
Anti-Inflammatory Activity of Bryophytes Extracts in LPS-Stimulated RAW264.7 Murine Macrophages

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Extracts on the Viability of RAW 264.7 Murine Macrophage Cells
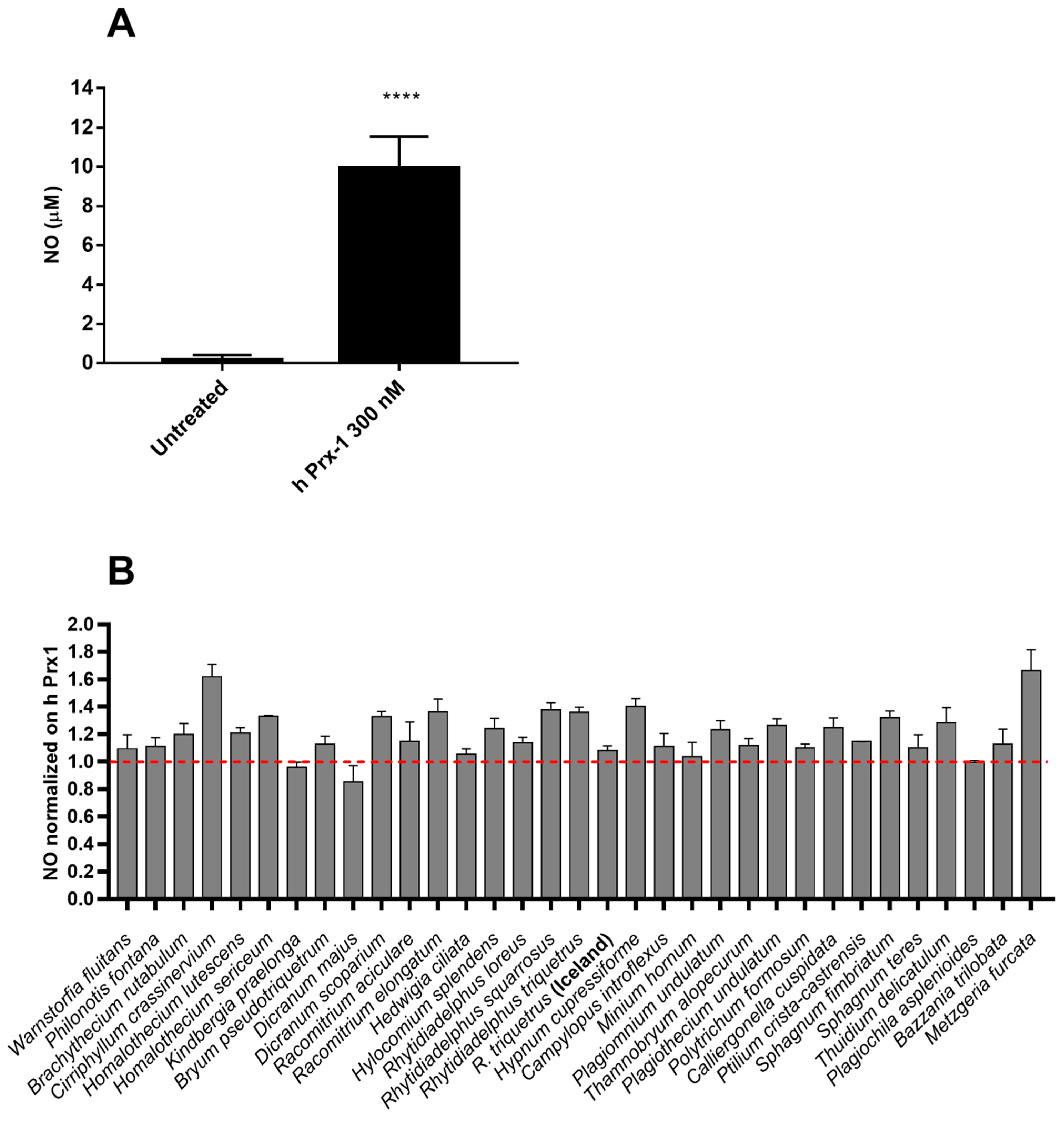
2.2. Anti-Inflammatory Effects of Extracts in hPrx1 or LPS Stimulated RAW264.7 Murine Macrophage Cells
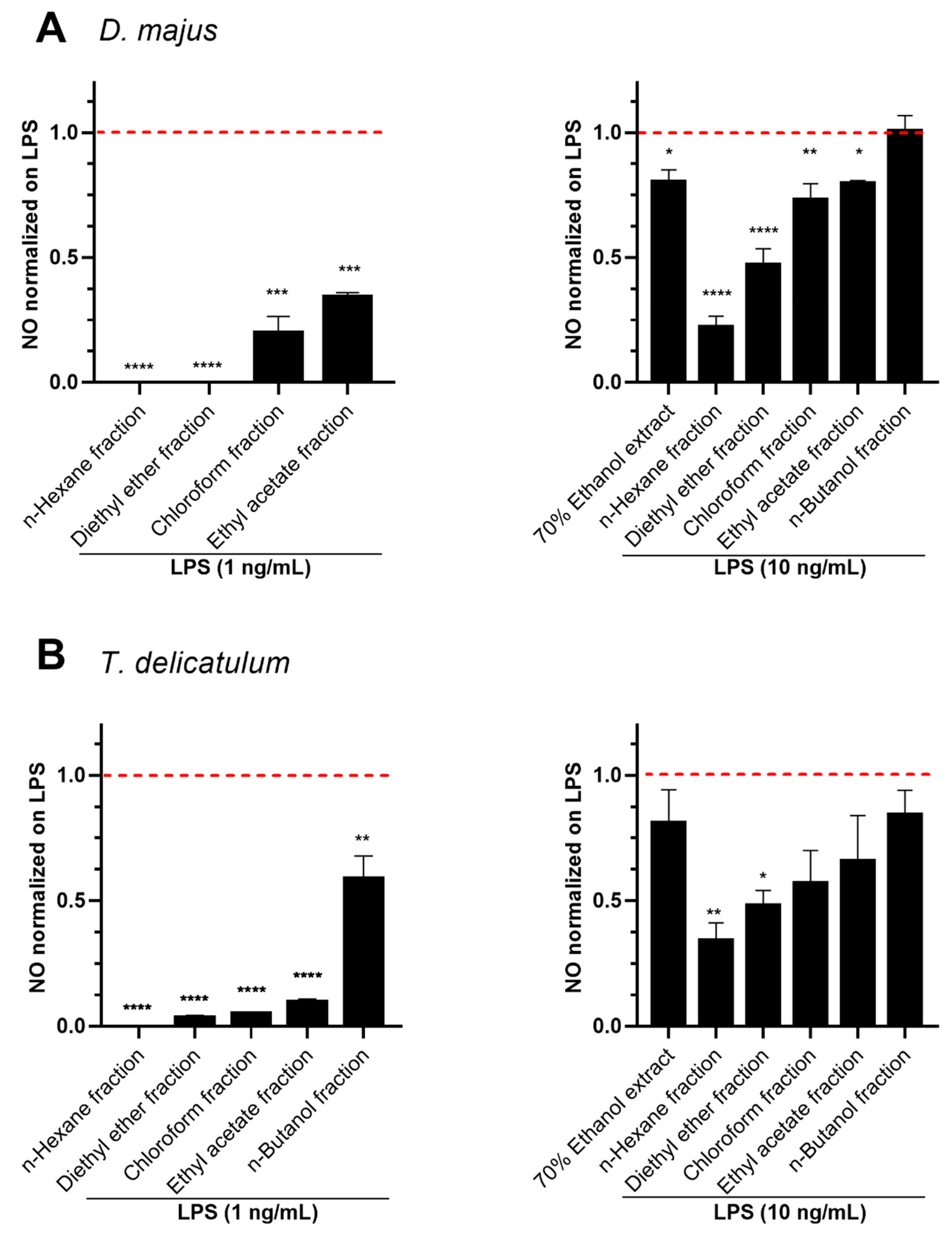
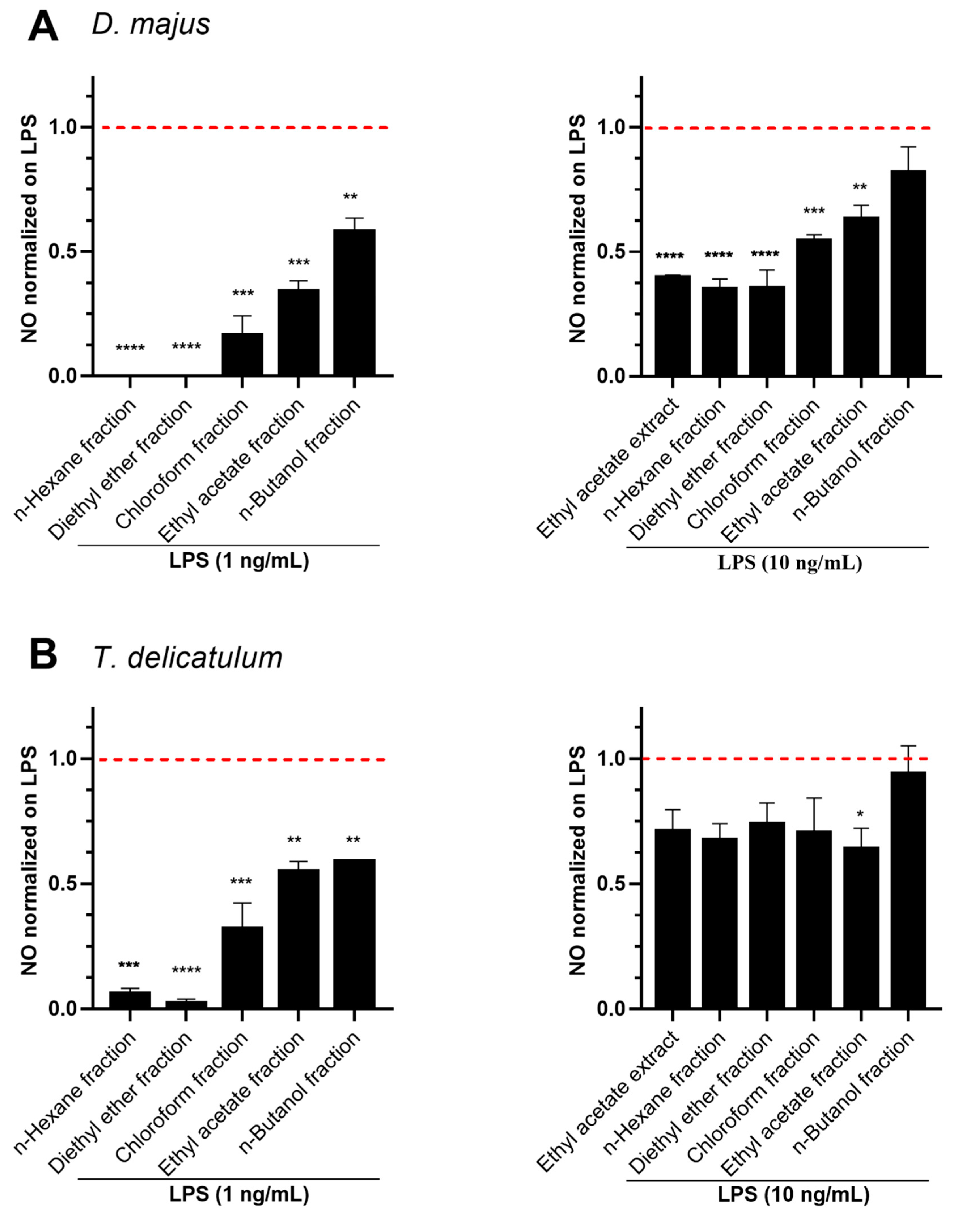
2.3. Anti-Inflammatory Effects of Fractionated Extracts in LPS-Stimulated RAW264.7 Murine Macrophage Cells
3. Material and Methods
3.1. Plant Material
3.2. Recombinant hPrx1 Production
3.3. Extraction of Small Molecules for Screening Activities
3.4. Extraction and Fractionation of Bioactive Bryophytes
3.5. Cell Culture
3.6. Cell Viability and Cytotoxicity Assay
3.7. Measurement of Nitric Oxide
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Sabovljević, M.S.; Sabovljević, A.D.; Ikram, N.; Kusaira, K.; Peramuna, A.; Bae, H.; Simonsen, H.T. Bryophytes-an Emerging Source for Herbal Remedies and Chemical Production. Plant Genet. Resour. Characterisation Util. 2016, 14, 314–327. [Google Scholar] [CrossRef]
- Chandra, S.; Chandra, D.; Barh, A.; Pandey, R.K.; Sharma, I.P. Bryophytes: Hoard of Remedies, an Ethno-Medicinal Review. J. Tradit. Complement. Med. 2017, 7, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, A.; Pascal, A.; Lončarević, I.; Volpatto Marques, R.; Lu, Y.; Miguel, S.; Bourgaud, F.; Thorsteinsdóttir, M.; Cronberg, N.; Becker, J.D.; et al. Natural Products from Bryophytes: From Basic Biology to Biotechnological Applications. CRC Crit. Rev. Plant Sci. 2021, 40, 191–217. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and Physiological Roles of Inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-Sensing Receptors in Sterile Inflammation and Inflammatory Diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms Regulating Skin Immunity and Inflammation. Nat. Rev. Immunol. 2014, 14, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.C.; Weinlich, R.; Bortoluci, K.R. Pattern Recognition Receptors and the Host Cell Death Molecular Machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 493–518. [Google Scholar] [CrossRef] [Green Version]
- Knoops, B.; Becker, S.; Poncin, M.A.; Glibert, J.; Derclaye, S.; Clippe, A.; Alsteens, D. Specific Interactions Measured by AFM on Living Cells between Peroxiredoxin-5 and TLR4: Relevance for Mechanisms of Innate Immunity. Cell Chem. Biol. 2018, 25, 550–559.e3. [Google Scholar] [CrossRef]
- Ishii, T. Close Teamwork between Nrf2 and Peroxiredoxins 1 and 6 for the Regulation of Prostaglandin D 2 and E 2 Production in Macrophages in Acute Inflammation. Free Radic. Biol. Med. 2015, 88, 189–198. [Google Scholar] [CrossRef]
- Knoops, B.; Argyropoulou, V.; Becker, S.; Ferté, L.; Kuznetsova, O. Multiple Roles of Peroxiredoxins in Inflammation. Mol. Cells 2016, 39, 60–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.-X.; Du, J.-R.; Zhou, H.-J.; Liu, D.-L.; Gu, M.-X.; Long, F.-Y. Differences in Proinflammatory Property of Six Subtypes of Peroxiredoxins and Anti-Inflammatory Effect of Ligustilide in Macrophages. PLoS ONE 2016, 11, e0164586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, C. Nitric Oxide Synthase in Innate and Adaptive Immunity: An Update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.C.; Lerner, E. Nitric Oxide Inhibition Strategies. Future Sci. OA 2015, 1, FSO35. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Kim, K.M.; Kim, Y.H.; Im, A.-R.; Lee, J.Y.; Park, B.; Na, M.; Chae, S. The Enhancing Immune Response and Anti-Inflammatory Effects of Anemarrhena Asphodeloides Extract in RAW 264.7 Cells. Adv. Integr. Med. 2019, 6, S76. [Google Scholar] [CrossRef]
- Liu, Y.; Kim, S.; Kim, Y.J.; Perumalsamy, H.; Lee, S.; Hwang, E.; Yi, T.H. Green Synthesis of Gold Nanoparticles Using Euphrasia Officinalis Leaf Extract to Inhibit Lipopolysaccharide-Induced Inflammation through NF-ΚB and JAK/STAT Pathways in RAW 264.7 Macrophages. Int. J. Nanomed. 2019, 14, 2945–2959. [Google Scholar] [CrossRef] [Green Version]
- Karinchai, J.; Budluang, P.; Temviriyanukul, P.; Ting, P.; Nuchuchua, O.; Wongnoppavich, A.; Imsumran, A.; Pitchakarn, P. Bioassay-Guided Study of the Anti-Inflammatory Effect of Anoectochilus Burmannicus Ethanolic Extract in RAW 264.7 Cells. J. Ethnopharmacol. 2021, 280, 114452. [Google Scholar] [CrossRef]
- Ahmad, W.; Jantan, I.; Kumolosasi, E.; Haque, M.A.; Bukhari, S.N.A. Immunomodulatory Effects of Tinospora Crispa Extract and Its Major Compounds on the Immune Functions of RAW 264.7 Macrophages. Int. Immunopharmacol. 2018, 60, 141–151. [Google Scholar] [CrossRef]
- Shichita, T.; Hasegawa, E.; Kimura, A.; Morita, R.; Sakaguchi, R.; Takada, I.; Sekiya, T.; Ooboshi, H.; Kitazono, T.; Yanagawa, T.; et al. Peroxiredoxin Family Proteins Are Key Initiators of Post-Ischemic Inflammation in the Brain. Nat. Med. 2012, 18, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.L.; Zhao, L.X.; Zhang, S.; Du, J.R. Peroxiredoxin 1-Mediated Activation of TLR4/NF-ΚB Pathway Contributes to Neuroinflammatory Injury in Intracerebral Hemorrhage. Int. Immunopharmacol. 2016, 41, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.S.; Jeong, J.W.; Kim, S.O.; Kim, G.Y.; Kim, B.W.; Kim, C.M.; Seo, Y.B.; Kim, W.Y.; Lee, S.Y.; Jo, K.H.; et al. Anti-Inflammatory Potential of Peat Moss Extracts in Lipopolysaccharide- Stimulated RAW 264.7 Macrophages. Int. J. Mol. Med. 2014, 34, 1101–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, W.; Park, S.J.; Shin, J.S.; Noh, Y.S.; Cho, E.J.; Nam, J.H.; Lee, K.T. Anti-Inflammatory Effects of the Methanol Extract of Polytrichum Commune via NF-ΚB Inactivation in RAW 264.7 Macrophage Cells. Biomol. Ther. 2008, 16, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Commisso, M.; Guarino, F.; Marchi, L.; Muto, A.; Piro, A.; Degola, F. Bryo-Activities: A Review on How Bryophytes Are Contributing to the Arsenal of Natural Bioactive Compounds against Fungi. Plants 2021, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Gorzolka, K.; Bruelheide, H.; Neumann, S. Seasonal Variation of Secondary Metabolites in Nine Different Bryophytes. Ecol. Evol. 2018, 8, 9105–9117. [Google Scholar] [CrossRef]
- Sevim, E.; Baş, Y.; Çelik, G.; Pinarbaş, M.; Bozdeveci, A.; Özdemir, T.; Akpinar, R.; Yayli, N.; Karaoğlu, Ş.A. Antibacterial Activity of Bryophyte Species against Paenibacillus Larvae Isolates. Turk. J. Vet. Anim. Sci. 2017, 41, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Borel, C.; Welti, D.H.; Fernandez, I.; Colmenares NestecLtd, M. Dicranin, an Antimicrobial and 15-Lipoxygenase Inhibitor from the Moss Dicranum Scoparium. J. Nat. Prod. 1993, 56, 1071–1077. [Google Scholar] [CrossRef]
- Kühn, H.; O’Donnell, V.B. Inflammation and Immune Regulation by 12/15-Lipoxygenases. Prog. Lipid Res. 2006, 45, 334–356. [Google Scholar] [CrossRef] [PubMed]
- Bodade, R.G.; Borkar, P.S.; Saiful, A.M.; Khobragade, C.N. In Vitro Screening of Bryophytes for Antimicrobial Activity. J. Med. Plants 2008, 7, 23–28. [Google Scholar]
- Available online: https://digitalcommons.mtu.edu/bryophyte-ecology5/4 (accessed on 15 March 2022).
- Greeshma, G.M.; Murugan, K. Mechanism of Anti-Inflammatory Potential of Terpenoids from Thuidium Tamariscellum (C. Muell.) Bosch. & Sande-Lac. A Moss Using Animal and Macrophage Models. Trends Biosci. 2018, 11, 1095–1101. [Google Scholar]
- Kriznik, A.; Libiad, M.; Le Cordier, H.; Boukhenouna, S.; Toledano, M.B.; Rahuel-Clermont, S. Dynamics of a Key Conformational Transition in the Mechanism of Peroxiredoxin Sulfinylation. ACS Catal. 2020, 10, 3326–3339. [Google Scholar] [CrossRef] [PubMed]
- Klavina, L. A Study on Bryophyte Chemical Composition-Search for New Applications. Agron. Res. 2015, 13, 969–978. [Google Scholar]
- Marques, R.V.; Guillaumin, A.; Abdelwahab, A.B.; Salwinski, A.; Gotfredsen, C.H.; Bourgaud, F.; Enemark-Rasmussen, K.; Miguel, S.; Simonsen, H.T. Collagenase and Tyrosinase Inhibitory Effect of Isolated Constituents from the Moss Polytrichum Formosum. Plants 2021, 10, 1271. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions Yield (%) | D. majus (70% Ethanol Crude Extract) | D. majus (Ethyl Acetate Crude Extract) | T. delicatulum (70% Ethanol Crude Extract) | T. delicatulum (Ethyl Acetate Crude Extract) |
|---|---|---|---|---|
| n-Hexane | 7 | 13 | 5 | 8 |
| Diethyl Ether | 16 | 30 | 56 | 29 |
| Chloroform | 15 | 16 | 10 | 18 |
| Ethyl Acetate | 7 | 9 | 8 | 19 |
| n-Butanol | 12 | 10 | 10 | 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, R.V.; Sestito, S.E.; Bourgaud, F.; Miguel, S.; Cailotto, F.; Reboul, P.; Jouzeau, J.-Y.; Rahuel-Clermont, S.; Boschi-Muller, S.; Simonsen, H.T.; et al. Anti-Inflammatory Activity of Bryophytes Extracts in LPS-Stimulated RAW264.7 Murine Macrophages. Molecules 2022, 27, 1940. https://doi.org/10.3390/molecules27061940
Marques RV, Sestito SE, Bourgaud F, Miguel S, Cailotto F, Reboul P, Jouzeau J-Y, Rahuel-Clermont S, Boschi-Muller S, Simonsen HT, et al. Anti-Inflammatory Activity of Bryophytes Extracts in LPS-Stimulated RAW264.7 Murine Macrophages. Molecules. 2022; 27(6):1940. https://doi.org/10.3390/molecules27061940
Chicago/Turabian StyleMarques, Raíssa Volpatto, Stefania Enza Sestito, Frédéric Bourgaud, Sissi Miguel, Frédéric Cailotto, Pascal Reboul, Jean-Yves Jouzeau, Sophie Rahuel-Clermont, Sandrine Boschi-Muller, Henrik Toft Simonsen, and et al. 2022. "Anti-Inflammatory Activity of Bryophytes Extracts in LPS-Stimulated RAW264.7 Murine Macrophages" Molecules 27, no. 6: 1940. https://doi.org/10.3390/molecules27061940
APA StyleMarques, R. V., Sestito, S. E., Bourgaud, F., Miguel, S., Cailotto, F., Reboul, P., Jouzeau, J. -Y., Rahuel-Clermont, S., Boschi-Muller, S., Simonsen, H. T., & Moulin, D. (2022). Anti-Inflammatory Activity of Bryophytes Extracts in LPS-Stimulated RAW264.7 Murine Macrophages. Molecules, 27(6), 1940. https://doi.org/10.3390/molecules27061940