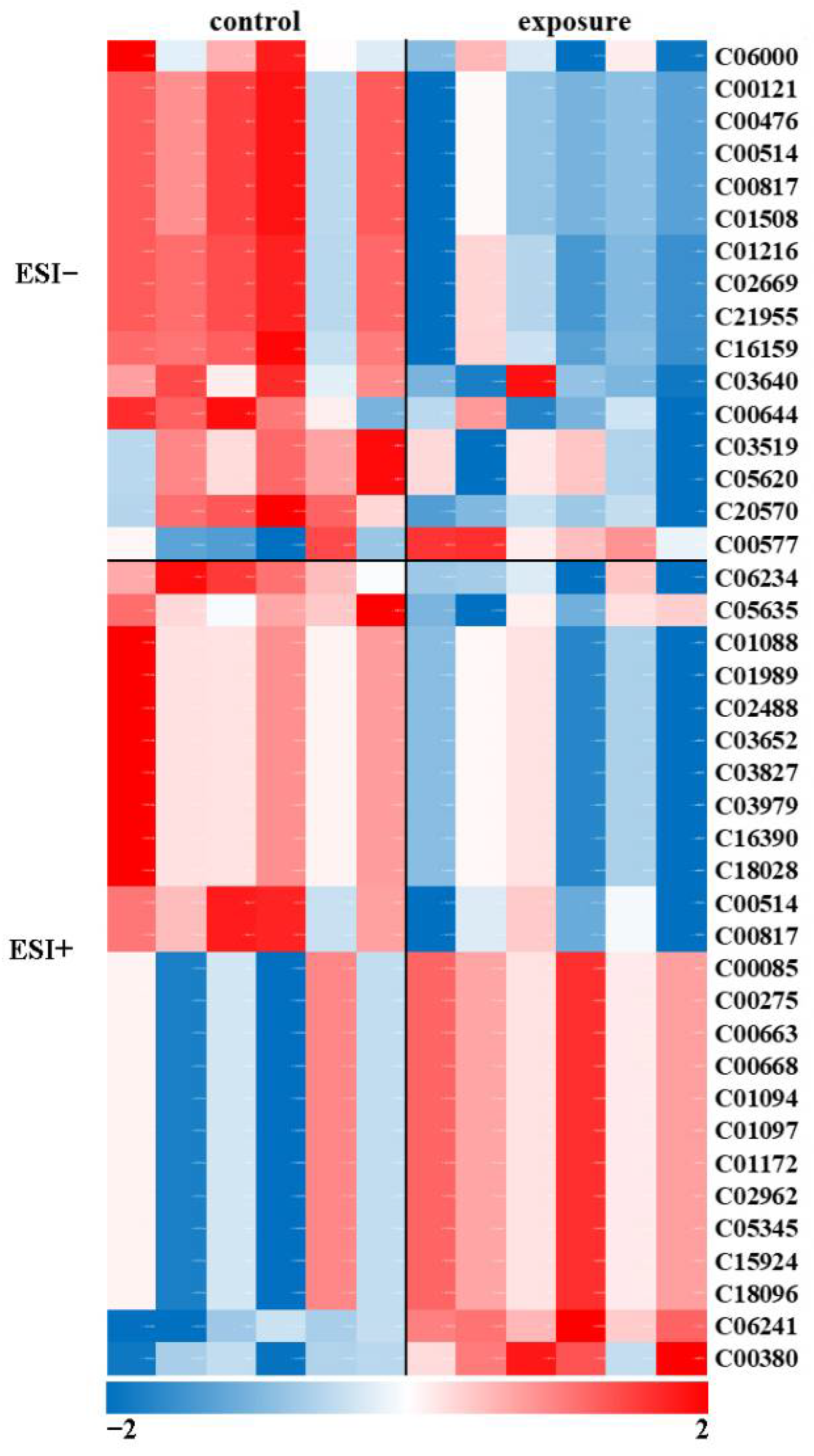
2.4. Metabolomic Analysis
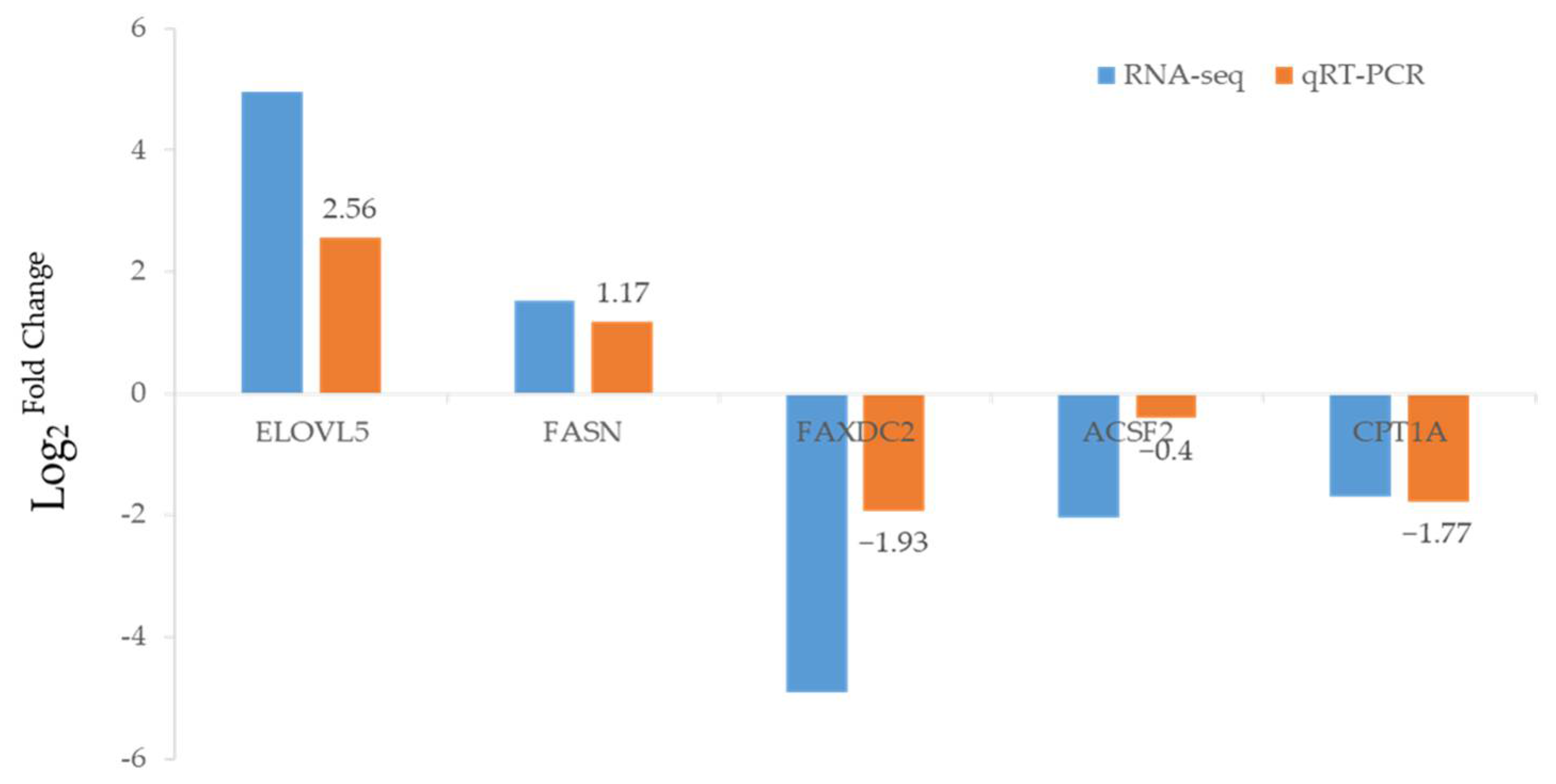
2.5. Confirmation of DEGs by qRT-PCR
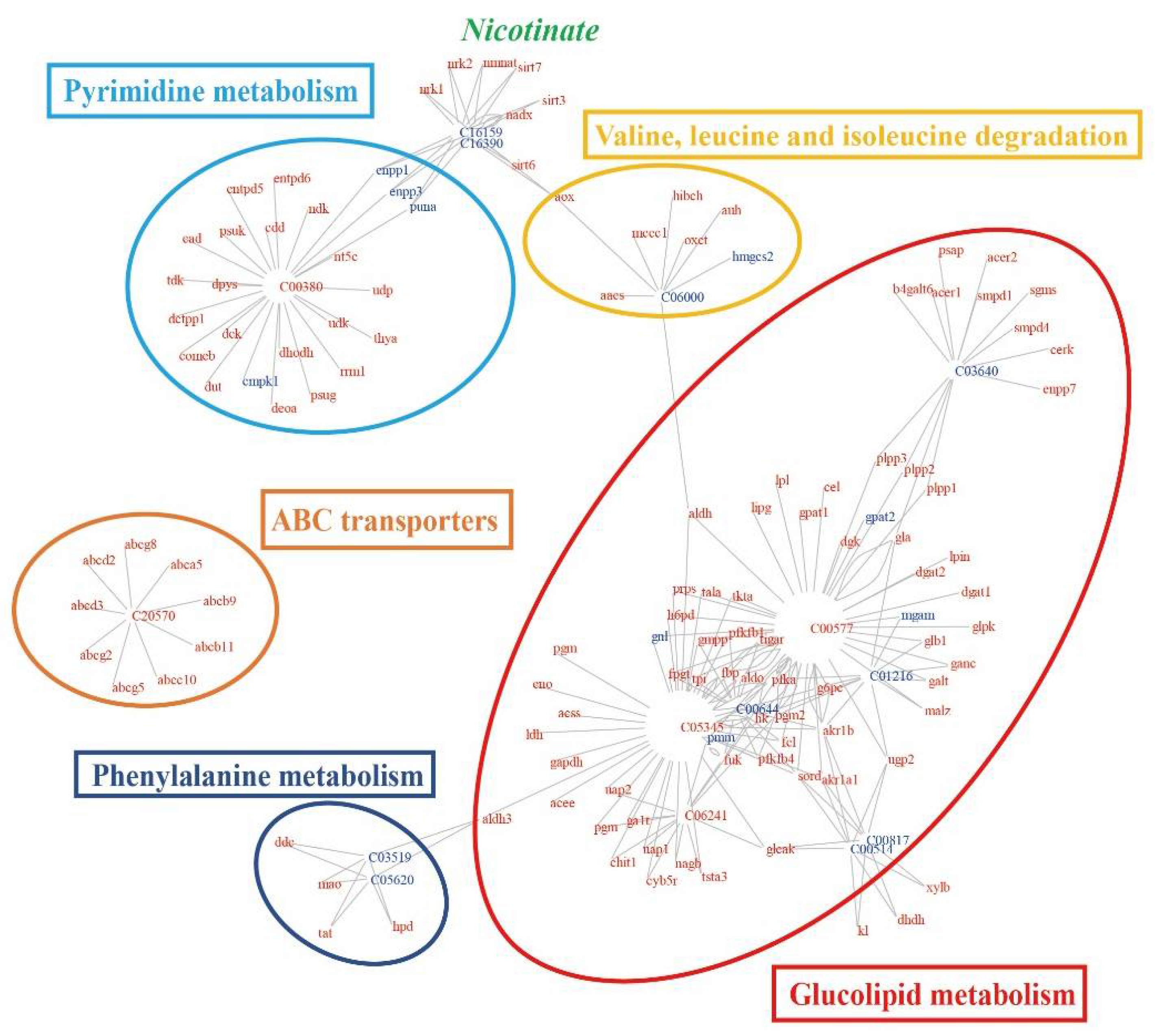
2.6. Pathway-Based Integration of Metabolomic and Transcriptomic Datasets
3. Discussion
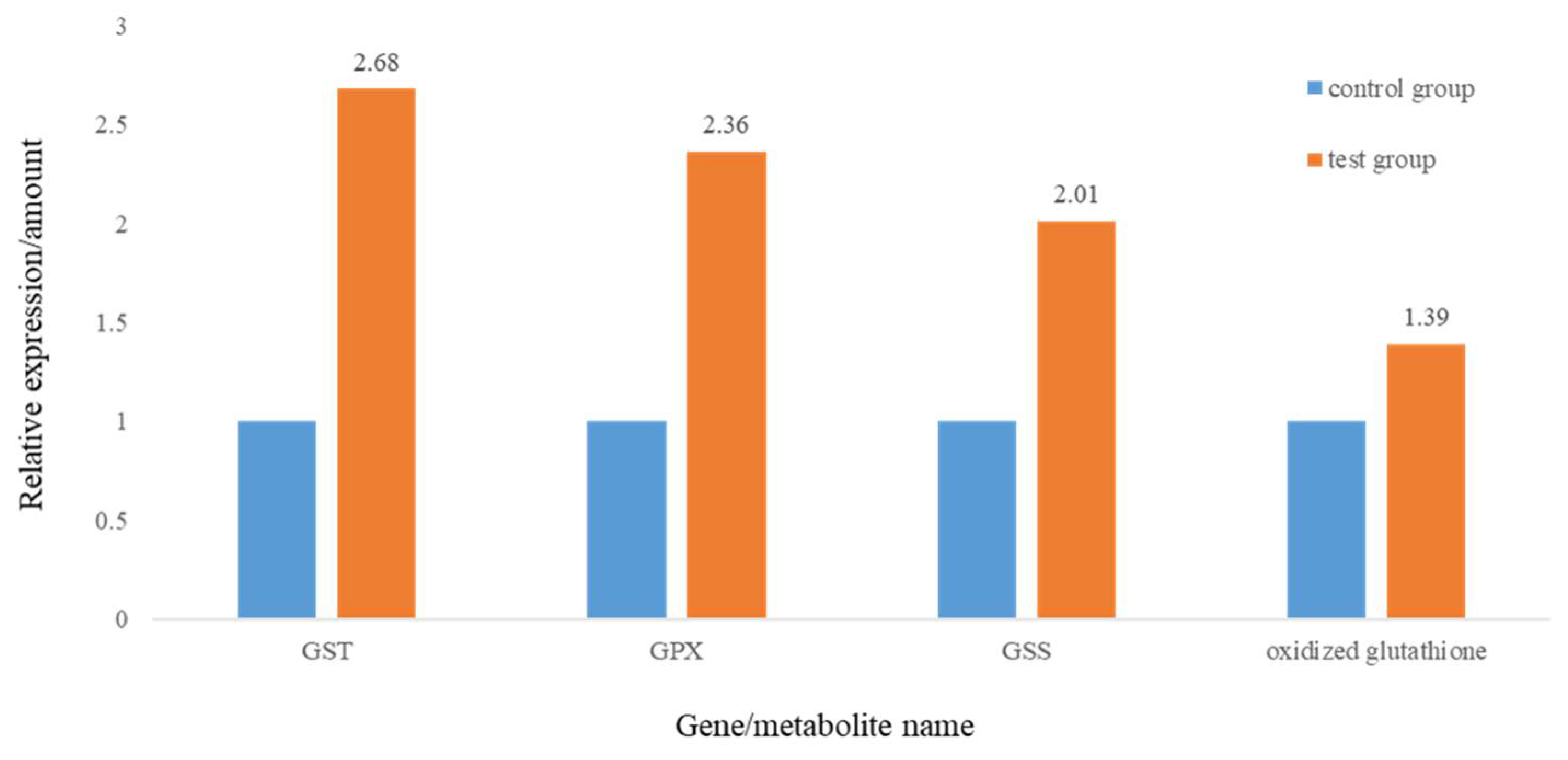
3.1. MP Exposure Induced Immune Reaction and Oxidative Stress in Rare Minnow
3.2. MP Exposure Disturbed Glycolipid and Energy Metabolism in Rare Minnow
4. Materials and Methods
4.1. Fish Maintenance
4.2. MPs Exposure
4.3. Histopathological Analysis
4.4. RNA Extraction, Library Preparation, and Transcriptome Sequencing
4.5. Metabolite Analysis
4.6. Metabolomics and Transcriptomics Combination Analysis
4.7. Validation of DEGs by qRT-PCR
4.8. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, C.; Wang, J.; Zhou, A.; Ye, Q.; Feng, Y.; Wang, Z.; Wang, S.; Xu, G.; Zou, J. Species-specific effect of microplastics on fish embryos and observation of toxicity kinetics in larvae. J. Hazard. Mater. 2021, 403, 123948. [Google Scholar] [CrossRef] [PubMed]
- Jenna, R.; Jambeck, R.G.; Chris, W.; Theodore, R.; Siegler, M.P.; Anthony, A.; Ramani, N.; Kara, L.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar]
- Frias, J.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.W.; Beckingham, B.A.; Ingram, B.C.; Ballenger, J.C.; Weinstein, J.E.; Sancho, G. Microplastic and tire wear particle occurrence in fishes from an urban estuary: Influence of feeding characteristics on exposure risk. Mar. Pollut. Bull. 2020, 160, 111539. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, K.; Su, L.; Li, J.; Yang, D.; Tong, C.; Mu, J.; Shi, H. Microplastics and mesoplastics in fish from coastal and fresh waters of China. Environ. Pollut. 2017, 221, 141–149. [Google Scholar] [CrossRef]
- McNeish, R.E.; Kim, L.H.; Barrett, H.A.; Mason, S.A.; Kelly, J.J.; Hoellein, T.J. Microplastic in riverine fish is connected to species traits. Sci. Rep. 2018, 8, 11639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assas, M.; Qiu, X.; Chen, K.; Ogawa, H.; Xu, H.; Shimasaki, Y.; Oshima, Y. Bioaccumulation and reproductive effects of fluorescent microplastics in medaka fish. Mar. Pollut. Bull. 2020, 158, 111446. [Google Scholar] [CrossRef]
- Wang, W.; Ge, J.; Yu, X. Bioavailability and toxicity of microplastics to fish species: A review. Ecotoxicol. Environ. Saf. 2020, 189, 109913. [Google Scholar] [CrossRef]
- Pang, M.; Wang, Y.; Tang, Y.; Dai, J.; Tong, J.; Jin, G. Transcriptome sequencing and metabolite analysis reveal the toxic effects of nanoplastics on tilapia after exposure to polystyrene. Environ. Pollut. 2021, 277, 116860. [Google Scholar] [CrossRef]
- Browne, M.A.; Dissanayake, A.; Gaiioway, T.S.; Lowe, D.M.; Thompson, R.C. Ingested microscopic plastic translocates to the circulatory system of the mussel, Mytilus edulis (L.). Environ. Sci. Technol. 2008, 42, 5026–5031. [Google Scholar] [CrossRef]
- Devriese, L.I.; van der Meulen, M.D.; Maes, T.; Bekaert, K.; Paul-Pont, I.; Frere, L.; Robbens, J.; Vethaak, A.D. Microplastic contamination in brown shrimp (Crangon crangon, Linnaeus 1758) from coastal waters of the Southern North Sea and channel area. Mar. Pollut. Bull. 2015, 98, 179–187. [Google Scholar] [CrossRef]
- Watts, A.J.; Urbina, M.A.; Corr, S.; Lewis, C.; Galloway, T.S. Ingestion of plastic Microfibers by the crab Carcinus maenas and its effect on food consumption and energy balance. Environ. Sci. Technol. 2015, 49, 14597–14604. [Google Scholar] [CrossRef]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The physical impacts of microplastics on marine organisms: A review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.; Le Goic, N.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S.L.; Kelly, F.J. Plastic and Human Health A Micro Issue? Environ. Sci. Technol. 2017, 51, 6634–6647. [Google Scholar] [CrossRef]
- Ian, A.K.; Michael, A.C.; Elda, M.; Roy, W.; James, J.R.; Pierre, G.; Florian, P. Seafloor microplastic hotspots controlled by deep-sea circulation. Science 2020, 368, 1140–1145. [Google Scholar]
- Yang, H.; Chen, G.; Wang, J. Microplastics in the Marine Environment: Sources, Fates, Impacts and Microbial Degradation. Toxics 2021, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Barboza, L.G.A.; Vieira, L.R.; Branco, V.; Figueiredo, N.; Carvalho, F.; Carvalho, C.; Guilhermino, L. Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchus labrax (Linnaeus, 1758). Aquat. Toxicol. 2018, 195, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhou, W.; Tang, Y.; Shi, W.; Shao, Y.; Ren, P.; Zhang, J.; Xiao, G.; Sun, H.; Liu, G. Microplastics aggravate the bioaccumulation of three veterinary antibiotics in the thick shell mussel Mytilus coruscus and induce synergistic immunotoxic effects. Sci. Total Environ. 2021, 770, 145273. [Google Scholar] [CrossRef]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding Type and Development Drive the Ingestion of Microplastics by Freshwater Invertebrates. Sci. Rep. 2017, 7, 17006. [Google Scholar] [CrossRef] [Green Version]
- Walkinshaw, C.; Lindeque, P.K.; Thompson, R.; Tolhurst, T.; Cole, M. Microplastics and Seafood: Lower Trophic Organisms at Highest Risk of Contamination. Ecotoxicol. Environ. Saf. 2020, 190, 110066. [Google Scholar] [CrossRef]
- Albano, M.; Panarello, G.; Di Paola, D.; Capparucci, F.; Crupi, R.; Gugliandolo, E.; Spanò, N.; Capillo, G.; Savoca, S. The influence of polystyrene microspheres abundance on development and feeding behavior of Artemia salina (Linnaeus, 1758). Appl. Sci. 2021, 11, 3352. [Google Scholar] [CrossRef]
- Plee, T.A.; Pomory, C.M. Microplastics in sandy environments in the Florida Keys and the panhandle of Florida, and the ingestion by sea cucumbers (Echinodermata: Holothuroidea) and sand dollars (Echinodermata: Echinoidea). Mar. Pollut. Bull. 2020, 158, 111437. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Qiu, X.C.; Xu, X.N.; Takai, Y.K.; Ogawa, H.; Shimasaki, Y. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Scopetani, C.; Esterhuizen, M.; Cincinelli, A.; Pflugmacher, S. Microplastics Exposure Causes Negligible Effects on the Oxidative Response Enzymes Glutathione Reductase and Peroxidase in the Oligochaete Tubifex tubifex. Toxics 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qiu, X.; Xu, X.; Takai, Y.; Ogawa, H.; Shimasaki, Y.; Oshima, Y. Uptake and depuration kinetics of microplastics with different polymer types and particle sizes in Japanese medaka (Oryzias latipes). Ecotoxicol. Environ. Saf. 2021, 212, 112007. [Google Scholar] [CrossRef]
- Espinosa, C.; Esteban, M.A.; Cuesta, A. Dietary administration of PVC and PE microplastics produces histological damage, oxidative stress and immunoregulation in European sea bass (Dicentrarchus labrax L.). Fish Shellfish Immunol. 2019, 95, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.X.; Wang, N.H.; Hu, X.D.; Yin, D.C.; Wu, C.X.; Liang, H.F.; Cao, W.X.; Cao, H. Acute toxic effects of lead (Pb2+) exposure to rare minnow (Gobiocypris rarus) revealed by histopathological examination and transcriptome analysis. Environ. Toxicol. Pharmacol. 2020, 78, 103385. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, S.; Zhang, Y.; Yang, Y.; Yuan, C.; Chen, S.; Wang, Z. Effects of 17 alpha-methyltestosterone on transcriptome, gonadal histology and sex steroid hormones in rare minnow (Gobiocypris rarus). Comp. Biochem. Physiol. D 2015, 15, 20–27. [Google Scholar]
- Wang, W.; Ndungu, A.W.; Li, Z.; Wang, J. Microplastics pollution in inland freshwaters of China: A case study in urban surface waters of Wuhan, China. Sci. Total Environ. 2017, 575, 1369–1374. [Google Scholar] [CrossRef]
- Nigamatzyanova, L.; Fakhrullin, R. Dark-field hyperspectral microscopy for label-free microplastics and nanoplastics detection and identification in vivo: A Caenorhabditis elegans study. Environ. Pollut. 2021, 271, 116337. [Google Scholar]
- Della Torre, C.; Bergami, E.; Salvati, A.; Faleri, C.; Cirino, P.; Dawson, K.A.; Cors, I. Accumulation and Embryotoxicity of Polystyrene Nanoparticles at Early Stage of Development of Sea Urchin Embryos Paracentrotus lividus. Environ. Sci. Technol. 2014, 48, 12302–12311. [Google Scholar]
- Gambardella, C.; Morgana, S.; Bramini, M.; Rotini, A.; Manfra, L.; Migliore, L.; Piazza, V.; Garaventa, F.; Faimali, M. Ecotoxicological effects of polystyrene microbeads in a battery of marine organisms belonging to different trophic levels. Mar. Environ. Res. 2018, 141, 313–321. [Google Scholar]
- Ortiz-Villanueva, E.; Navarro-Martin, L.; Jaumot, J.; Benavente, F.; Sanz-Nebot, V.; Pina, B.; Tauler, R. Metabolic disruption of zebrafish (Danio rerio) embryos by bisphenol A. An integrated metabolomic and transcriptomic approach. Environ. Pollut. 2017, 231, 22–36. [Google Scholar] [CrossRef]
- Holland, M.C.; Lambris, J.D. The complement system in teleosts. Fish Shellfish Immunol. 2002, 12, 399–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Moos, N.; Burkhardt-Holm, P.; Kohler, A. Uptake and effects of microplastics on cells and tissue of the blue mussel Mytilus edulis L. after an experimental exposure. Environ. Sci. Technol. 2012, 46, 11327–11335. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yu, Y.B.; Choi, J.H. Toxic effects on bioaccumulation, hematological parameters, oxidative stress, immune responses and neurotoxicity in fish exposed to microplastics: A review. J. Hazard. Mater. 2021, 413, 125423. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, P.; Katsumiti, A.; Nieto, J.A.; Bori, J.; Jimeno-Romero, A.; Reip, P.; Arostegui, I.; Orbea, A.; Cajaraville, M.P. Short-term effects on antioxidant enzymes and long-term genotoxic and carcinogenic potential of CuO nanoparticles compared to bulk CuO and ionic copper in mussels Mytilus galloprovincialis. Mar. Environ. Res. 2015, 111, 107–120. [Google Scholar] [CrossRef]
- Shi, W.; Han, Y.; Sun, S.; Tang, Y.; Zhou, W.; Du, X.; Liu, G. Immunotoxicities of microplastics and sertraline, alone and in combination, to a bivalve species: Size-dependent interaction and potential toxication mechanism. J. Hazard. Mater. 2020, 396, 122603. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhou, W.; Sun, S.; Du, X.; Han, Y.; Shi, W.; Liu, G. Immunotoxicity and neurotoxicity of bisphenol A and microplastics alone or in combination to a bivalve species, Tegillarca granosa. Environ. Pollut. 2020, 265, 115115. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Aspects Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.G.; Hou, X.; Chen, X.W.; Zhang, K.J.; Wang, J.; Wang, C.H. Comprehensive analysis of metabolomics and transcriptomics provides insights into growth difference of juvenile Eriocheir sinensis during the molting cycle. Aquaculture 2021, 539, 736661. [Google Scholar] [CrossRef]
- Jeong, C.B.; Kang, H.M.; Lee, M.C.; Kim, D.H.; Han, J.; Hwang, D.S.; Souissi, S.; Lee, S.J.; Shin, K.H.; Park, H.G.; et al. Adverse effects of microplastics and oxidative stress-induced MAPK/Nrf2 pathway-mediated defense mechanisms in the marine copepod Paracyclopina nana. Sci. Rep. 2017, 7, 41323. [Google Scholar] [CrossRef] [Green Version]
- Wood, T. Physiological Functions of the Pentose Phosphate Pathway. Cell Biochem. Funct. 1986, 4, 241–247. [Google Scholar] [CrossRef]
- Riganti, C.; Gazzano, E.; Polimeni, M.; Aldieri, E.; Ghigo, D. The pentose phosphate pathway: An antioxidant defense and a crossroad in tumor cell fate. Free Radic. Biol. Med. 2012, 53, 421–436. [Google Scholar] [CrossRef]
- Deng, Y.F.; Zhang, Y.; Lemos, B.; Ren, H.Q. Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Wang, C.; Zhou, J.; Shen, M.; Wang, X.; Fu, Z.; Jin, Y. Effects of polystyrene microplastics on the composition of the microbiome and metabolism in larval zebrafish. Chemosphere 2019, 217, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.; Zhao, J.M.; Zhu, X.P.; Shan, E.; Wang, Q. Oxidative stress biomarkers, physiological responses and proteomic profiling in oyster (Crassostrea gigas) exposed to microplastics with irregular-shaped PE and PET microplastic. Sci. Total Environ. 2021, 786, 147425. [Google Scholar] [CrossRef]
- De Bose-Boyd, R.A. Significance and regulation of lipid metabolism. Semin. Cell Dev. Biol. 2018, 81, 97. [Google Scholar] [CrossRef]
- Laudicella, V.A.; Whitfield, P.D.; Carboni, S.; Doherty, M.K.; Hughes, A.D. Application of lipidomics in bivalve aquaculture, a review. Rev. Aquac. 2019, 12, 678–702. [Google Scholar] [CrossRef]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.L.; Zhang, F.T.; Cao, W.X.; Wang, J.W. The identification of apolipoprotein C-I in rare minnow (Gobiocypris rarus) and its expression following cadmium exposure. Environ. Toxicol. Pharmacol. 2013, 35, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Magni, S.; Gagne, F.; Andre, C.; Della Torre, C.; Auclair, J.; Hanana, H.; Parenti, C.C.; Bonasoro, F.; Binelli, A. Evaluation of uptake and chronic toxicity of virgin polystyrene microbeads in freshwater zebra mussel Dreissena polymorpha (Mollusca: Bivalvia). Sci. Total Environ. 2018, 631–632, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Wu, S.; Lu, S.; Liu, M.; Song, Y.; Fu, Z.; Shi, H.; Raley-Susman, K.M.; He, D. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans. Sci. Total Environ. 2018, 619–620, 1–8. [Google Scholar] [CrossRef]
- Leng, X.Q.; Zhou, H.; Tan, Q.S.; Du, H.; Wu, J.P.; Liang, X.F.; He, S.; Wei, Q.W. Integrated metabolomic and transcriptomic analyses suggest that high dietary lipid levels facilitate ovary development through the enhanced arachidonic acid metabolism, cholesterol biosynthesis and steroid hormone synthesis in Chinese sturgeon (Acipenser sinensis). Br. J. Nutr. 2019, 122, 1230–1241. [Google Scholar]
- Qiao, R.; Sheng, C.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics induce intestinal inflammation, oxidative stress, and disorders of metabolome and microbiome in zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cavill, R.; Jennen, D.; Kleinjans, J.; Briede, J.J. Transcriptomic and metabolomic data integration. Brief. Bioinform. 2016, 17, 891–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Before/After Refreshing Water | Dissolved Oxygen (mg/L) | Conductivity (μs/cm) | pH | Temperature (°C) |
|---|---|---|---|---|---|
| Control | before | 7.55 ± 0.36 | 747 ± 7 | 8.03 ± 0.17 | 26.8 ± 1.1 |
| after | 7.77 ± 0.15 | 732 ± 3 | 8.29 ± 0.06 | 27.3 ± 0.9 | |
| MPs-treated | before | 7.57 ± 0.46 | 750 ± 9 | 8.04 ± 0.22 | 26.6 ± 1.2 |
| after | 7.71 ± 0.20 | 734 ± 2 | 8.25 ± 0.07 | 27.1 ± 1.0 |
| Gene Name | Description | Log2fc | Q-Value | Biological Process | Function |
|---|---|---|---|---|---|
| hutU | urocanate hydratase | −12.08 | 5.27 × 10−254 | Histidine metabolism | Catalyzes the conversion of urocanic acid |
| C3 | Complement component 3 | −11.71 | 2.10 × 10−210 | Immune process | Part of the complement system |
| UGT | glucuronosyltransferase | −11.43 | 1.25 × 10−181 | Glucuronidation process | Making relative enzymes |
| C1R | Complement component 1, r subcomponent | −11.24 | 1.66 × 10−164 | Immune process | Part of the complement system |
| MHC1 | major histocompatibility complex | −10.96 | 3.80 × 10−142 | Immune process | Making proteins in certain immune system cells |
| FKBP4 | FK506-binding protein 4 | −9.89 | 2.23 × 10−80 | Immune process | Signal transduction |
| HSD11B2 | corticosteroid 11-beta-dehydrogenase isozyme 2 | −9.57 | 8.38 × 10−68 | Immune process | Modulates intracellular glucocorticoid levels |
| FMN2 | formin 2 | −8.90 | 5.06 × 10−47 | Organismal Systems | Development |
| RAP1GAP | RAP1 GTPase activating protein 1 | 9.60 | 8.43 × 10−68 | Cancer tumorigenesis | GTPase activator |
| NRAS | GTPase NRas | 9.67 | 1.35 × 10−70 | Cell division | Signal transduction |
| RELA | transcription factor p65 | 9.69 | 2.28 × 10−71 | Inflammatory reaction | Signal transduction |
| C4 | Complement component 4 | 10.52 | 6.84 × 10−111 | Immune process | Part of the complement system |
| TACC3 | transforming acidic coiled-coil-containing protein 3 | 10.59 | 2.30 × 10−115 | Cell growth and differentiation | Signal transduction |
| CAMK1 | calcium/calmodulin- dependent protein kinase I | 10.61 | 1.48 × 10−116 | Cell growth and death | Signal transduction |
| TF | transferrin | 10.65 | 2.84 × 10−119 | Environmental Information Process | Signal transduction |
| A2M | alpha-2-macroglobulin | 10.79 | 3.15 × 10−128 | Immune process | Proteinase inhibitor |
| PLAUR | plasminogen activator, urokinase receptor | 9.35 | 3.49 × 10−59 | Immune process | Signal transduction |
| RNF19A | E3 ubiquitin-protein ligase RNF19A | 9.25 | 3.81 × 10−56 | Genetic Information Process | Translation |
| SHOC2 | leucine-rich repeat protein SHOC2 | 9.04 | 6.74 × 10−50 | Environmental Information Process | Signal transduction |
| ALDH | aldehyde dehydrogenase | 8.74 | 2.74 × 10−42 | Metabolism process | Producing aldehyde enzymes |
| Compound | Molecular Formula | Mode | Measured Molecular Mass (Da) | Mass Error (ppm) | Fold-Change | Trend | p-Value | Kegg C-Code |
|---|---|---|---|---|---|---|---|---|
| 2-Formylglutarate | C6H8O5 | ESI- | 159.0293 | −3.91076 | 0.64 | down | 0.002 | C16159 |
| 2-Dehydro-3-deoxy-d-galactonate | C6H10O6 | ESI- | 177.04 | −2.53718 | 0.63 | down | 0.001 | C01216 |
| d-Galactono-1,5-lactone | C6H10O6 | ESI- | 177.04 | −2.53718 | 0.63 | down | 0.001 | C02669 |
| l-Galactono-1,5-lactone | C6H10O6 | ESI- | 177.04 | −2.53718 | 0.63 | down | 0.001 | C21955 |
| d-Mannitol 1-phosphate | C6H15O9P | ESI- | 307.043 | −2.00474 | 0.67 | down | 0.023 | C00644 |
| d-Altronate | C6H12O7 | ESI- | 195.0505 | −2.85294 | 0.61 | down | 0.001 | C00817 |
| l-Lyxose | C5H10O5 | ESI- | 195.0505 | −3.72763 | 0.61 | down | 0.001 | C01508 |
| d-Ribose | C5H10O5 | ESI- | 195.0505 | −3.72763 | 0.61 | down | 0.001 | C00121 |
| d-Lyxose | C5H10O5 | ESI- | 195.0505 | −3.72763 | 0.61 | down | 0.001 | C00476 |
| d-Mannonate | C6H12O7 | ESI- | 195.0505 | −2.85294 | 0.61 | down | 0.001 | C00514 |
| (S)-3-Hydroxyisobutyryl-CoA | C25H42N7O18P3S | ESI- | 852.1423 | −2.79862 | 0.59 | down | 0.044 | C06000 |
| Sphingosyl-phosphocholine | C23H50N2O5P+ | ESI- | 510.3457 | 3.888599 | 0.64 | down | 0.040 | C03640 |
| N-Acetyl-d-phenylalanine | C11H13NO3 | ESI- | 252.087 | −3.40067 | 0.68 | down | 0.048 | C05620 |
| N-Acetyl-l-phenylalanine | C11H13NO3 | ESI- | 252.087 | −3.40067 | 0.68 | down | 0.048 | C03519 |
| d-Mannonate | C6H12O7 | ESI+ | 219.0459 | −8.34259 | 0.67 | down | 0.010 | C00514 |
| d-Altronate | C6H12O7 | ESI+ | 219.0459 | −8.34259 | 0.67 | down | 0.010 | C00817 |
| 5-Hydroxyindoleacetate | C10H9NO3 | ESI+ | 192.0659 | 1.905397 | 0.61 | down | 0.044 | C05635 |
| (R)-3,3-Dimethylmalate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C01088 |
| l-Fucono-1,5-lactone | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C18028 |
| (S)-2-(Hydroxymethyl)glutarate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C16390 |
| 2-Dehydro-3-deoxy-l-fuconate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C03827 |
| 3-Ethylmalate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C01989 |
| (2R,3S)-2,3-Dimethylmalate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C03652 |
| (R)-2-Ethylmalate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C02488 |
| 2-Dehydro-3-deoxy-l-rhamnonate | C6H10O5 | ESI+ | 180.0877 | 6.414412 | 0.66 | down | 0.016 | C03979 |
| 4-Methyl-l-glutamate | C6H11NO4 | ESI+ | 144.0655 | -0.08917 | 0.48 | down | 0.006 | C06234 |
| alpha-1,5-l-Arabinobiose | C10H18O9 | ESI- | 281.0882 | 1.31699 | 0.78 | down | 0.004 | C20570 |
| beta-d-Glucose 1-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C00663 |
| beta-d-Fructose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C05345 |
| alpha-d-Glucose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C00668 |
| d-Mannose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C00275 |
| d-Tagatose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C01097 |
| d-Fructose 1-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C01094 |
| d-Allulose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C18096 |
| l-Gulose 1-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C15924 |
| beta-d-Glucose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C01172 |
| d-Allose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C02962 |
| N-Acetylneuraminate 9-phosphate | C11H20NO12P | ESI+ | 428.037 | 3.966776 | 1.57 | up | 0.000 | C06241 |
| Cytosine | C4H5N3O | ESI+ | 112.0506 | 0.697346 | 1.76 | up | 0.002 | C00380 |
| d-Glyceraldehyde | C3H6O3 | ESI- | 135.0307 | 9.084401 | 1.59 | up | 0.046 | C00577 |
| d-Fructose 6-phosphate | C6H13O9P | ESI+ | 283.019 | 0.420669 | 1.28 | up | 0.046 | C00085 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Hou, M.; Shang, K.; Wang, H.; Wang, J. Microplastics (Polystyrene) Exposure Induces Metabolic Changes in the Liver of Rare Minnow (Gobiocypris rarus). Molecules 2022, 27, 584. https://doi.org/10.3390/molecules27030584
Wang C, Hou M, Shang K, Wang H, Wang J. Microplastics (Polystyrene) Exposure Induces Metabolic Changes in the Liver of Rare Minnow (Gobiocypris rarus). Molecules. 2022; 27(3):584. https://doi.org/10.3390/molecules27030584
Chicago/Turabian StyleWang, Chunling, Miaomiao Hou, Kunyu Shang, Huanshan Wang, and Jianwei Wang. 2022. "Microplastics (Polystyrene) Exposure Induces Metabolic Changes in the Liver of Rare Minnow (Gobiocypris rarus)" Molecules 27, no. 3: 584. https://doi.org/10.3390/molecules27030584
APA StyleWang, C., Hou, M., Shang, K., Wang, H., & Wang, J. (2022). Microplastics (Polystyrene) Exposure Induces Metabolic Changes in the Liver of Rare Minnow (Gobiocypris rarus). Molecules, 27(3), 584. https://doi.org/10.3390/molecules27030584