
Protic Ionic Liquid Cation Alkyl Chain Length Effect on Lysozyme Structure

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Materials
2.2. Sample Preparation
2.3. Analytical Characterisation
3. Results and Discussion
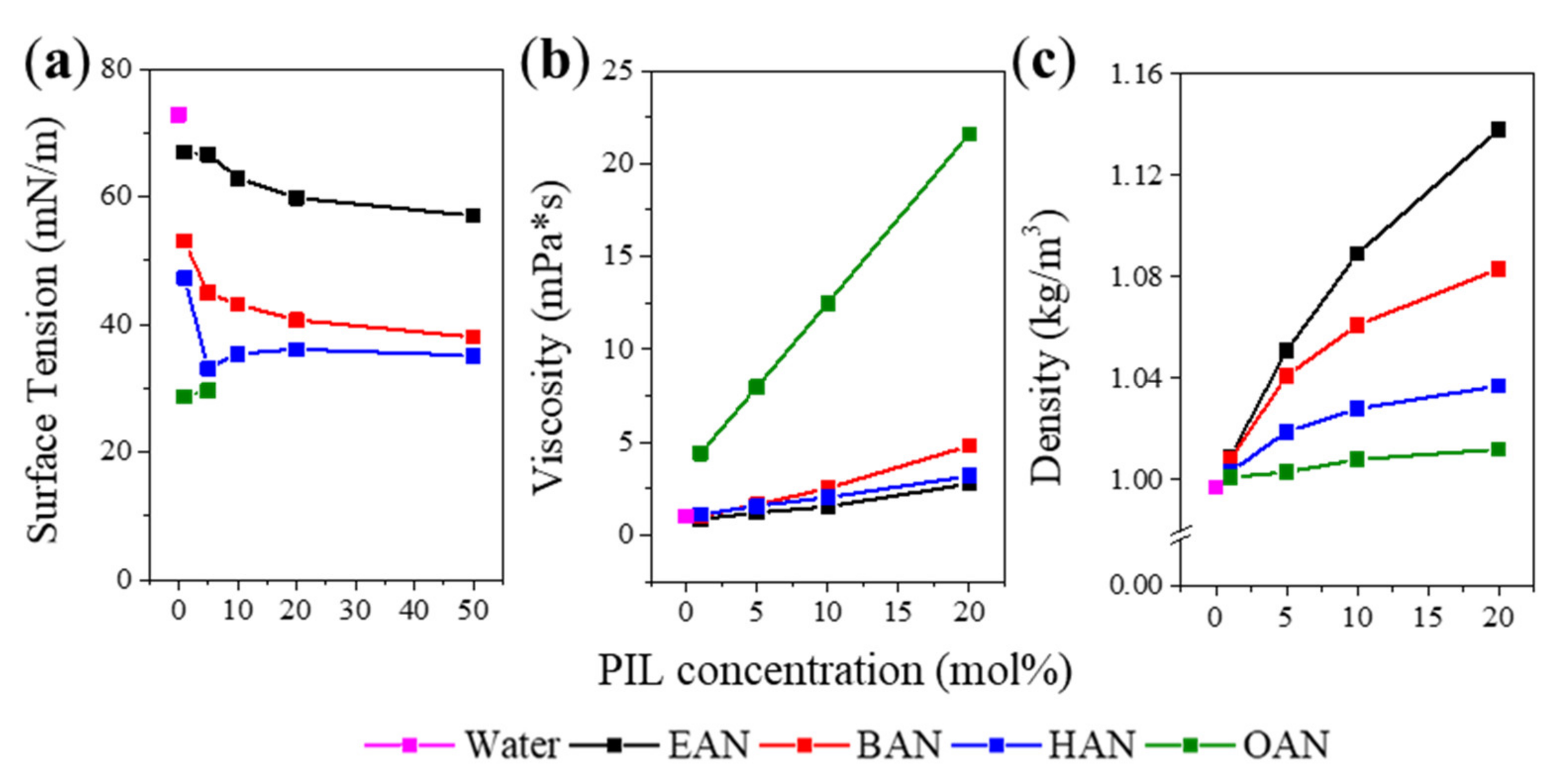
3.1. Physicochemical Properties of PIL-Water Mixture
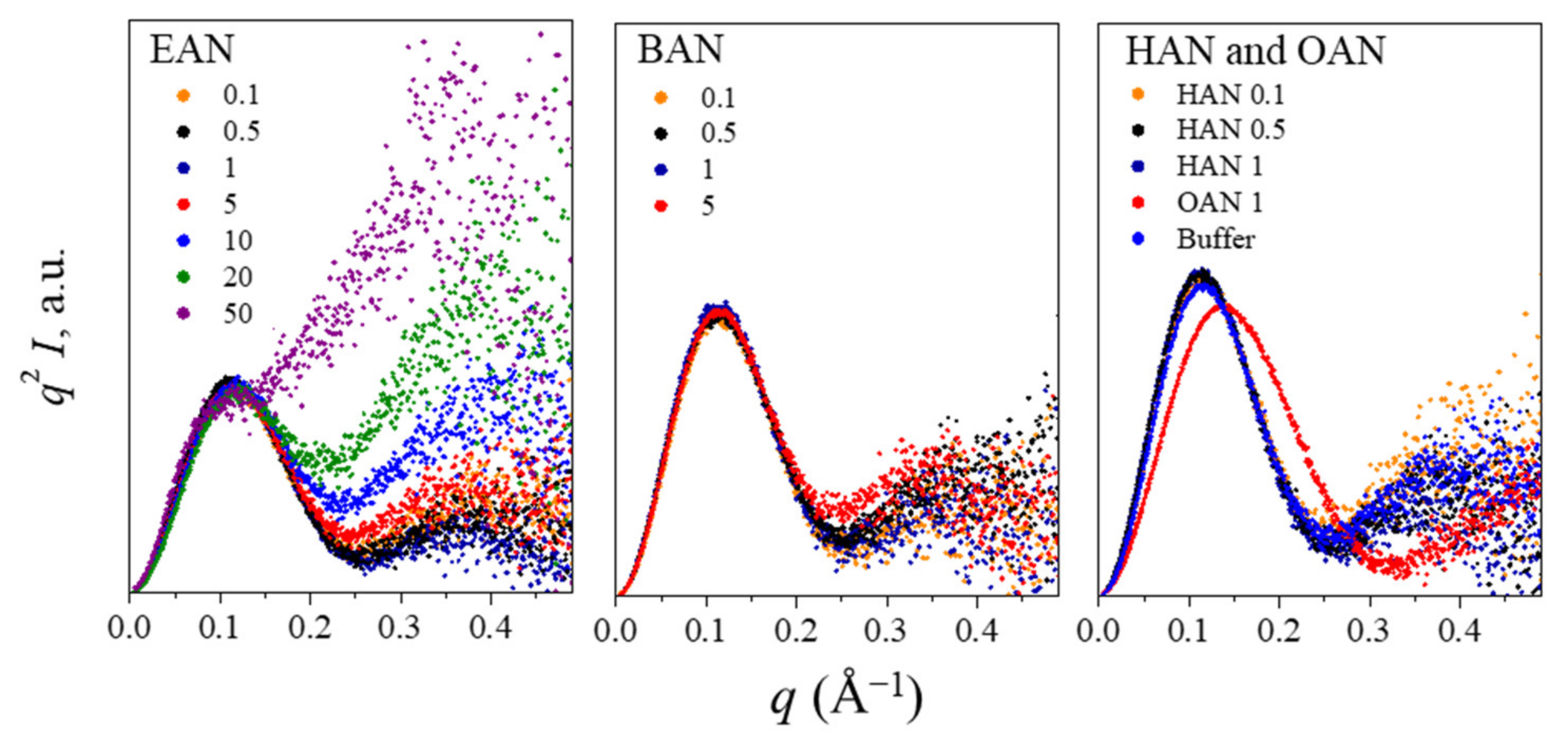
3.2. Liquid Nanostructure
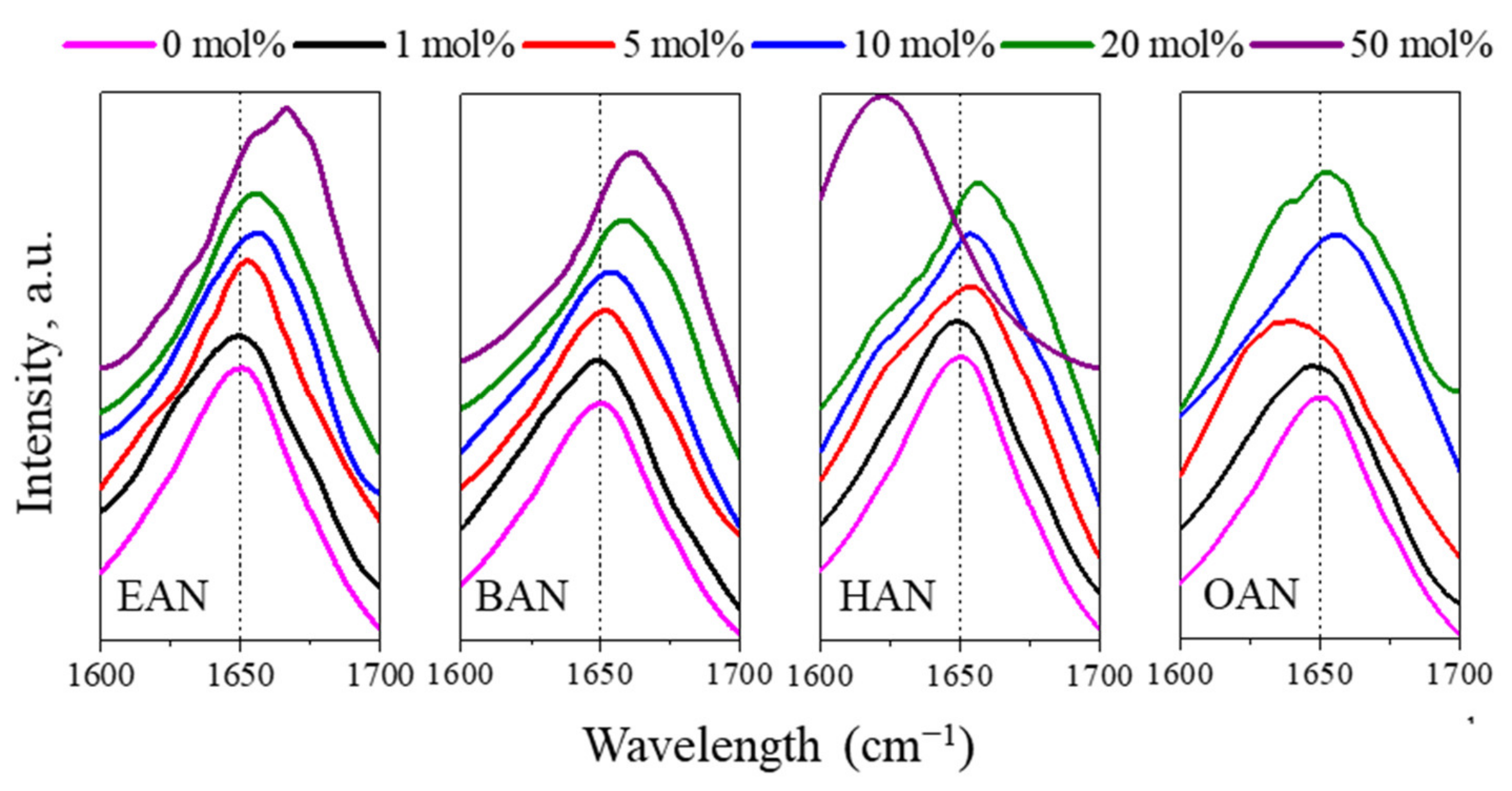
3.3. Conformational Changes of Lysozyme
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Balcão, V.M.; Vila, M.M.D.C. Structural and functional stabilization of protein entities: State-of-the-art. Adv. Drug Deliv. Rev. 2015, 93, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Scharnagl, C.; Reif, M.; Friedrich, J. Stability of proteins: Temperature, pressure and the role of the solvent. Biochim. Biophys. Acta Proteins Proteom. BBA-PROTEINS PROTEOM. 2005, 1749, 187–213. [Google Scholar] [CrossRef] [PubMed]
- Chi, E.Y.; Krishnan, S.; Randolph, T.W.; Carpenter, J.F. Physical stability of proteins in aqueous solution: Mechanism and driving forces in nonnative protein aggregation. Pharm. Res. 2003, 20, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. Advanced protein formulations. Protein Sci. 2015, 24, 1031–1039. [Google Scholar] [CrossRef]
- Jessop, P.G. Searching for green solvents. Green Chem. Lett. Rev. 2011, 13, 1391–1398. [Google Scholar] [CrossRef]
- MacFarlane, D.R.; Chong, A.L.; Forsyth, M.; Kar, M.; Vijayaraghavan, R.; Somers, A.; Pringle, J.M. New dimensions in salt–solvent mixtures: A 4th evolution of ionic liquids. Faraday Discuss. 2018, 206, 9–28. [Google Scholar] [CrossRef]
- E Silva, F.A.; Pereira, J.F.B.; Kurnia, K.A.; Ventura, S.P.M.; Silva, A.M.S.; Rogers, R.D.; Coutinho, J.A.P.; Freire, M.G. Temperature dependency of aqueous biphasic systems: An alternative approach for exploring the differences between Coulombic-dominated salts and ionic liquids. Chem. Commun. 2017, 53, 7298–7301. [Google Scholar] [CrossRef]
- Welton, T. Room-temperature ionic liquids. Solvents for synthesis and catalysis. Chem. Rev. 1999, 99, 2071–2083. [Google Scholar] [CrossRef]
- Greaves, T.L.; Drummond, C.J. Protic ionic liquids: Properties and applications. Chem. Rev. 2008, 108, 206–237. [Google Scholar] [CrossRef]
- MacFarlane, D.R.; Kar, M.; Pringle, J.M. Fundamentals of Ionic Liquids: From Chemistry to Applications; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Yoshizawa, M.; Xu, W.; Angell, C.A. Ionic Liquids by Proton Transfer: Vapor Pressure, Conductivity, and the Relevance of ΔpKa from Aqueous Solutions. J. Am. Chem. Soc. 2003, 125, 15411–15419. [Google Scholar] [CrossRef]
- Greaves, T.L.; Drummond, C.J. Protic Ionic Liquids: Evolving Structure-Property Relationships and Expanding Applications. Chem. Rev. 2015, 115, 11379–11448. [Google Scholar] [CrossRef] [PubMed]
- Schröder, C. Proteins in ionic liquids: Current status of experiments and simulations. In Ionic Liquids II; Kirchner, B., Perlt, E., Eds.; Springer: Cham, Switzerland, 2017; Volume 375, pp. 127–152. [Google Scholar]
- Schindl, A.; Hagen, M.L.; Muzammal, S.; Gunasekera, H.A.D.; Croft, A.K. Proteins in ionic liquids: Reactions, applications, and futures. Front. Chem. 2019, 7, 347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H. Protein stabilization and enzyme activation in ionic liquids: Specific ion effects. J. Chem. Technol. Biot. 2016, 91, 25–50. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Brown, S.; Drummond, C.J.; Greaves, T.L. Protein aggregation and crystallization with ionic liquids: Insights into the influence of solvent properties. J. Colloid Interf. Sci. 2022, 608, 1173–1190. [Google Scholar] [CrossRef]
- Kohno, Y.; Ohno, H. Ionic liquid/water mixtures: From hostility to conciliation. Chem. Commun. 2012, 48, 7119–7130. [Google Scholar] [CrossRef]
- Lai, J.Q.; Li, Z.; Lu, Y.H.; Yang, Z. Specific ion effects of ionic liquids on enzyme activity and stability. Green Chem. 2011, 13, 1860–1868. [Google Scholar] [CrossRef]
- Van Rantwijk, F.; Sheldon, R.A. Biocatalysis in ionic liquids. Chem. Rev. 2007, 107, 2757–2785. [Google Scholar] [CrossRef]
- Han, Q.; Wang, X.; Bynre, N. Utilizing water activity as a simple measure to understand hydrophobicity in ionic liquids. Front. Chem. 2019, 7, 112. [Google Scholar] [CrossRef]
- Takekiyo, T.; Ishikawa, Y.; Yoshimura, Y. Cryopreservation of proteins using ionic liquids: A case study of cytochrome c. J. Phys. Chem 2017, 121, 7614–7620. [Google Scholar] [CrossRef]
- Byrne, N.; Wang, L.M.; Belieres, J.P.; Angell, C.A. Reversible folding-unfolding, aggregation protection, and multi-year stabilization, in high concentration protein solutions, using ionic liquids. Chem. Commun. 2007, 26, 2714–2716. [Google Scholar] [CrossRef]
- Han, Q.; Ryan, T.M.; Rosado, C.J.; Drummond, C.J.; Greaves, T.L. Effect of ionic liquids on the fluorescence properties and aggregation of superfolder green fluorescence protein. J. Colloid Interf. Sci. 2021, 591, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Binns, J.; Zhai, J.; Guo, X.; Ryan, T.M.; Drummond, C.J.; Greaves, T.L. Insights on lysozyme aggregation in protic ionic liquid solvents by using small angle X-ray scattering and high throughput screening. J. Mol. Liq. 2022, 345, 117816. [Google Scholar] [CrossRef]
- Weingartner, H.; Cabrele, C.; Herrmann, C. How ionic liquids can help to stabilize native proteins. Phys. Chem. Chem. Phys. 2012, 14, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.F.; Drummond, C.J.; Peat, T.S.; Newman, J. Evaluating protic ionic liquids as protein crystallization additives. Cryst. Growth Des. 2011, 11, 1777–1785. [Google Scholar] [CrossRef]
- Han, Q.; Smith, K.M.; Darmanin, C.; Ryan, T.M.; Drummond, C.J.; Greaves, T.L. Lysozyme conformational changes with ionic liquids: Spectroscopic, small angle x-ray scattering and crystallographic study. J. Colloid Interf. Sci. 2021, 585, 433–443. [Google Scholar] [CrossRef]
- MacFarlane, D.R.; Vijayaraghavan, R.; Ha, H.N.; Izgorodin, A.; Weaver, K.D.; Elliott, G.D. Ionic liquid "buffers"-pH control in ionic liquid systems. Chem. Commun. 2010, 46, 7703–7705. [Google Scholar] [CrossRef]
- Taha, M.; Almeida, M.R.; Silva, F.A.E.; Domingues, P.; Ventura, S.P.M.; Coutinho, J.A.P.; Freire, M.G. Novel Biocompatible and Self-buffering Ionic Liquids for Biopharmaceutical Applications. Chem.-Eur. J. 2015, 21, 4781–4788. [Google Scholar] [CrossRef]
- Taha, M.; Quental, M.V.; Correia, I.; Freire, M.G.; Coutinho, J.A.P. Extraction and stability of bovine serum albumin (BSA) using cholinium-based Good’s buffers ionic liquids. Process. Biochem. 2015, 50, 1158–1166. [Google Scholar] [CrossRef]
- Wijaya, E.C.; Separovic, F.; Drummond, C.J.; Greaves, T.L. Stability and activity of lysozyme in stoichiometric and non-stoichiometric protic ionic liquid (PIL)-water systems. J. Chem. Phys. 2018, 148, 193838. [Google Scholar] [CrossRef]
- Han, Q.; Wang, X.; Byrne, N. Understanding the Influence of Key Ionic Liquid Properties on the Hydrolytic Activity of Thermomyces lanuginosus Lipase. ChemCatChem 2016, 8, 1551–1556. [Google Scholar] [CrossRef]
- Han, Q.; Wang, X.; Byrne, N. A Simple Approach to Achieve Self-Buffering Protic Ionic Liquid-Water Mixtures. ChemistrySelect 2017, 2, 4294–4299. [Google Scholar] [CrossRef]
- Arunkumar, R.; Drummond, C.J.; Greaves, T.L. FTIR Spectroscopic Study of the Secondary Structure of Globular Proteins in Aqueous Protic Ionic Liquids. Front. Chem. 2019, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.F.; Queirós, R.P.; Santos, M.D.; Fidalgo, L.G.; Delgadillo, I.; Saraiva, J.A. Effect of ionic liquids alkyl chain length on horseradish peroxidase thermal inactivation kinetics and activity recovery after inactivation. World J. Microbiol. Biotechnol. 2014, 30, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Jha, I.; Bisht, M.; Mogha, N.K.; Venkatesu, P. Effect of imidazolium-based ionic liquids on the structure and stability of stem bromelain: Concentration and alkyl chain length effect. J. Phys. Chem. B 2018, 122, 7522–7529. [Google Scholar] [CrossRef] [PubMed]
- Rakowska, P.W.; Kloskowski, A. Impact of the Alkyl Side Chains of Cations and Anions on the Activity and Renaturation of Lysozyme: A Systematic Study Performed Using Six Amino-Acid-Based Ionic Liquids. ChemistrySelect 2021, 6, 3089–3095. [Google Scholar] [CrossRef]
- Islam, M.M.; Barik, S.; Sarkar, M. Probing the interactions of 1-alkyl-3-methylimidazolium tetrafluoroborate (Alkyl= Octyl, Hexyl, Butyl, and Ethyl) ionic liquids with bovine serum albumin: An alkyl chain length-dependent study. J. Phys. Chem. B 2019, 123, 1512–1526. [Google Scholar] [CrossRef]
- Greaves, T.L.; Drummond, C.J. Solvent nanostructure, the solvophobic effect and amphiphile self-assembly in ionic liquids. Chem. Soc. Rev. 2013, 42, 1096–1120. [Google Scholar] [CrossRef]
- Greaves, T.L.; Kennedy, D.F.; Weerawardena, A.; Tse, N.M.; Kirby, N.; Drummond, C.J. Nanostructured protic ionic liquids retain nanoscale features in aqueous solution while precursor Brønsted acids and bases exhibit different behavior. J. Phys. Chem. B 2011, 115, 2055–2066. [Google Scholar] [CrossRef]
- Greaves, T.L.; Ha, K.; Muir, B.W.; Howard, S.C.; Weerawardena, A.; Kirby, N.; Drummond, C.J. Protic ionic liquids (PILs) nanostructure and physicochemical properties: Development of high-throughput methodology for PIL creation and property screens. Phys. Chem. Chem. Phys. 2015, 17, 2357–2365. [Google Scholar] [CrossRef]
- Greaves, T.L.; Kennedy, D.F.; Mudie, S.T.; Drummond, C.J. Diversity observed in the nanostructure of protic ionic liquids. J. Phys. Chem. B 2010, 114, 10022–10031. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Hammel, M.; Tainer, J.A.; Sali, A. Accurate SAXS profile computation and its assessment by contrast variation experiments. Biophys. J. 2013, 105, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Greaves, T.L.; Broomhall, H.; Weerawardena, A.; Osborne, D.A.; Canonge, B.A.; Drummond, C.J. How ionic species structure influences phase structure and transitions from protic ionic liquids to liquid crystals to crystals. Faraday Discuss. 2018, 206, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Drummond, C.J.; Grieser, F.; Healy, T.W. Acid–base equilibria in aqueous micellar solutions. Part 1.—‘Simple’ weak acids and bases. J. Chem. Soc. Faraday Trans. 1 Phys. Chem. Condens. Phases 1989, 85, 521–535. [Google Scholar]
- Ohno, H.; Fujita, K.; Kohno, Y. Is seven the minimum number of water molecules per ion pair for assured biological activity in ionic liquid-water mixtures? Phys. Chem. Chem. Phys. 2015, 17, 14454–14460. [Google Scholar] [CrossRef]
- Kirby, N.; Cowieson, N.; Hawley, A.M.; Mudie, S.T.; McGillivray, D.J.; Kusel, M.; Samardzic-Boban, V.; Ryan, T.M. Improved radiation dose efficiency in solution SAXS using a sheath flow sample environment. Acta Crystallogr. D 2016, 72, 1254–1266. [Google Scholar] [CrossRef]
- Jeffries, C.M.; Graewert, M.A.; Blanchet, C.E.; Langley, D.B.; Whitten, A.E.; Svergun, D.I. Preparing monodisperse macromolecular samples for successful biological small-angle X-ray and neutron-scattering experiments. Nat. Protoc. 2016, 11, 2122–2153. [Google Scholar] [CrossRef]
- Panjkovich, A.; Svergun, D.I. Deciphering conformational transitions of proteins by small angle X-ray scattering and normal mode analysis. Phys. Chem. Chem. Phys. 2016, 18, 5707–5719. [Google Scholar] [CrossRef]
- Lin, T.Y.; Timasheff, S.N. On the role of surface tension in the stabilization of globular proteins. Protein Sci. 1996, 5, 372–381. [Google Scholar] [CrossRef]
- Uribe, S.; Sampedro, J.G. Measuring Solution Viscosity and its Effect on Enzyme Activity. Biol. Proc. Online 2003, 5, 108–115. [Google Scholar] [CrossRef]
- Das, S.K.; Majhi, D.; Sahu, P.K.; Sarkar, M. Investigation of the influence of alkyl side chain length on the fluorescence response of C153 in a series of room temperature ionic liquids. RSC Adv. 2015, 5, 41585–41594. [Google Scholar] [CrossRef]
- Clough, M.T.; Crick, C.R.; Grasvik, J.; Hunt, P.A.; Niedermeyer, H.; Welton, T.; Whitaker, O.P. A physicochemical investigation of ionic liquid mixtures. Chem. Sci. 2015, 6, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Hayes, R.; Warr, G.G.; Atkin, R. Structure and nanostructure in ionic liquids. Chem. Rev. 2015, 115, 6357–6426. [Google Scholar] [CrossRef] [PubMed]
- Pádua, A.A.H.; Costa Gomes, M.F.; Canongia Lopes, J.N.A. Molecular solutes in ionic liquids: A structural perspective. Accounts. Chem. Res. 2007, 40, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Greaves, T.L.; Kennedy, D.F.; Kirby, N.; Drummond, C.J. Nanostructure changes in protic ionic liquids (PILs) through adding solutes and mixing PILs. Phys. Chem Chem Phys. 2011, 13, 13501–13509. [Google Scholar] [CrossRef] [PubMed]
- Hayes, R.; Imberti, S.; Warr, G.G.; Atkin, R. How water dissolves in protic ionic liquids. Angew. Chem. Int. Ed. 2012, 51, 7468–7471. [Google Scholar] [CrossRef] [PubMed]
- Kikhney, A.G.; Svergun, D.I. A practical guide to small angle X-ray scattering (SAXS) of flexible and intrinsically disordered proteins. Febs Lett. 2015, 589, 2570–2577. [Google Scholar] [CrossRef] [PubMed]
- Demirdöven, N.; Cheatum, C.M.; Chung, H.S.; Khalil, M.; Knoester, J.; Tokmakoff, A. Two-dimensional infrared spectroscopy of antiparallel β-sheet secondary structure. J. Am. Chem Soc. 2004, 126, 7981–7990. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Q.; Broomhall, H.C.; Vieira Veríssimo, N.; Ryan, T.M.; Drummond, C.J.; Pereira, J.F.B.; Greaves, T.L. Protic Ionic Liquid Cation Alkyl Chain Length Effect on Lysozyme Structure. Molecules 2022, 27, 984. https://doi.org/10.3390/molecules27030984
Han Q, Broomhall HC, Vieira Veríssimo N, Ryan TM, Drummond CJ, Pereira JFB, Greaves TL. Protic Ionic Liquid Cation Alkyl Chain Length Effect on Lysozyme Structure. Molecules. 2022; 27(3):984. https://doi.org/10.3390/molecules27030984
Chicago/Turabian StyleHan, Qi, Hayden C. Broomhall, Nathalia Vieira Veríssimo, Timothy M. Ryan, Calum J. Drummond, Jorge F. B. Pereira, and Tamar L. Greaves. 2022. "Protic Ionic Liquid Cation Alkyl Chain Length Effect on Lysozyme Structure" Molecules 27, no. 3: 984. https://doi.org/10.3390/molecules27030984
APA StyleHan, Q., Broomhall, H. C., Vieira Veríssimo, N., Ryan, T. M., Drummond, C. J., Pereira, J. F. B., & Greaves, T. L. (2022). Protic Ionic Liquid Cation Alkyl Chain Length Effect on Lysozyme Structure. Molecules, 27(3), 984. https://doi.org/10.3390/molecules27030984