
Characterization of a Recombinant Laccase B from Trametes hirsuta MX2 and Its Application for Decolorization of Dyes


Abstract
:1. Introduction
2. Results
2.1. Screening and Expression of Laccase Recombinants
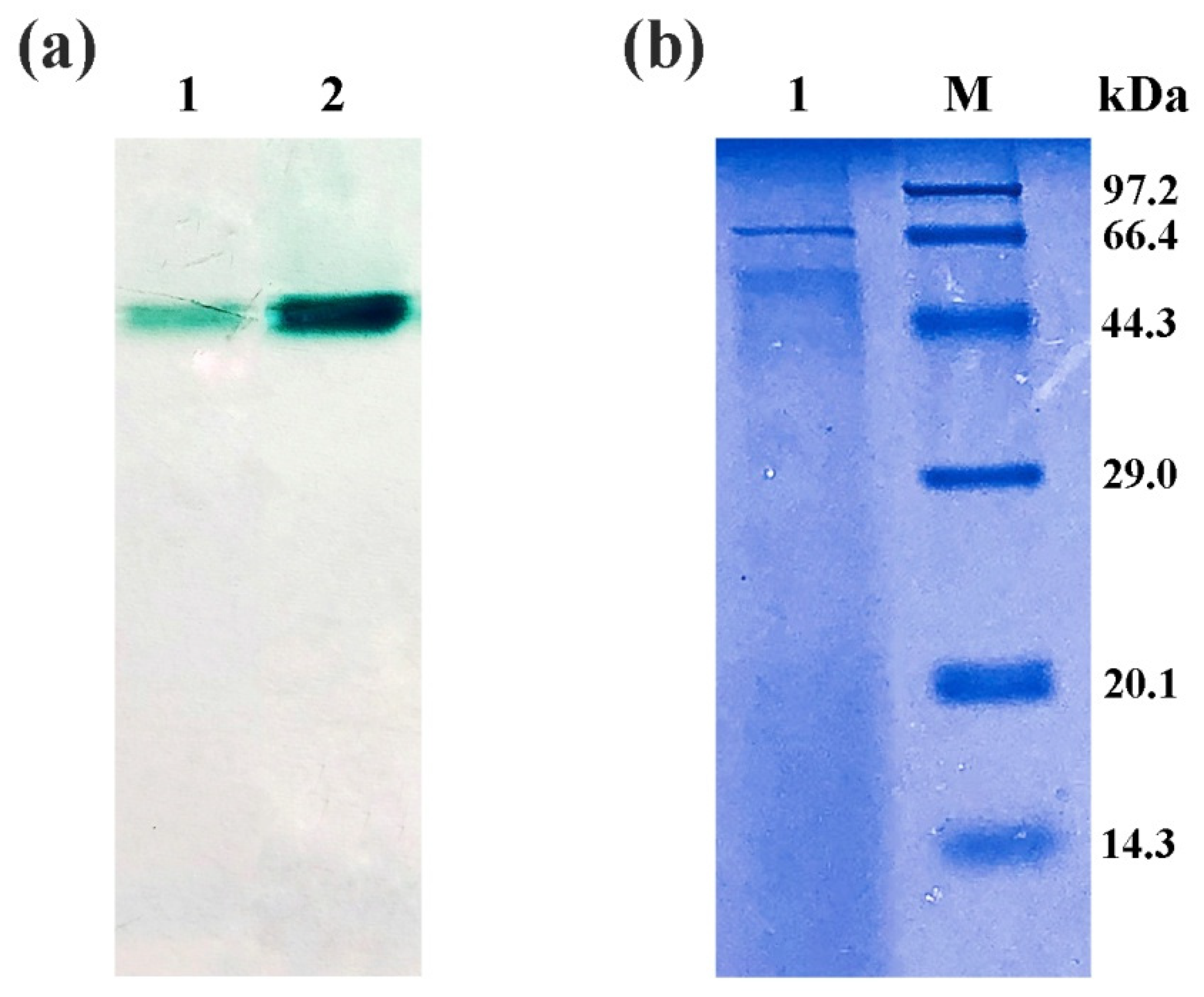
2.2. Purification of Recombinant Laccase
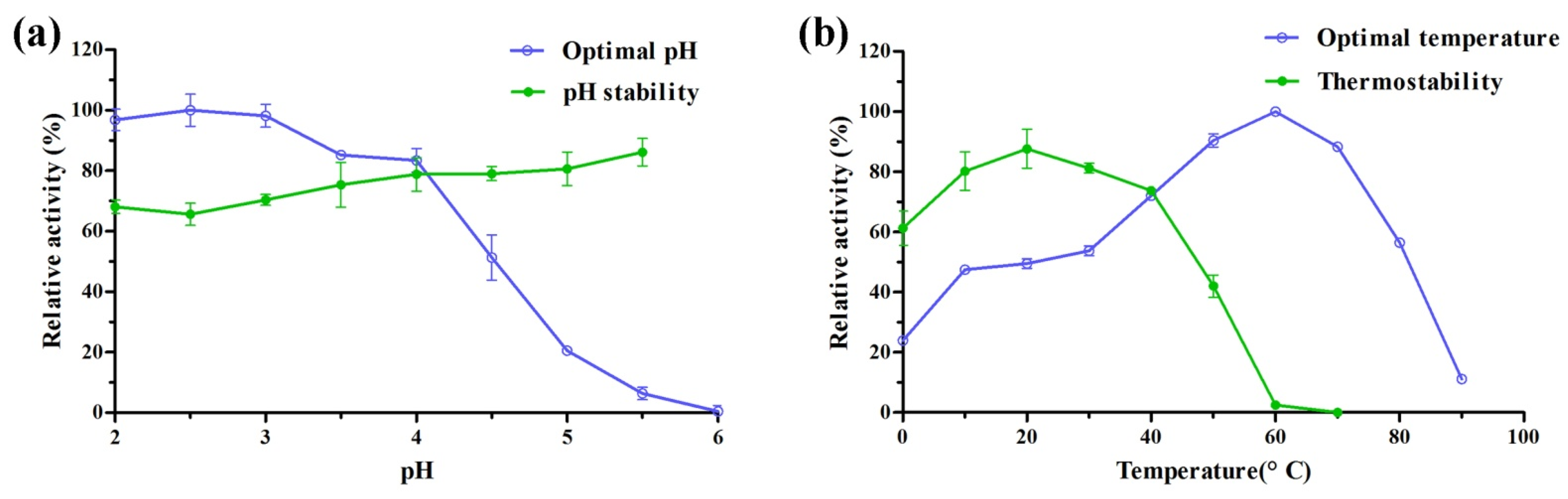
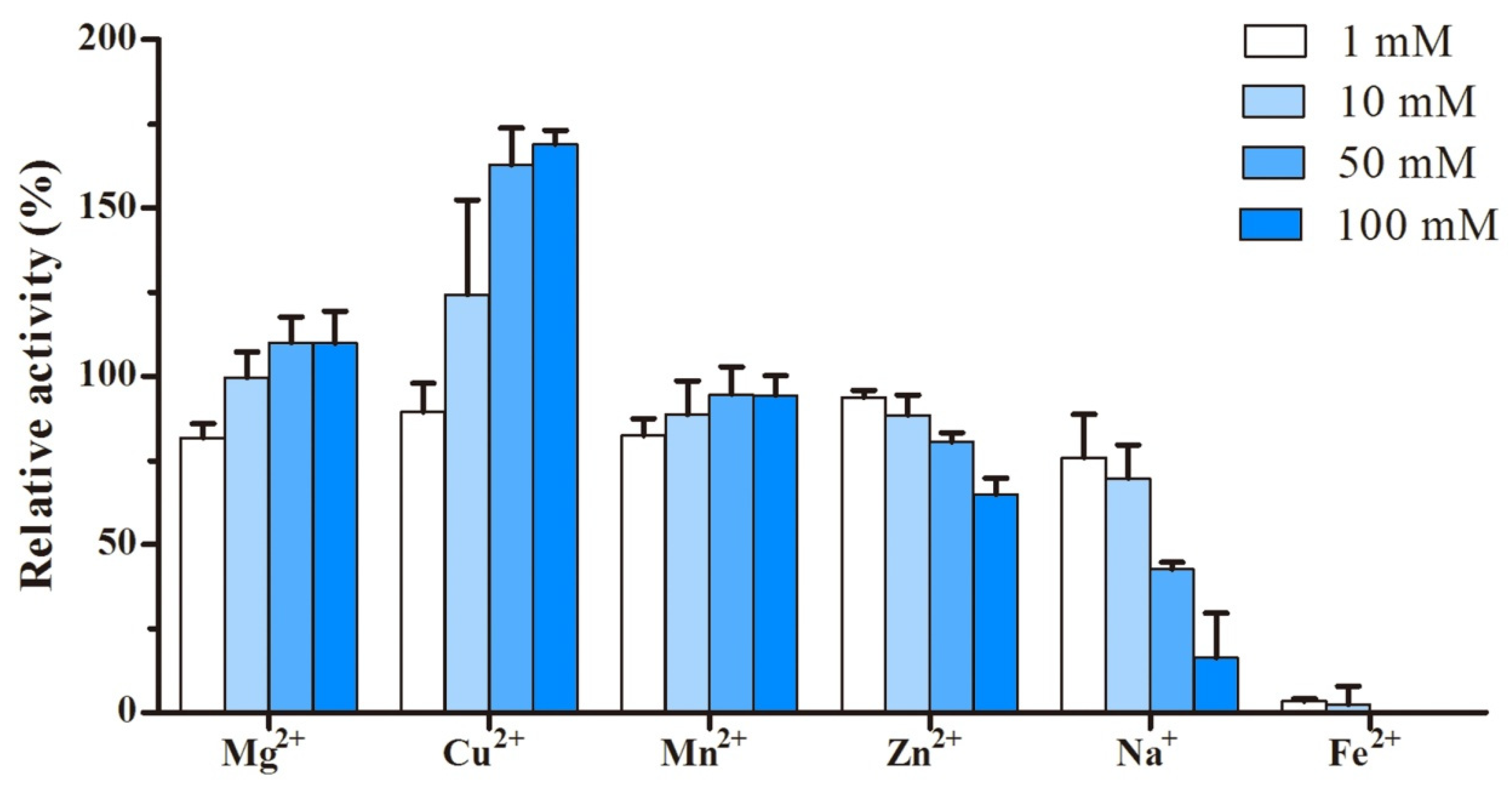
2.3. Characterization of Recombinant Laccase
2.4. Dye Decolorization
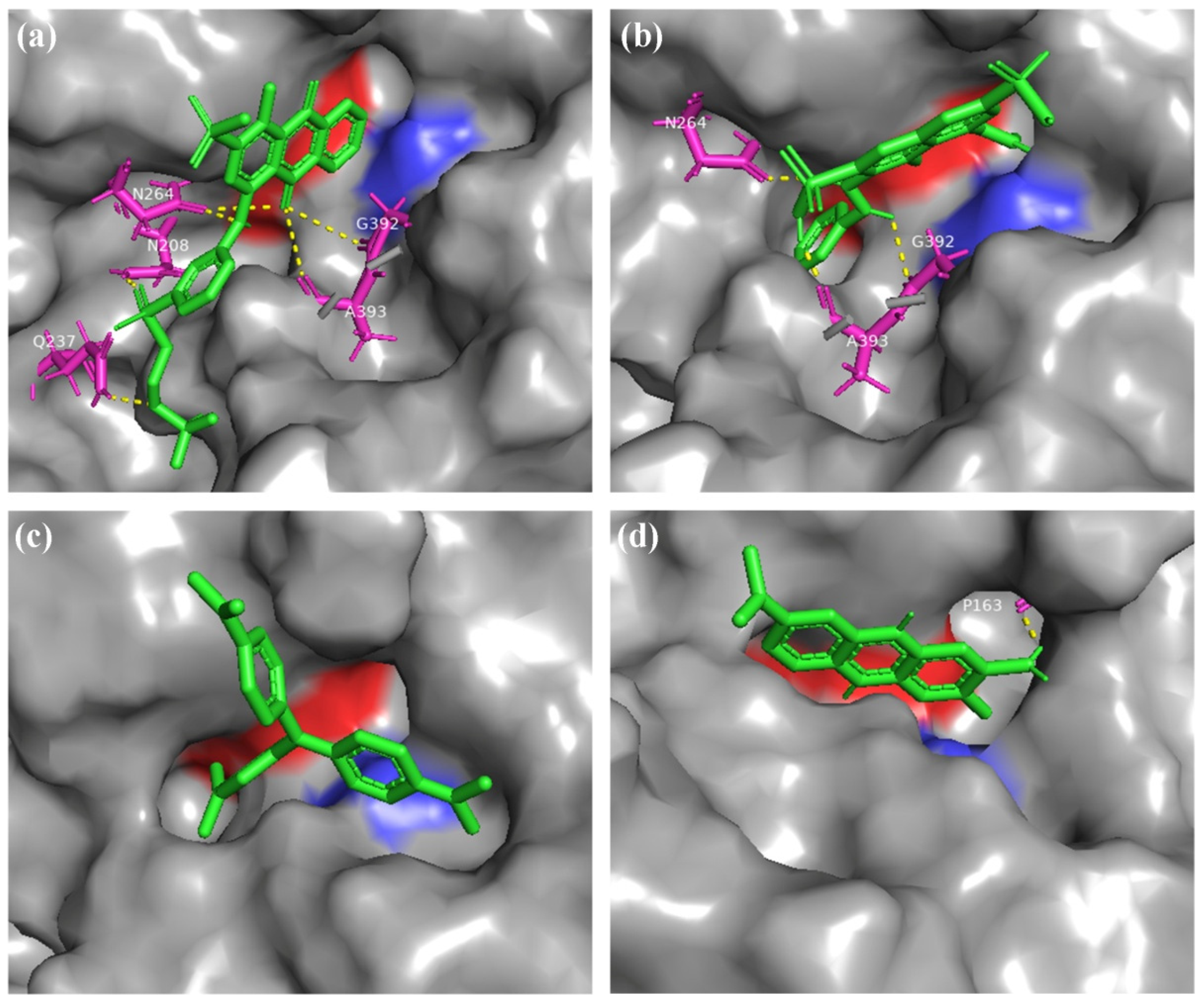
2.5. Molecular Docking
3. Discussions
4. Materials and Methods
4.1. Strains, Vector, and Culture Media
4.2. Cloning of Laccase Gene
4.3. Screening and Expression of Laccase Recombinants
4.4. Laccase Activity Assay
4.5. Purification of Recombinant Laccase
4.6. Characterization of Recombinant Laccase
4.7. Dye Decolorization
4.8. Molecular Docking between Laccase and Dyes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tkaczyk, A.; Mitrowska, K.; Posyniak, A. Synthetic organic dyes as contaminants of the aquatic environment and their implications for ecosystems: A review. Sci. Total. Environ. 2020, 717, 137222. [Google Scholar] [CrossRef] [PubMed]
- Arora, S. Textile dyes: It’s impact on environment and its treatment. J. Bioremed. Biodeg. 2014, 5, e146. [Google Scholar] [CrossRef]
- Selvaraj, V.; Karthika, T.S.; Mansiya, C.; Alagar, M. An over review on recently developed techniques, mechanisms and intermediate involved in the advanced azo dye degradation for industrial applications. J. Mol. Struct. 2021, 1224, 129195. [Google Scholar] [CrossRef]
- Abe, F.R.; Soares, A.M.V.M.; Oliveira, D.P.; Gravato, C. Toxicity of dyes to zebrafish at the biochemical level: Cellular energy allocation and neurotoxicity. Environ. Pollut. 2018, 235, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.S.G.; Tallarico, L.D.; Rosa, J.M.; Suzuki, C.F.; Roubicek, D.A.; Nakano, E.; Borrely, S.I. Multiple adverse effects of textile effluents and reactive Red 239 dye to aquatic organisms. Environ. Sci. Pollut. R. 2021, 28, 63202–63214. [Google Scholar] [CrossRef]
- Robinson, T.; McMullan, G.; Marchant, R.; Nigam, P. Remediation of dyes in textile effluent: A critical review on current treatment technologies with a proposed alternative. Bioresource Technol. 2001, 77, 247–255. [Google Scholar] [CrossRef]
- Singh, R.L.; Singh, P.K.; Singh, R.P. Enzymatic decolorization and degradation of azo dyes—A review. Int. Biodeter. Biodegr. 2015, 104, 21–31. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, C.; Zhu, L.; Zhang, D.; Pan, C. Purification, characterization, and gene cloning of two laccase isoenzymes (Lac1 and Lac2) from Trametes hirsuta MX2 and their potential in dye decolorization. Mol. Biol. Rep. 2020, 47, 477–488. [Google Scholar] [CrossRef]
- Yadav, A.; Yadav, P.; Singh, A.K.; Kumar, V.; Sonawane, V.C.; Markandeya; Bharagava, R.N.; Raj, A. Decolourisation of textile dye by laccase: Process evaluation and assessment of its degradation bioproducts. Bioresour. Technol. 2021, 340, 125591. [Google Scholar] [CrossRef]
- Morsi, R.; Bilal, M.; Iqbal, H.M.N.; Ashraf, S.S. Laccases and peroxidases: The smart, greener and futuristic biocatalytic tools to mitigate recalcitrant emerging pollutants. Sci. Total Environ. 2020, 714, 136572. [Google Scholar] [CrossRef]
- Senthivelan, T.; Kanagaraj, J.; Panda, R.C. Recent trends in fungal laccase for various industrial applications: A eco-friendly approachߞA review. Biotechnol. Bioproc. E. 2016, 21, 19–38. [Google Scholar] [CrossRef]
- Legerska, B.; Chmelova, D.; Ondrejovic, M. Decolourization and detoxification of monoazo dyes by laccase from the white-rot fungus Trametes versicolor. J. Biotechnol. 2018, 285, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Chen, L.; Cao, Z.; Hong, F.F. Enhanced decolourization efficiency of textile dye Reactive Blue 19 in a horizontal rotating reactor using strips of BNC-immobilized laccase: Optimization of conditions and comparison of decolourization efficiency. Biochem. Eng. J. 2020, 156, 107501. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.L.; Hu, B.Y.; Chen, Q.J.; Yang, A.Z.; Wang, Q.Y.; Li, X.F.; Zhang, J.Y.; Zhang, G.Q.; Zhao, Y.C. Purification and characterization of a thermo- and pH-stable laccase from the litter-decomposing fungus Gymnopusluxurians and laccase mediator systems for dye decolorization. Front. Microbiol. 2021, 12, 672620. [Google Scholar] [CrossRef]
- Mate, D.M.; Alcalde, M. Laccase: A multi-purpose biocatalyst at the forefront of biotechnology. Microb. Biotechnol. 2017, 10, 1457–1467. [Google Scholar] [CrossRef]
- Zapata-Castillo, P.; Villalonga-Santana, L.; Islas-Flores, I.; Rivera-Muñoz, G.; Ancona-Escalante, W.; Solís-Pereira, S. Synergistic action of laccases from Trametes hirsuta Bm2 improves decolourization of indigo carmine. Lett. Appl. Microbiol. 2015, 61, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Savinova, O.S.; Moiseenko, K.V.; Vavilova, E.A.; Tyazhelova, T.V.; Vasina, D.V. Properties of two laccases from the Trametes hirsuta 072 multigene family: Twins with different faces. Biochimie 2017, 142, 183–190. [Google Scholar] [CrossRef]
- Couto, S.R. Decolouration of industrial azo dyes by crude laccase from Trametes hirsuta. J. Hazard. Mater. 2007, 148, 768–770. [Google Scholar] [CrossRef]
- Ancona-Escalante, W.; Tapia-Tussell, R.; Pool-Yam, L.; Can-Cauich, A.; Lizama-Uc, G.; Solis-Pereira, S. Laccase-mediator system produced by Trametes hirsuta Bm-2 on lignocellulosic substrate improves dye decolorization. 3 Biotech 2018, 8, 298. [Google Scholar] [CrossRef]
- Anita, S.H.; Ardiati, F.C.; Oktaviani, M.; Sari, F.P.; Nurhayat, O.D.; Ramadhan, K.P.; Yanto, D.H.Y. Immobilization of laccase from Trametes hirsuta EDN 082 in light expanded clay aggregate for decolorization of Remazol Brilliant Blue R dye. Bioresour. Technol. Rep. 2020, 12, 100602. [Google Scholar] [CrossRef]
- Yanto, D.H.Y.; Guntoro, M.A.; Nurhayat, O.D.; Anita, S.H.; Oktaviani, M.; Ramadhan, K.P.; Pradipta, M.F.; Watanabe, T. Biodegradation and biodetoxification of batik dye wastewater by laccase from Trametes hirsuta EDN 082 immobilised on light expanded clay aggregate. 3 Biotech 2021, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Moiseenko, K.V.; Vasina, D.V.; Farukshina, K.T.; Savinova, O.S.; Glazunova, O.A.; Fedorova, T.V.; Tyazhelova, T.V. Orchestration of the expression of the laccase multigene family in white-rot basidiomycete Trametes hirsuta 072: Evidences of transcription level subfunctionalization. Fungal Biol. 2018, 122, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Savinova, O.S.; Moiseenko, K.V.; Vavilova, E.A.; Chulkin, A.M.; Fedorova, T.V.; Tyazhelova, T.V.; Vasina, D.V. Evolutionary relationships between the laccase genes of Polyporales: Orthology-based classification of laccase isozymes and functional insight from Trametes hirsuta. Front. Microbiol. 2019, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Vasina, D.V.; Pavlov, A.R.; Koroleva, O.V. Extracellular proteins of Trametes hirsuta st. 072 induced by copper ions and a lignocellulose substrate. BMC Microbiol. 2016, 16, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Li, W.; Ng, T.B.; Deng, X.; Lin, J.; Ye, X. Laccases: Production, expression regulation and applications in pharmaceutical biodegradation. Front. Microbiol. 2017, 8, 00832. [Google Scholar] [CrossRef] [PubMed]
- Savinova, O.S.; Zorov, I.N.; Vasina, D.V.; Sinitsyn, A.P.; Fedorova, T.V. The minor recombinant laccase isozymes of Trametes hirsuta 072: Preparation and properties. Mosc. Univ. Chem. Bull. 2019, 74, 173–179. [Google Scholar] [CrossRef]
- Yaver, D.S.; Xu, F.; Golightly, E.J.; Brown, K.M.; Brown, S.H.; Rey, M.W.; Schneider, P.; Halkier, T.; Mondorf, K. Purification, characterization, molecular cloning, and expression of two laccase genes from the white rot basidiomycete Trametesvillosa. Appl. Environ. Microb. 1996, 62, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Moiseenko, K.V.; Savinova, O.S.; Vasina, D.V.; Kononikhin, A.S.; Tyazhelova, T.V.; Fedorova, T.V. Laccase isoenzymes of Trametes hirsuta LE-BIN072: Degradation of industrial dyes and secretion under the different induction conditions. Appl. Biochem. Microbiol. 2018, 54, 834–841. [Google Scholar] [CrossRef]
- Bronikowski, A.; Hagedoorn, P.-L.; Koschorreck, K.; Urlacher, V.B. Expression of a new laccase from Moniliophthoraroreri at high levels in Pichia pastoris and its potential application in micropollutant degradation. AMB Express 2017, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Bieler, N.; Kenzom, T.; Chhabra, M.; Ansorge-Schumacher, M.; Mishra, S. Cloning, sequence analysis, expression of Cyathusbulleri laccase in Pichia pastoris and characterization of recombinant laccase. BMC Biotechnol. 2012, 12, 75. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Wang, J.; Yin, Q.; Fang, W.; Xiao, Y.; Fang, Z. Expression of a thermo- and alkali-philic fungal laccase in Pichia pastoris and its application. Protein Express. Purif. 2019, 154, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Antosova, Z.; Sychrova, H. Yeast hosts for the production of recombinant laccases: A review. Mol. Biotechnol. 2016, 58, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Zhou, G.; Peng, C.; Zhang, Y.; Kues, U.; Liu, J.; Xiao, Y.; Fang, Z. The first fungal laccases with an alkaline pH optimum obtained by directed evolution and its application in indigo dye decolorization. AMB Express 2019, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Huy, N.D.; Le, N.T.M.; Chew, K.W.; Park, S.-M.; Show, P.L. Characterization of a recombinant laccase from Fusarium oxysporum HUIB02 for biochemical application on dyes removal. Biochem. Eng. J. 2021, 168, 107958. [Google Scholar] [CrossRef]
- Xu, G.; Wu, Y.; Zhang, Y.; Fang, W.; Xiao, Y.; Fang, Z. Role of N-glycosylation on the specific activity of a Coprinopsiscinerea laccase Lcc9 expressed in Pichia pastoris. J. Biosci. Bioeng. 2019, 128, 518–524. [Google Scholar] [CrossRef]
- Savinova, O.S.; Chulkin, A.M.; Savinova, T.S.; Vavilova, E.A.; Vasina, D.V.; Solyev, P.N.; Fedorova, T.V. Construction of the heterologous laccase producer Aspergillus nidulans lac№4 (argB–) and its application for the progesterone transformation. Appl. Biochem. Microbiol. 2020, 56, 321–328. [Google Scholar] [CrossRef]
- Koschorreck, K.; Richter, S.M.; Swierczek, A.; Beifuss, U.; Schmid, R.D.; Urlacher, V.B. Comparative characterization of four laccases from Trametes versicolor concerning phenolic C–C coupling and oxidation of PAHs. Arch. Biochem. Biophys. 2008, 474, 213–219. [Google Scholar] [CrossRef]
- Yang, X.; Wu, Y.; Zhang, Y.; Yang, E.; Qu, Y.; Xu, H.; Chen, Y.; Irbis, C.; Yan, J. A thermo-active laccase isoenzyme from Trametestrogii and its potential for dye decolorization at high temperature. Front. Microbiol. 2020, 11, 241. [Google Scholar] [CrossRef] [Green Version]
- Navada, K.K.; Kulal, A. Kinetic characterization of purified laccase from Trametes hirsuta: A study on laccase catalyzed biotransformation of 1,4-dioxane. Biotechnol. Lett. 2021, 43, 613–626. [Google Scholar] [CrossRef]
- Vasina, D.V.; Mustafaev, O.N.; Moiseenko, K.V.; Sadovskaya, N.S.; Glazunova, O.A.; Tyurin, A.A.; Fedorova, T.V.; Pavlov, A.R.; Tyazhelova, T.V.; Goldenkova-Pavlova, I.V.; et al. The Trametes hirsuta 072 laccase multigene family: Genes identification and transcriptional analysis under copper ions induction. Biochimie 2015, 116, 154–164. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametespubescens. Bioresour. Technol. 2013, 128, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, W.; Qu, M.; Pan, K.; Zhao, X. Heterologous expression of laccase from Lentinula edodes in Pichia pastoris and its application in degrading rape straw. Front. Microbiol. 2020, 11, 1086. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, S.; Wang, Y.; Song, J.; Wang, H.; Kong, D.; Zhu, J.; Dong, W.; He, M.; Hu, G.; et al. Characterization of a highly thermostable and organic solvent-tolerant copper-containing polyphenol oxidase with dye-decolorizing ability from Kurthiahuakuii LAM0618T. PloS ONE 2016, 11, 164810. [Google Scholar] [CrossRef]
- Keum, Y.S.; Li, Q.X. Fungal laccase-catalyzed degradation of hydroxy polychlorinated biphenyls. Chemosphere 2004, 56, 23–30. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Yaropolov, Y.I. Laccase-mediator systems and their applications: A review. Appl. Biochem. Microbiol. 2007, 43, 523–535. [Google Scholar] [CrossRef]
- Maestre-Reyna, M.; Liu, W.C.; Jeng, W.Y.; Lee, C.C.; Hsu, C.A.; Wen, T.N.; Wang, A.H.J.; Shyur, L.F. Structural and functional roles of glycosylation in fungal laccase from Lentinus sp. PloS ONE 2015, 10, 120601. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, P.; Muhammad, Y.; Tang, Y.; Shao, S.; Gao, Z.; Wang, J.; Wang, R.; Hu, Y.; Kuang, L.; et al. High-density immobilization of laccase on hollow nano-sphere NH2-MIL88 (Fe) host with interfacial defects to improve enzyme activity and stability for remazol brilliant blue R decolorization. Chem. Eng. J. 2021, 405, 127003. [Google Scholar] [CrossRef]
- Cannatelli, M.D.; Ragauskas, A.J. Two decades of laccases: Advancing sustainability in the chemical industry. Chem. Rec. 2017, 17, 122–140. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, D.; Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput.-Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Volume (mL) | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Percent Recovery (%) | Purification Fold |
|---|---|---|---|---|---|---|
| Culture supernatant | 870 | 2235 | 116.8 | 19.1 | 100 | 1 |
| Ultrafiltrate | 186 | 1971 | 53.6 | 36.8 | 88.2 | 1.9 |
| (NH4)2SO4 precipitation | 30 | 484 | 7.8 | 62.1 | 21.7 | 3.3 |
| DEAE-Sepharose FF | 14 | 129 | 1.4 | 92.1 | 5.8 | 4.8 |
| Substrate | Lac1 * | rLac1 | ||||
|---|---|---|---|---|---|---|
| Km (µM) | kcat (s−1) | kcat/Km (M−1 s−1) | Km (µM) | kcat (s−1) | kcat/Km (M−1 s−1) | |
| ABTS | 22.4 | 91.7 | 4.1 × 106 | 28.4 | 343.2 | 1.2 × 107 |
| DMP | 351.7 | 28.4 | 8.1 × 104 | 394.1 | 141.7 | 3.6 × 105 |
| Organic Solvents (v/v) | Relative Activity | ||
|---|---|---|---|
| 5% | 10% | 50% | |
| Acetone | 69.1 ± 7.8 | 44.2 ± 3.9 | 0.4 ± 0.6 |
| Acetonitrile | 80.0 ± 12.3 | 61.3 ± 3.1 | 9.0 ± 0.2 |
| Dimethylsulfoxide | 64.5 ± 6.4 | 43.9 ± 1.4 | 4.3 ± 2.3 |
| Ethanol | 70.0 ± 4.3 | 53.3 ± 2.8 | 4.0 ± 1.0 |
| Methanol | 86.2 ± 7.3 | 62.4 ± 3.1 | 7.0 ± 0.9 |
| Isopropanol | 65.2 ± 9.8 | 42.5 ± 8.8 | 3.0 ± 0.9 |
| Dyes | Mediator | Decolorization Rate (%) |
|---|---|---|
| RBBR | − | 92.6 ± 0.9 |
| + | 99.2 ± 0.6 | |
| AR 1 | − | 15.3 ± 0.8 |
| + | 67.1 ± 0.4 | |
| CV | − | 14.2 ± 0.4 |
| + | 38.9 ± 0.9 | |
| NR | − | 12.3 ± 0.3 |
| + | 52.3 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Huang, Q.; Zhu, L.; Pan, C. Characterization of a Recombinant Laccase B from Trametes hirsuta MX2 and Its Application for Decolorization of Dyes. Molecules 2022, 27, 1581. https://doi.org/10.3390/molecules27051581
Jia Y, Huang Q, Zhu L, Pan C. Characterization of a Recombinant Laccase B from Trametes hirsuta MX2 and Its Application for Decolorization of Dyes. Molecules. 2022; 27(5):1581. https://doi.org/10.3390/molecules27051581
Chicago/Turabian StyleJia, Yitong, Qianqian Huang, Lanlan Zhu, and Chengyuan Pan. 2022. "Characterization of a Recombinant Laccase B from Trametes hirsuta MX2 and Its Application for Decolorization of Dyes" Molecules 27, no. 5: 1581. https://doi.org/10.3390/molecules27051581
APA StyleJia, Y., Huang, Q., Zhu, L., & Pan, C. (2022). Characterization of a Recombinant Laccase B from Trametes hirsuta MX2 and Its Application for Decolorization of Dyes. Molecules, 27(5), 1581. https://doi.org/10.3390/molecules27051581