
Curcumin–Induced Stabilization of Protein–Based Nano-Delivery Vehicles Reduces Disruption of Zwitterionic Giant Unilamellar Vesicles




Abstract
:1. Introduction
2. Results and Discussion
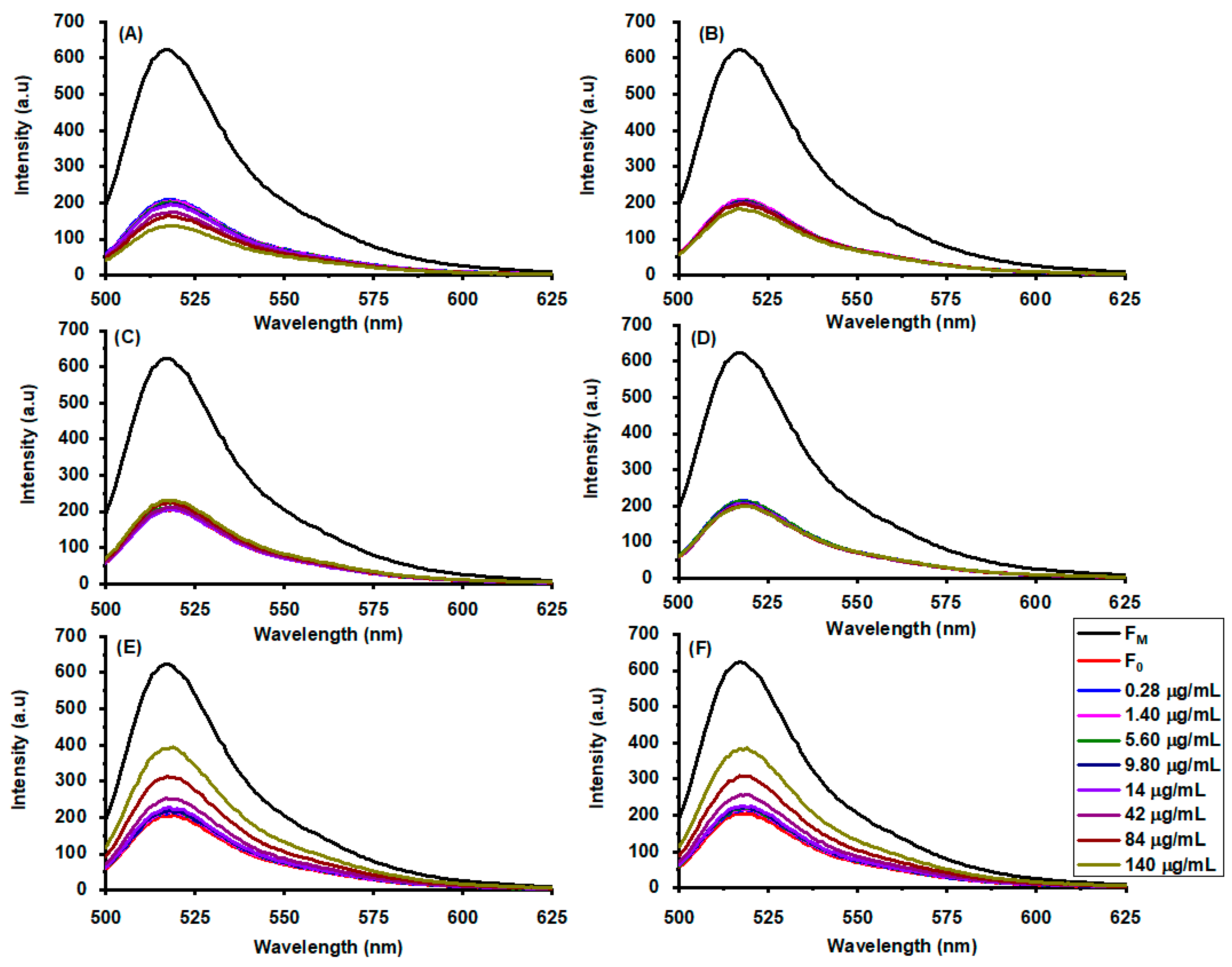
2.1. Nanoparticle-Induced Giant Unilamellar Vesicle Disruption at Physiological pH
2.2. Influence of Nanoparticle-GUV Interaction on the Size, Zeta Potential and Polydispersity Index of the Vesicles
2.3. Morphological Properties of the GUV after Interaction with Nanoparticles
3. Materials and Methods
3.1. Chemical Materials
3.2. Synthesis of Hollow and Curcumin-Loaded Protein Nanoparticles
3.2.1. Preparation of Reaction Solutions
3.2.2. Preparation of Curcumin-Loaded Native and Succinylated Pea Protein Nano-Complexes
3.2.3. Preparation of Curcumin-Loaded Protein-Chitosan Nano-Complexes
3.2.4. Preparation of Native/Succinylated Pea Protein-Chitosan Hollow Nano-Complexes
3.3. Interactions of the Protein Nanoparticles with Calcein-Loaded GUV
3.3.1. Preparation of Empty and Calcein-Loaded Giant Unilamellar Vesicles
3.3.2. Zwitterionic GUV Leakage Assay
3.4. Morphological Characterization of the Structural Integrity of Zwitterionic GUV after Interaction with Nanoparticles
3.5. Dynamic Light Scattering Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Okagu, O.D.; Jin, J.; Udenigwe, C.C. Impact of succinylation on pea protein-curcumin interaction, polyelectrolyte complexation with chitosan, and gastrointestinal release of curcumin in loaded-biopolymer nano-complexes. J. Mol. Liq. 2020, 325, 115248. [Google Scholar] [CrossRef]
- Guo, Q.; Bayram, I.; Zhang, W.; Su, J.; Shu, X.; Yuan, F.; Mao, L.; Gao, Y. Fabrication and characterization of curcumin-loaded pea protein isolate-surfactant complexes at neutral pH. Food Hydrocoll. 2021, 111, 106214. [Google Scholar] [CrossRef]
- Okagu, O.D.; Udenigwe, C.C. Molecular Interactions of Pea Globulin, Albumin and Glutelin With Curcumin: Formation and Gastric Release Mechanisms of Curcumin-loaded Bio-nanocomplexes. Food Biophys. 2022, 17, 10–25. [Google Scholar] [CrossRef]
- Okagu, O.D.; Verma, O.; McClements, D.J.; Udenigwe, C.C. Utilization of insect proteins to formulate nutraceutical delivery systems: Encapsulation and release of curcumin using mealworm protein-chitosan nano-complexes. Int. J. Biol. Macromol. 2020, 151, 333–343. [Google Scholar] [CrossRef]
- Wang, F.; Yang, Y.; Ju, X.; Udenigwe, C.C.; He, R. Polyelectrolyte Complex Nanoparticles from Chitosan and Acylated Rapeseed Cruciferin Protein for Curcumin Delivery. J. Agric. Food Chem. 2018, 66, 2685–2693. [Google Scholar] [CrossRef] [PubMed]
- Sneharani, A.H. Curcumin–sunflower protein nanoparticles—A potential antiinflammatory agent. J. Food Biochem. 2019, 43, e12909. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wang, C.; Liu, X.; Liao, W.; Zhang, L.; Chen, S.; Liu, J.; Mao, L.; Yuan, F.; Gao, Y. Effects of microfluidization and thermal treatment on the characterization and digestion of curcumin loaded protein-polysaccharide-tea saponin complex nanoparticles. Food Funct. 2021, 12, 1192–1206. [Google Scholar] [CrossRef]
- Mirpoor, S.F.; Hosseini, S.M.H.; Yousefi, G.H. Mixed biopolymer nanocomplexes conferred physicochemical stability and sustained release behavior to introduced curcumin. Food Hydrocoll. 2017, 71, 216–224. [Google Scholar] [CrossRef]
- McClements, D.J.; Xiao, H. Is nano safe in foods? Establishing the factors impacting the gastrointestinal fate and toxicity of organic and inorganic food-grade nanoparticles. NPJ Sci. Food 2017, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.Y.; Al-Salami, H.; Dass, C.R. Cellular assays and applied technologies for characterisation of orally administered protein nanoparticles: A systematic review. J. Drug Target 2020, 28, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, M.; Sun, Y.; Zhang, L. A cell-penetrating peptide conjugated carboxymethyl-β-cyclodextrin to improve intestinal absorption of insulin. Int. J. Biol. Macromol. 2018, 111, 685–695. [Google Scholar] [CrossRef]
- Hirano, A.; Uda, K.; Maeda, Y.; Akasaka, T.; Shiraki, K. One-dimensional protein-based nanoparticles induce lipid bilayer disruption: Carbon nanotube conjugates and amyloid fibrils. Langmuir 2010, 26, 17256–17259. [Google Scholar] [CrossRef] [PubMed]
- Araújo, F.; Shrestha, N.; Granja, P.L.; Hirvonen, J.; Santos, H.A.; Sarmento, B. Safety and toxicity concerns of orally delivered nanoparticles as drug carriers. Expert Opin. Drug Metab. Toxicol. 2015, 11, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Van Os, W.L.; Tian, X.; Zu, G.; Ribovski, L.; Bron, R.; Bussmann, J.; Kros, A.; Liu, Y.; Zuhorn, I.S. Development of curcumin-loaded zein nanoparticles for transport across the blood-brain barrier and inhibition of glioblastoma cell growth. Biomater. Sci. 2021, 9, 7092–7103. [Google Scholar] [CrossRef]
- Saleh, T.; Soudi, T.; Shojaosadati, S.A. Aptamer functionalized curcumin-loaded human serum albumin (HSA) nanoparticles for targeted delivery to HER-2 positive breast cancer cells. Int. J. Biol. Macromol. 2019, 130, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Saleh, T.; Soudi, T.; Shojaosadati, S.A. Redox responsive curcumin-loaded human serum albumin nanoparticles: Preparation, characterization and in vitro evaluation. Int. J. Biol. Macromol. 2018, 114, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Larson, R.G. Lipid bilayer curvature and pore formation induced by charged linear polymers and dendrimers: The effects of charge density, concentration, molecular size and shape. In Proceedings of the AIChE Annual Meeting, Conference Proceedings, Philadelphia, PA, USA, 16–21 November 2008. [Google Scholar]
- Mecke, A.; Majoros, I.J.; Patri, A.K.; Baker, J.R.; Banaszak Holl, M.M.; Orr, B.G. Lipid bilayer disruption by polycationic polymers: The roles of size and chemical functional group. Langmuir 2005, 21, 10348–10354. [Google Scholar] [CrossRef] [PubMed]
- Jing, B.; Zhu, Y. Disruption of supported lipid bilayers by semihydrophobic nanoparticles. J. Am. Chem. Soc. 2011, 133, 10983–10989. [Google Scholar] [CrossRef]
- Mantil, E.; Buznytska, I.; Daly, G.; Ianoul, A.; Avis, T.J. Role of Lipid Composition in the Interaction and Activity of the Antimicrobial Compound Fengycin with Complex Membrane Models. J. Membr. Biol. 2019, 252, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Ambroggio, E.E.; Separovic, F.; Bowie, J.H.; Fidelio, G.D.; Bagatolli, L.A. Direct visualization of membrane leakage induced by the antibiotic peptides: Maculatin, citropin, and aurein. Biophys. J. 2005, 89, 1874–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, A.; Yoshikawa, H.; Matsushita, S.; Yamada, Y.; Shiraki, K. Adsorption and disruption of lipid bilayers by nanoscale protein aggregates. Langmuir 2012, 28, 3887–3895. [Google Scholar] [CrossRef] [PubMed]
- Alkhammash, H.I.; Li, N.; Berthier, R.; De Planque, M.R.R. Native silica nanoparticles are powerful membrane disruptors. Phys. Chem. Chem. Phys. 2015, 17, 15547–15560. [Google Scholar] [CrossRef] [Green Version]
- Aydin, F.; Dutt, M. Surface Reconfiguration of Binary Lipid Vesicles via Electrostatically Induced Nanoparticle Adsorption. J. Phys. Chem. B 2016, 120, 6646–6656. [Google Scholar] [CrossRef] [PubMed]
- Di Silvio, D.; Maccarini, M.; Parker, R.; Mackie, A.; Fragneto, G.; Baldelli Bombelli, F. The effect of the protein corona on the interaction between nanoparticles and lipid bilayers. J. Colloid Interface Sci. 2017, 504, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, L.; Sung, C.B.; Granick, S. Nanoparticle-induced surface reconstruction of phospholipid membranes. Proc. Natl. Acad. Sci. USA 2008, 105, 18171–18175. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.; Král, P. Nanoparticles Self-Assembly within Lipid Bilayers. ACS Omega 2018, 3, 10631–10637. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, S.; Ho, S.; Ros, U.; Lanio, M.E.; Alvarez, C.; van der Goot, F.G. Damage of eukaryotic cells by the pore-forming toxin sticholysin II: Consequences of the potassium efflux. Biochim. Biophys. Acta Biomembr. 2017, 1859, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Wolfram, J.; Suri, K.; Yang, Y.; Shen, J.; Celia, C.; Fresta, M.; Zhao, Y.; Shen, H.; Ferrari, M. Shrinkage of pegylated and non-pegylated liposomes in serum. Colloids Surf. B Biointerfaces 2014, 114, 294–300. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Size (nm) | PDI | Zeta Potential (mV) |
|---|---|---|---|
| lGUV | 1933.3 ± 29.0 b | 0.30 ± 0.03 a,b,c | −5.7 ± 0.2 b |
| lGUV + CUR/PPI | 1091.3 ± 5.0 e | 0.33 ± 0.04 a,b,c | −10.0 ± 0.1 e,f |
| lGUV + CUR/SPPI | 1028.7 ± 26.2 e | 0.25 ± 0.02 a,b,c | −11.5 ± 0.3 f,g |
| lGUV + CUR/PPI/CHI | 1689.7 ± 55.2 c | 0.42 ± 0.10 a,b | −9.6 ± 0.3 d,e |
| lGUV + CUR/SPPI/CHI | 2222.7 ± 36.4 a | 0.18 ± 0.05 b,c | −7.9 ± 0.4 c,d |
| lGUV + PPI/CHI | 1464.7 ± 64.8 d | 0.41 ± 0.06 a,b | −11.8 ± 0.2 g |
| lGUV + SPPI/CHI | 1551.7 ± 20.0 c,d | 0.43 ± 0.05 a | −7.4 ± 0.2 b,c |
| lGUV + Triton X-100 | 10.7 ± 0.0 f | 0.10 ± 0.00 c | −1.0 ± 0.8 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okagu, O.D.; Abioye, R.O.; Udenigwe, C.C. Curcumin–Induced Stabilization of Protein–Based Nano-Delivery Vehicles Reduces Disruption of Zwitterionic Giant Unilamellar Vesicles. Molecules 2022, 27, 1941. https://doi.org/10.3390/molecules27061941
Okagu OD, Abioye RO, Udenigwe CC. Curcumin–Induced Stabilization of Protein–Based Nano-Delivery Vehicles Reduces Disruption of Zwitterionic Giant Unilamellar Vesicles. Molecules. 2022; 27(6):1941. https://doi.org/10.3390/molecules27061941
Chicago/Turabian StyleOkagu, Ogadimma D., Raliat O. Abioye, and Chibuike C. Udenigwe. 2022. "Curcumin–Induced Stabilization of Protein–Based Nano-Delivery Vehicles Reduces Disruption of Zwitterionic Giant Unilamellar Vesicles" Molecules 27, no. 6: 1941. https://doi.org/10.3390/molecules27061941
APA StyleOkagu, O. D., Abioye, R. O., & Udenigwe, C. C. (2022). Curcumin–Induced Stabilization of Protein–Based Nano-Delivery Vehicles Reduces Disruption of Zwitterionic Giant Unilamellar Vesicles. Molecules, 27(6), 1941. https://doi.org/10.3390/molecules27061941