
Dihydrohomoplantagin and Homoplantaginin, Major Flavonoid Glycosides from Salvia plebeia R. Br. Inhibit oxLDL-Induced Endothelial Cell Injury and Restrict Atherosclerosis via Activating Nrf2 Anti-Oxidation Signal Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
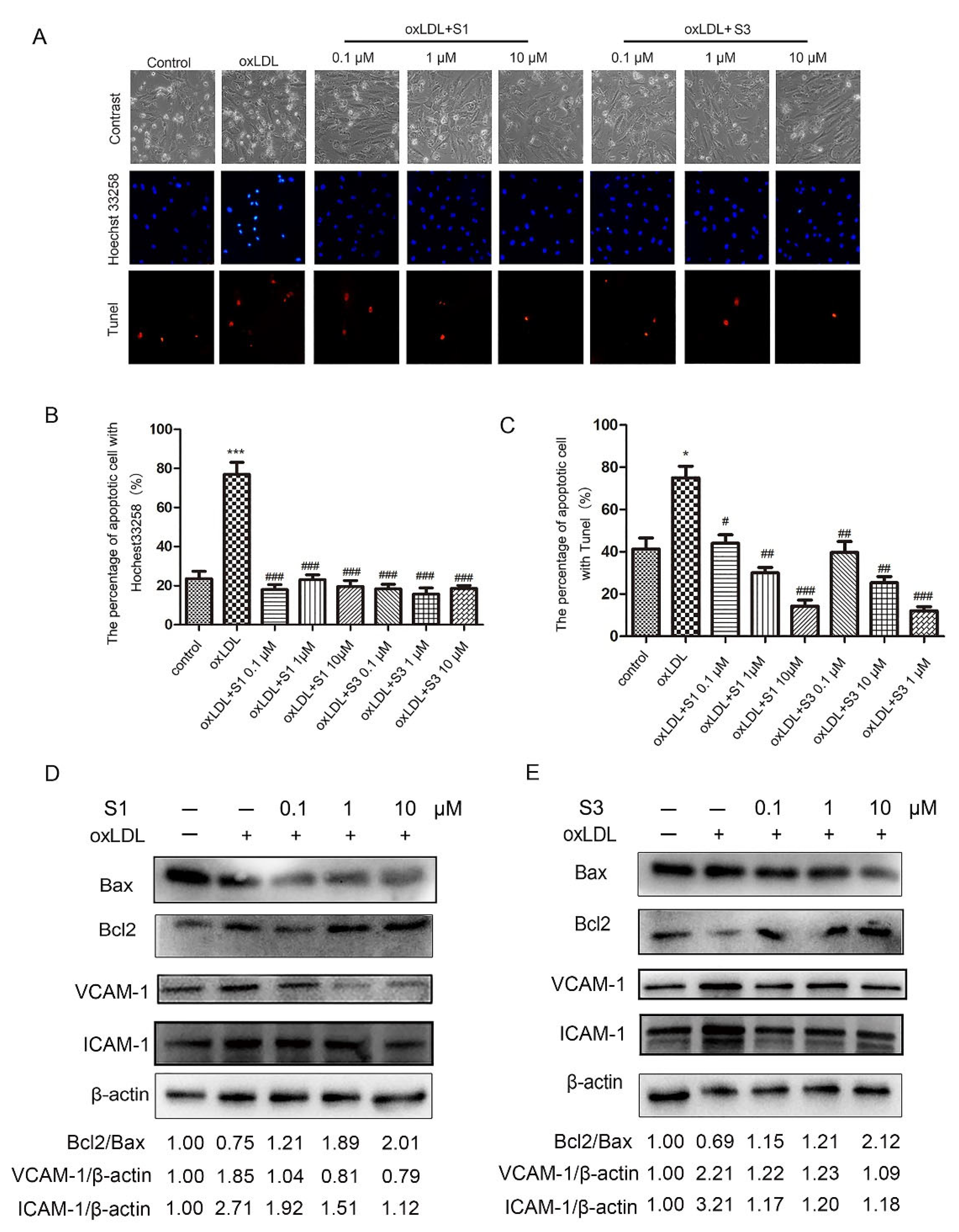
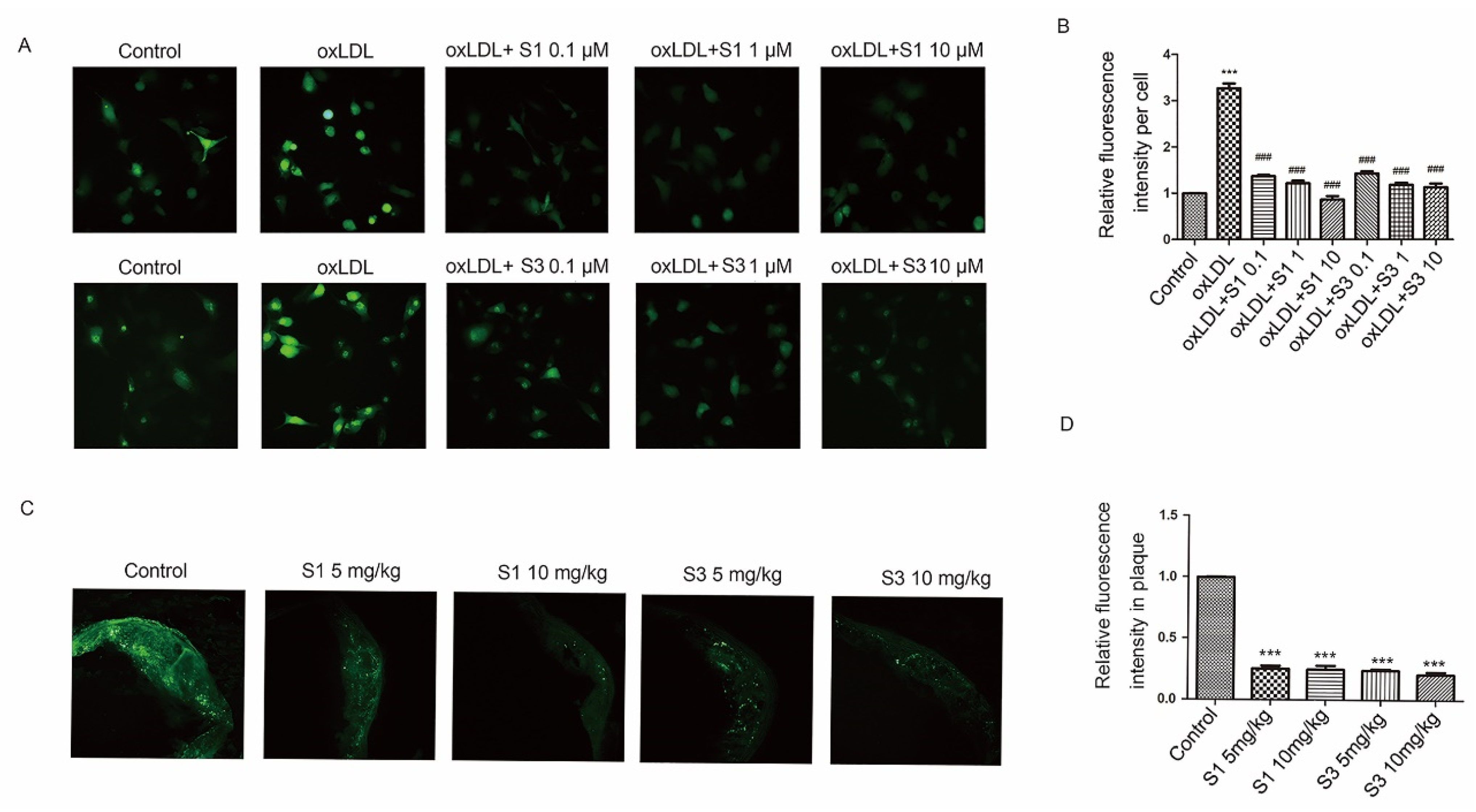
2.1. Dihydrohomoplantagin and Homoplantaginin Inhibited oxLDL-Induced Apoptosis and the Increase in ICAM-1 and VCAM-1 Protein Levels in HUVECs
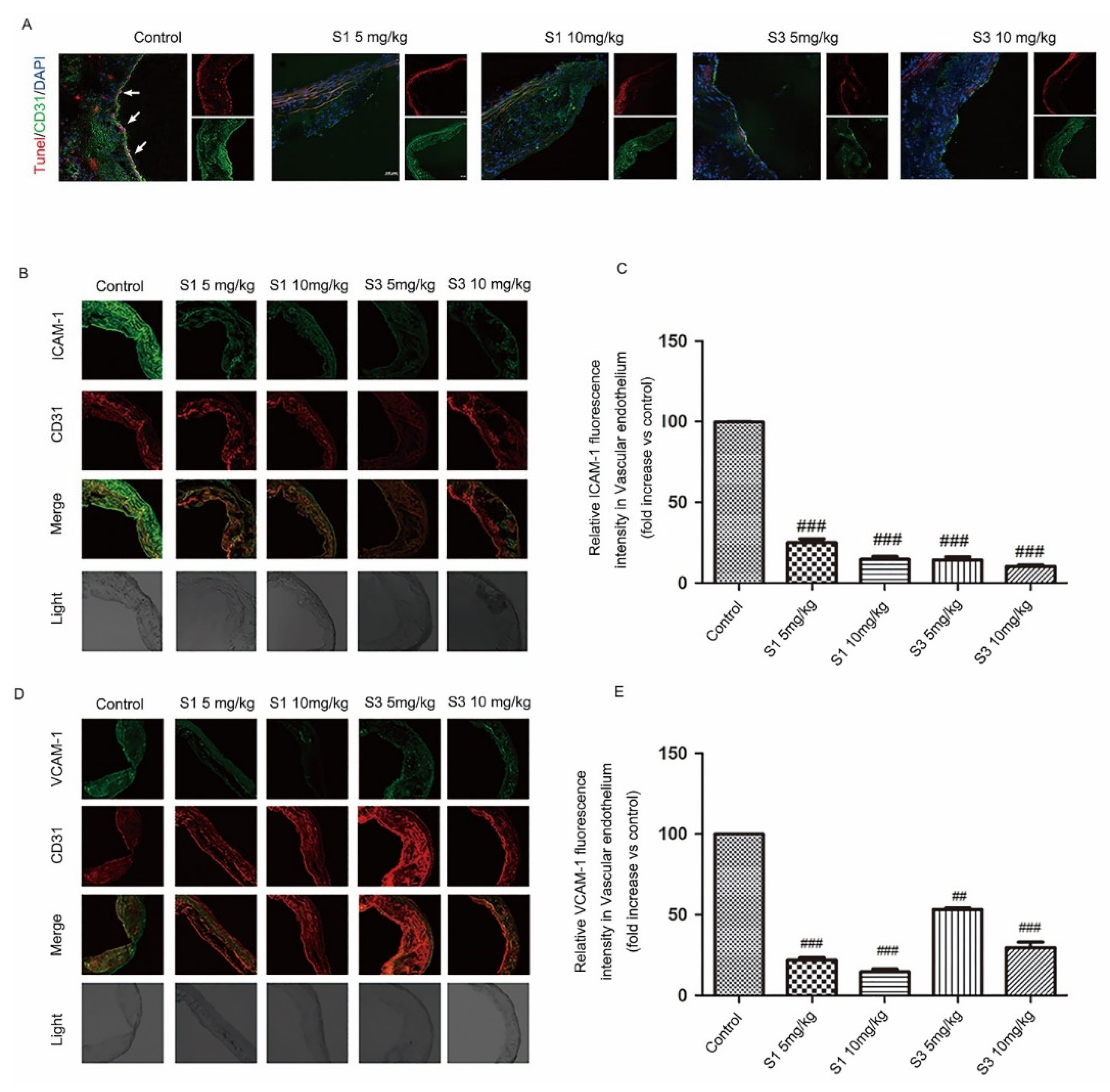
2.2. Dihydrohomoplantagin and Homoplantaginin Inhibited Plaque Endothelium Apoptosis and the Adhesion Factor Protein Level in apoE-/- Mice
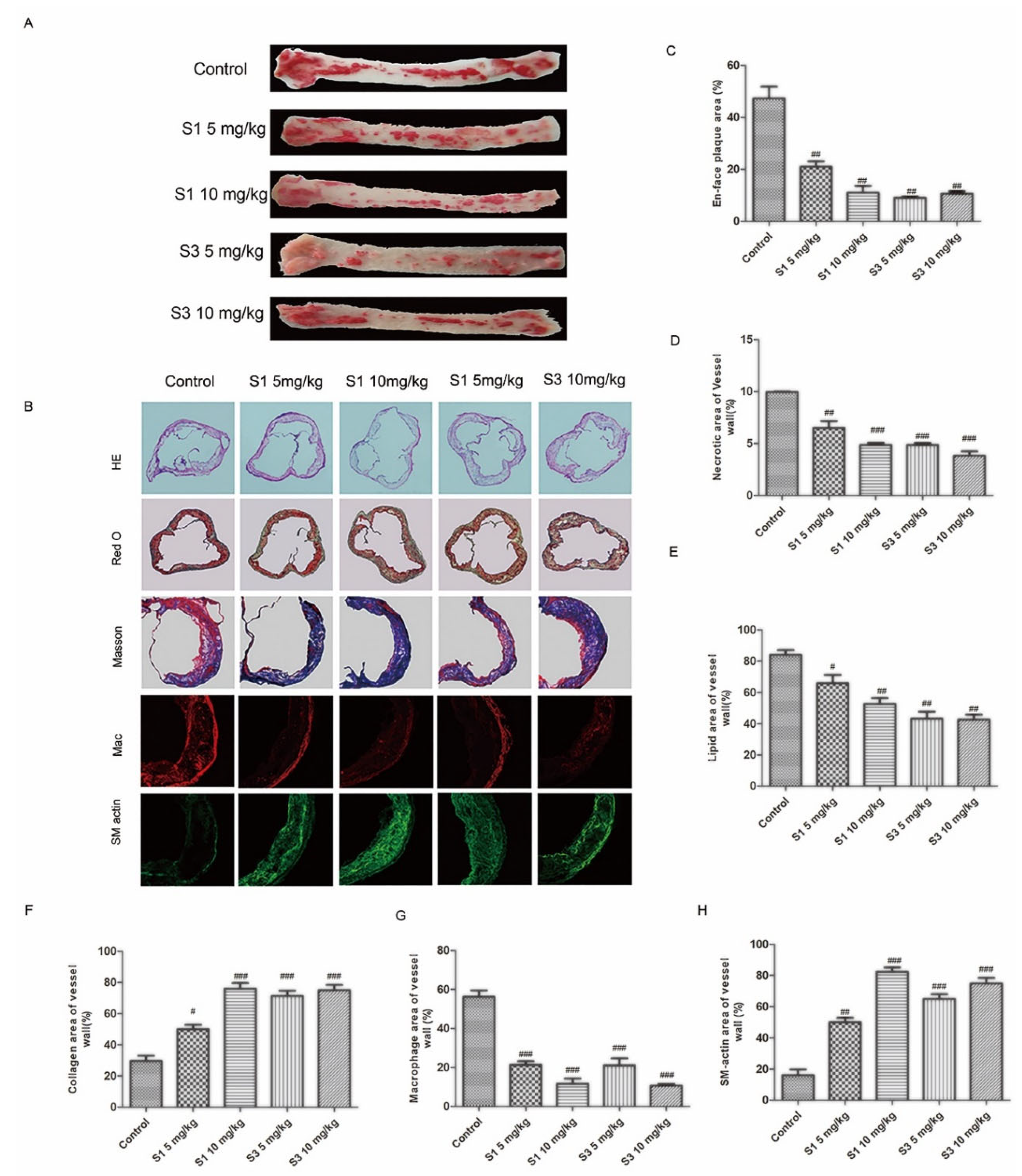
2.3. Dihydrohomoplantagin and Homoplantaginin Restricted Atherosclerosis Development in apoE-/- Mice
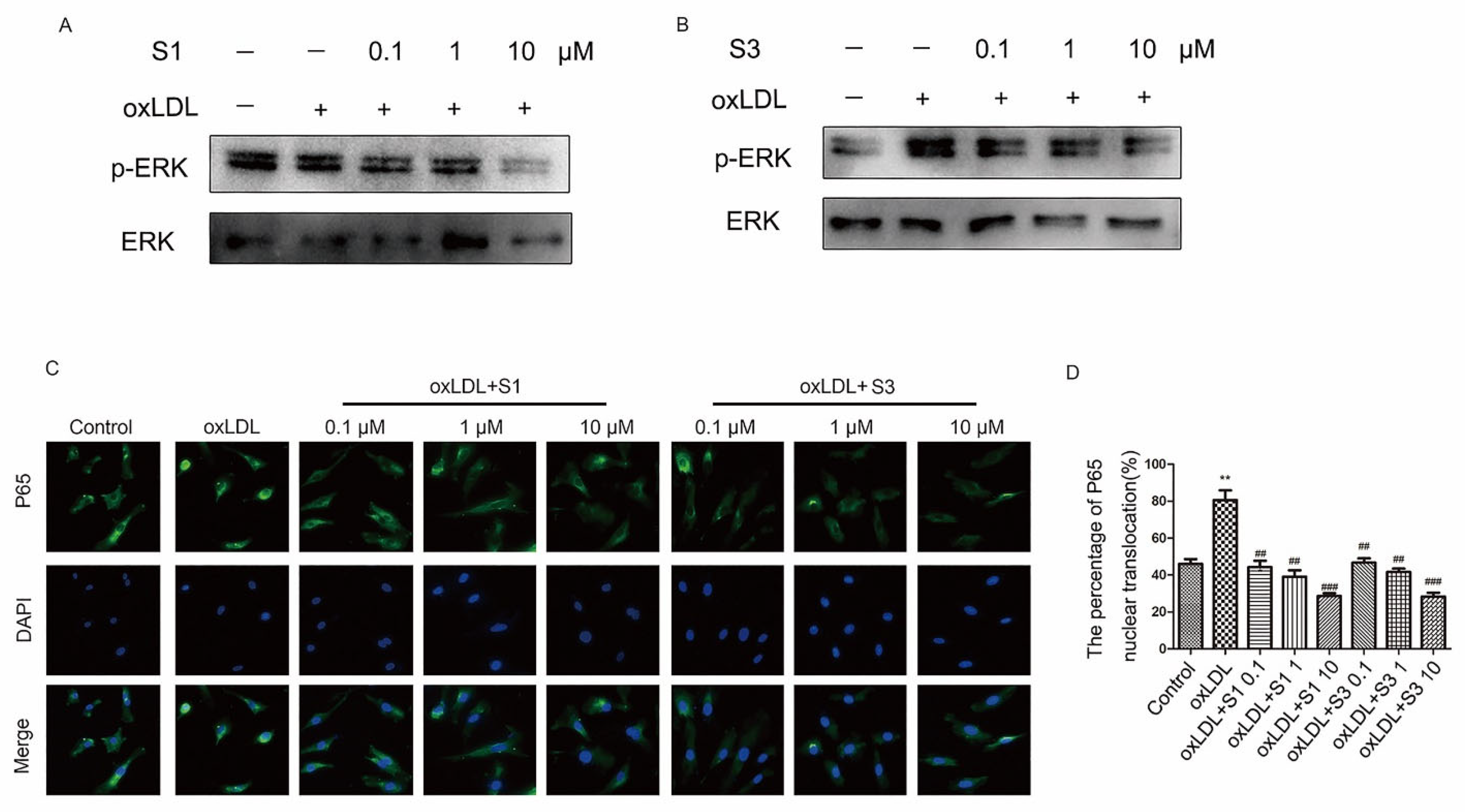
2.4. Dihydrohomoplantagin and Homoplantaginin Inhibited ROS/ERK/NF-ĸB Signaling Pathway
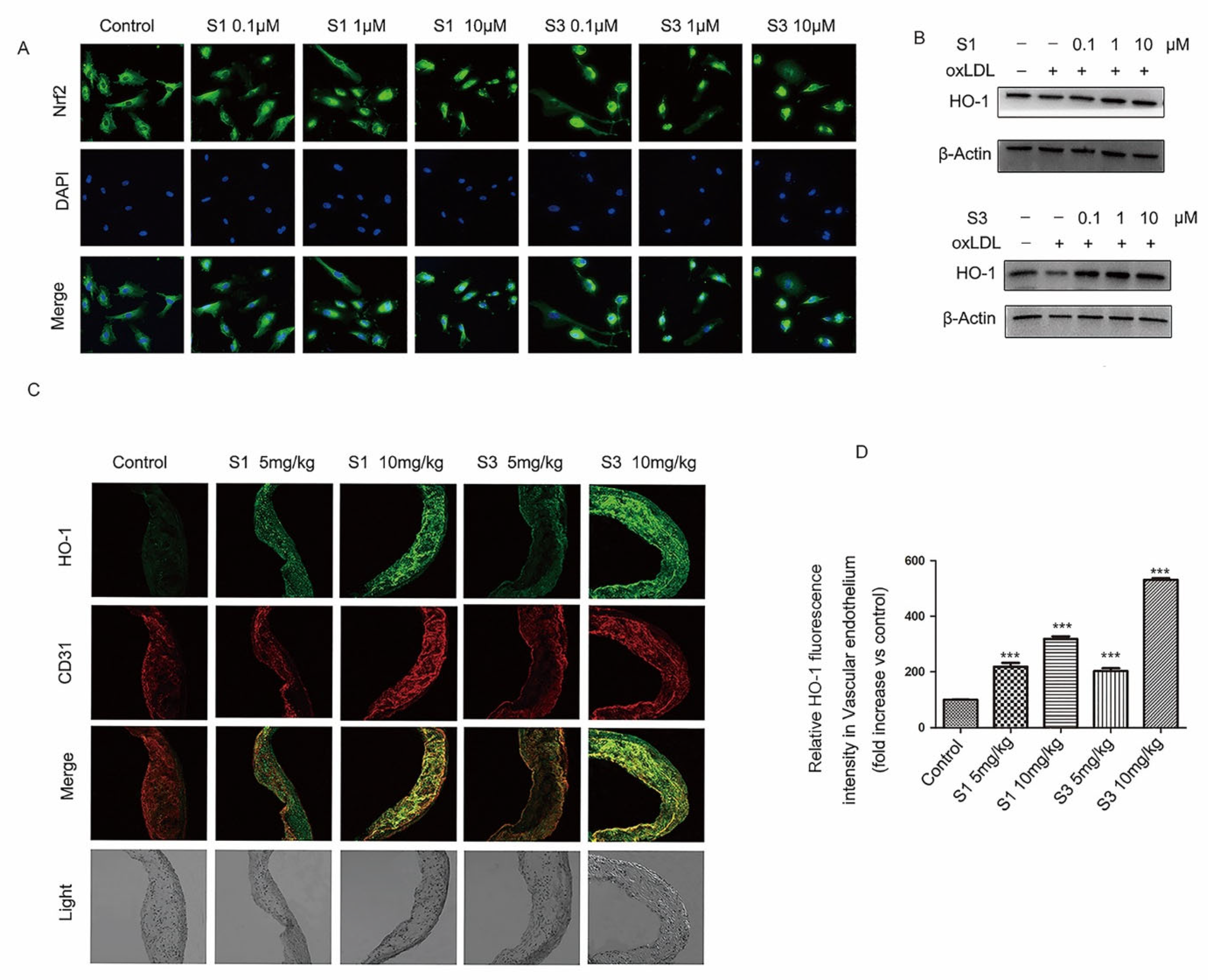
2.5. Dihydrohomoplantagin and Homoplantaginin Activated Nrf2 /HO-1 Anti-Oxidation Signal Pathway
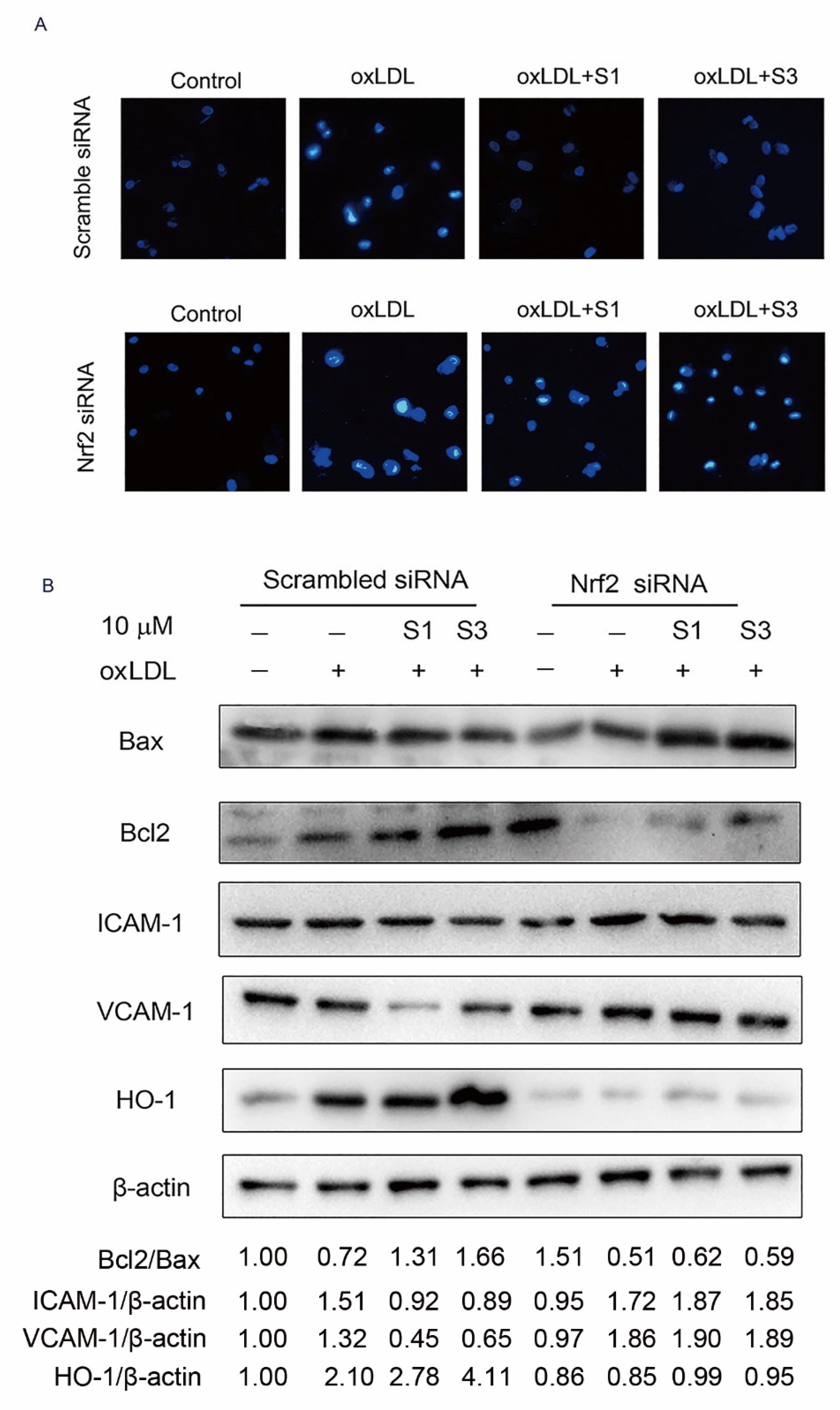
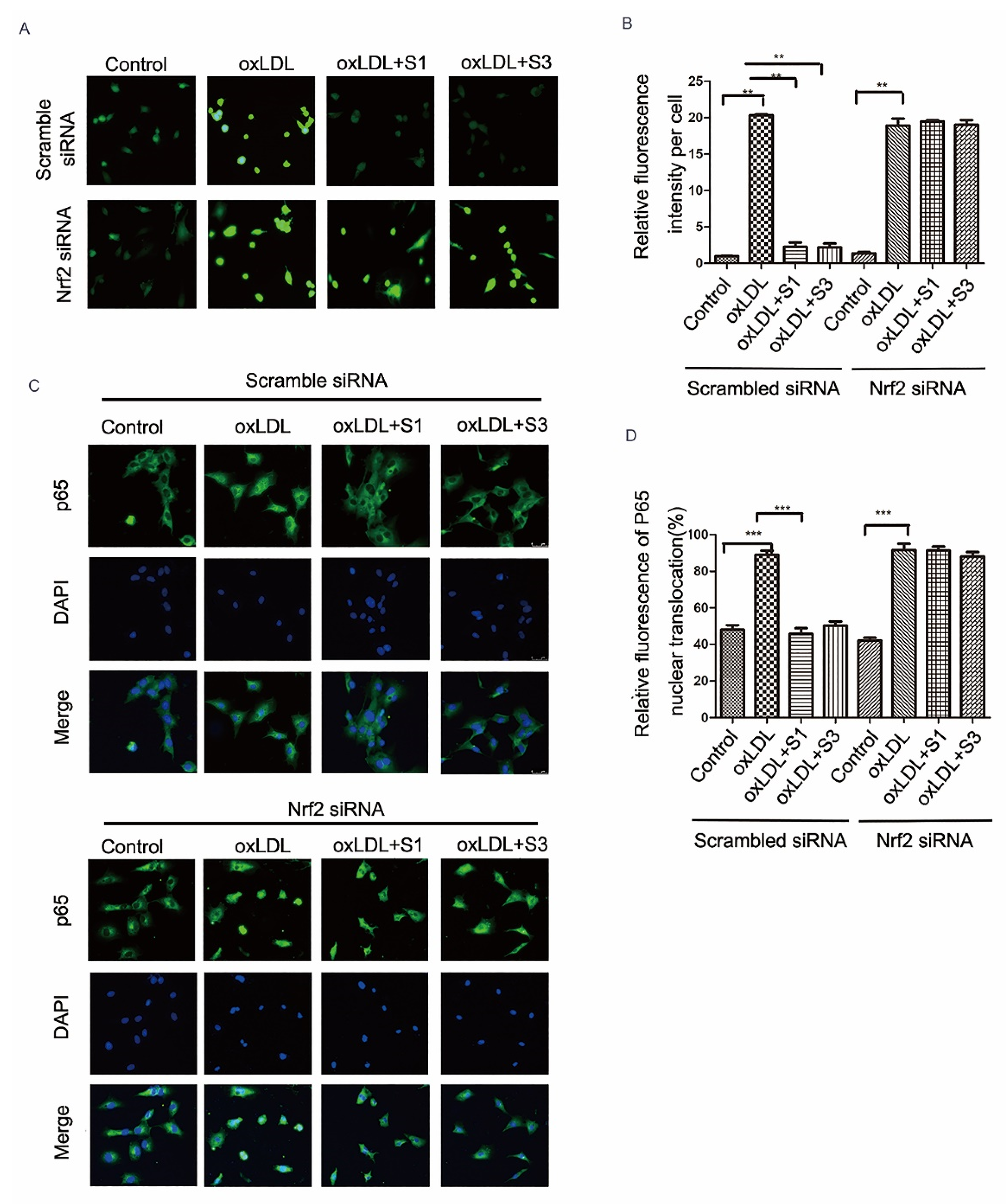
2.6. Dihydrohomoplantagin and Homoplantaginin Inhibition of oxLDL-Induced VEC Injury and ROS/NF-κB Activation Were Nrf2-Dependent
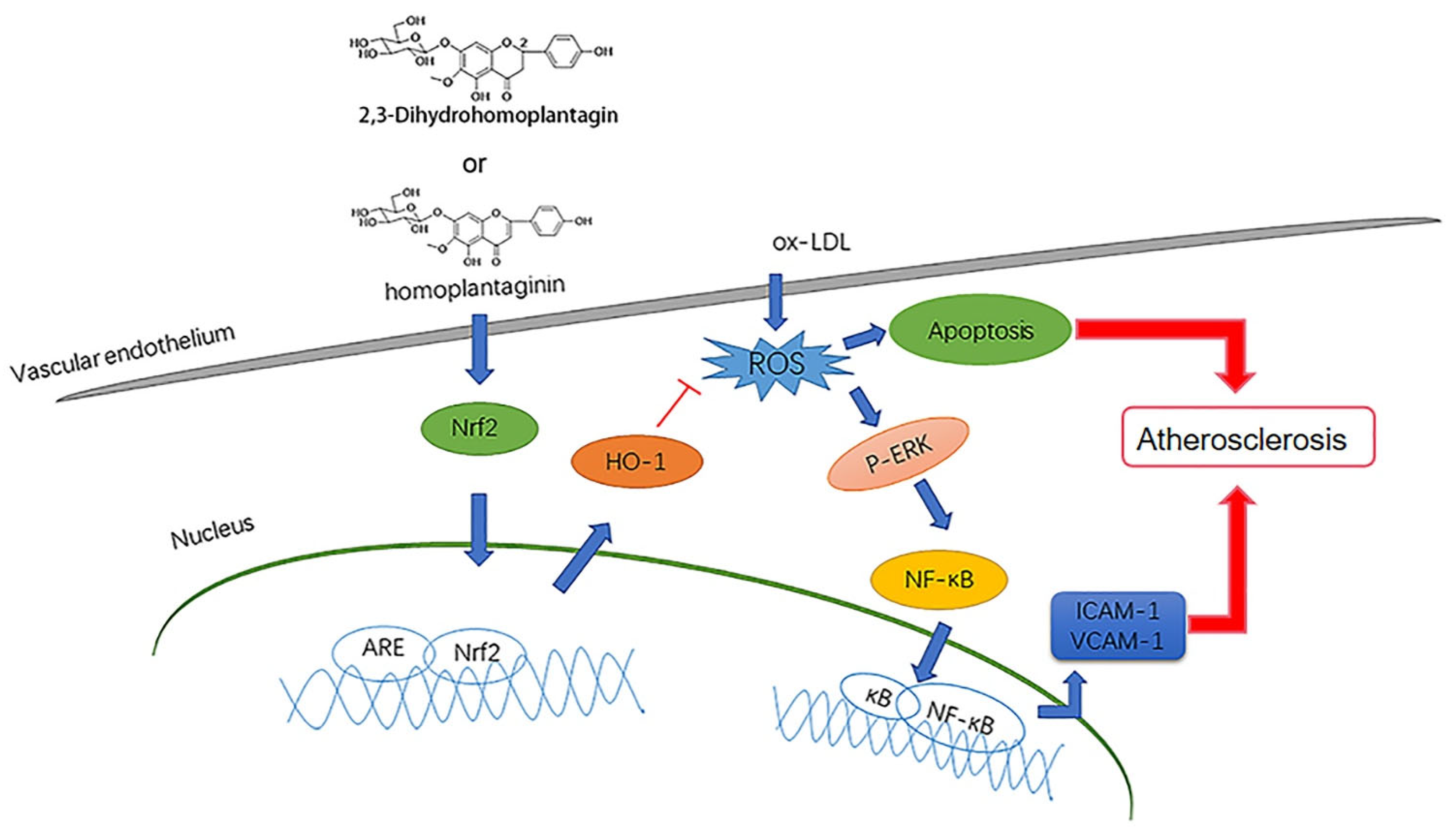
3. Discussion
4. Materials and Methods
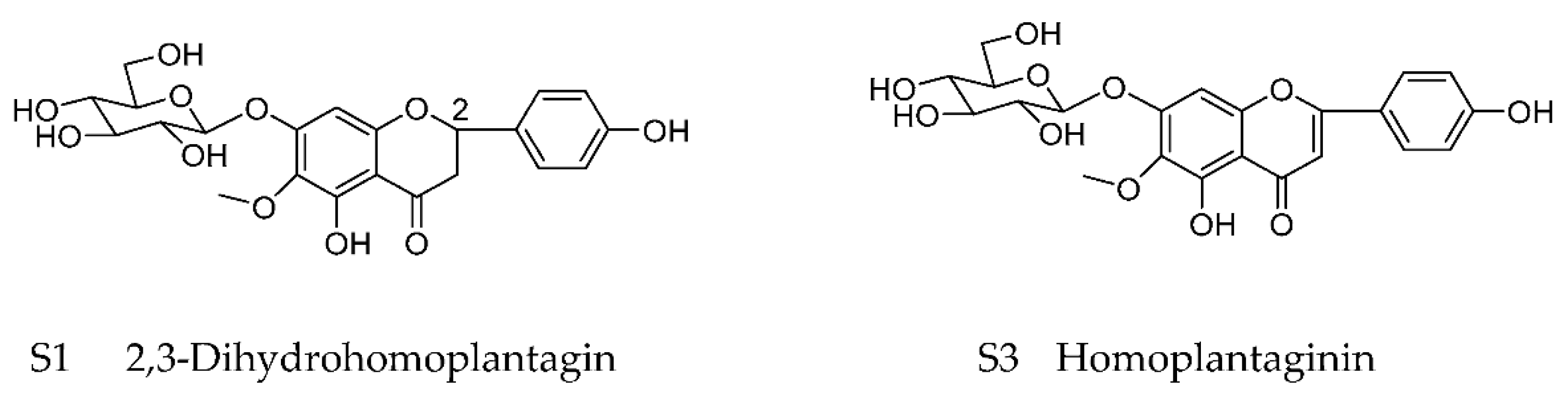
4.1. Preparation of Dihydrohomoplantagin and Homoplantaginin
4.2. Cell Culture and Treatment
4.3. Antibodies
4.4. Western Blot Analysis
4.5. Hoechst 33258 Staining
4.6. Intracellular ROS Assay
4.7. Cells for Immunofluorescence Staining
4.8. TUNEL Staining
4.9. Animal Model
4.10. Histological and Immunofluorescence
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Gimbrone, M.A., Jr.; García-Cardeña, G. Endothelial cell dysfunction and the pathobiology of atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suciu, C.F.; Prete, M.; Ruscitti, P.; Favoino, E.; Giacomelli, R.; Perosa, F. Oxidized low density lipoproteins: The bridge between atherosclerosis and autoimmunity. Possible implications in accelerated atherosclerosis and for immune intervention in autoimmune rheumatic disorders. Autoimmun. Rev. 2018, 17, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.; Jing, X.; Yang, Z.; Shi, Z.; Chen, R.; Xu, A.; Wang, N.; Jiang, J.; Yang, C.; Zhang, D.; et al. Downregulation of LncRNA NORAD promotes oxLDL-induced vascular endothelial cell injury and atherosclerosis. Aging (Albany NY) 2020, 12, 6385–6400. [Google Scholar] [CrossRef] [PubMed]
- Pirillo, A.; Norata, G.D.; Catapano, A.L. LOX-1, OxLDL, and atherosclerosis. Mediat. Inflamm. 2013, 2013, 152786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, P.; Jiang, F.; Cheng, J.; Ma, L.; Zhang, Y.; Zhao, Y. Traditional Chinese medicine for cardiovascular disease: Evidence and potential mechanisms. J. Am. Coll. Cardiol. 2017, 69, 2952–2966. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Niimi, M.; Watanabe, T.; Wang, Y.; Liang, J.; Fan, J. Treatment of atherosclerosis by traditional Chinese medicine: Questions and quandaries. Atherosclerosis 2018, 277, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.F.; Lu, Y.H.; Wei, D.Z.; Wang, Z.T. Chemical fingerprint and quantitative analysis of Salvia plebeia R. Br. by high-performance liquid chromatography. J. Pharm. Biomed. Anal. 2008, 48, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Y.; Wan, X.H.; Niu, F.J.; Xie, S.M.; Guo, H.; Yang, Y.Y.; Guo, L.-Y.; Zhou, C.-Z. Salvia plebeia R. Br.: An overview about its traditional uses, chemical constituents, pharmacology and modern applications. Biomed. Pharmacother. 2020, 121, 109589. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Xia, X.; Wang, Y.S.; Song, M.J.; Liu, L.L.; Xie, Y.Y.; Cheng, Y.N.; Liu, X.J.; Qiu, L.L.; Xiang, L.; et al. Protective effects of Salvia plebeia compound homoplantaginin on hepatocyte injury. Food Chem. Toxicol. 2009, 47, 1710–1715. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, J.L.; Kang, M.K.; Gong, J.H.; Han, S.Y.; Shim, J.H.; Lim, S.S.; Kang, Y.H. Sage weed (Salvia plebeia) extract antago-nizes foam cell formation and promotes cholesterol efflux in murine macrophages. Int. J. Mol. Med. 2012, 30, 1105–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Wang, H.; Li, J.; Liang, J.; Ma, S. Homoplantaginin modulates insulin sensitivity in endothelial cells by inhibiting in-flammation. Biol. Pharm. Bull. 2012, 35, 1171–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Zhang, B.; Wu, F.; Wang, L.; Shi, X.; Qin, W.; Lin, Y.; Ma, S.; Liang, J. Homoplantaginin inhibits palmitic acid-induced endothelial cells inflammation by suppressing TLR4 and NLRP3 inflammasome. J. Cardiovasc. Pharmacol. 2016, 67, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Xia, N.; Li, H. Roles of vascular oxidative stress and nitric oxide in the pathogenesis of atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Huang, C.S.; Lin, A.H.; Liu, C.T.; Tsai, C.W.; Chang, I.S.; Chen, H.W.; Lii, C.-K. Isothiocyanates protect against oxidized LDL-induced endothelial dysfunction by upregulating Nrf2-dependent antioxidation and suppressing NFκB activation. Mol. Nutr. Food Res. 2013, 57, 1918–1930. [Google Scholar] [CrossRef]
- Wang, R.; Wang, M.; Ye, J.; Sun, G.; Sun, X. Mechanism overview and target mining of atherosclerosis: Endothelial cell injury in atherosclerosis is regulated by glycolysis (Review). Int. J. Mol. Med. 2021, 47, 65–76. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Kanuri, S.H.; Mehta, J.L. Role of oxLDL and LOX-1 in atherogenesis. Curr. Med. Chem. 2019, 26, 1693–1700. [Google Scholar] [CrossRef]
- Barreca, D.; Gattuso, G.; Bellocco, E.; Calderaro, A.; Trombetta, D.; Smeriglio, A.; Laganà, G.; Daglia, M.; Meneghini, S.; Nabavi, S.M. Flavanones: Citrus phytochemical with health-promoting properties. Biofactors 2017, 43, 495–506. [Google Scholar] [CrossRef]
- Moharram, F.A.; Nagy, M.M.; El Dib, R.A.; El-Tantawy, M.M.; El Hossary, G.G.; El-Hosari, D.G. Pharmacological activity and fla-vonoids constituents of Artemisia judaica L aerial parts. J. Ethnopharmacol. 2021, 270, 113777. [Google Scholar] [CrossRef]
- Choi, H.J.; Eun, J.S.; Kim, B.G.; Kim, S.Y.; Jeon, H.; Soh, Y. Vitexin, an HIF-1alpha inhibitor, has anti-metastatic potential in PC12 cells. Mol. Cells 2006, 22, 291–299. [Google Scholar] [PubMed]
- Cominacini, L.; Garbin, U.; Pasini, A.F.; Davoli, A.; Campagnola, M.; Pastorino, A.M.; Gaviraghi, G.; Cascio, V.L. Oxidized low-density lipoprotein increases the production of intracellular reactive oxygen species in endothelial cells: Inhibitory effect of lacidipine. J. Hypertens. 1998, 16, 1913–1919. [Google Scholar] [CrossRef]
- Zheng, S.; Chen, Y.; Wang, Z.; Che, Y.; Wu, Q.; Yuan, S.; Zhong, X. Combination of matrine and tacrolimus alleviates acute re-jection in murine heart transplantation by inhibiting DCs maturation through ROS/ERK/NF-κB pathway. Int. Immunopharmacol. 2021, 101, 108218. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Qin, J.; Hao, Y.; Liu, M.; Luo, J.; Luo, T. Astragalus polysaccharide suppresses skeletal muscle myostatin expres-sion in diabetes: Involvement of ROS-ERK and NF-κB pathways. Oxid. Med. Cell. Longev. 2013, 2013, 782497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yuan, B.; Huang, H.; Qu, S.; Yang, S.; Zeng, Z. Gastrodin induced HO-1 and Nrf2 up-regulation to alleviate H2O2-induced oxidative stress in mouse liver sinusoidal endothelial cells through p38 MAPK phosphorylation. Braz. J. Med. Biol. Res. 2018, 51, e7439. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Feng, W.; Liu, J.; Jiang, L.; Chen, S.; Yuan, T.; Yu, C.; Xie, H.; Geng, D.; Qin, J. 7,8-Dihydroxyflavone activates Nrf2/HO-1 signaling pathways and protects against osteoarthritis. Exp. Ther. Med. 2019, 18, 1677–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Xu, J.; Cai, Y.; Yang, Y.; Liu, Y.; Cao, S. Cytoprotection against oxidative stress by methylnis-solin-3-O-β-d-glucopyranoside from Astragalus membranaceus mainly via the activation of the Nrf2/HO-1 pathway. Molecules 2021, 26, 3852. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S.; Sun, S.; Lu, Y.; Gao, K.; Guo, C.; Li, R.; Li, W.; Zhao, X.; Tang, H. Apig-enin-7-O-β-d-(-6″-p-coumaroyl)-glucopyranoside treatment elicits a neuroprotective effect through GSK-3β phosphory-lation-mediated Nrf2 activation. Aging (Albany NY) 2020, 12, 23872–23888. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.T.; Duan, C.H.; Lee, J.N.; Lee, K.S.; Hong, J.T.; Lee, K.K. Phytochemical constituents from Salvia plebeia. Nat. Prod. Sci. 2010, 16, 207–210. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, N.; Chen, K.; Wang, Y.; Hou, J.; Chu, W.; Xie, S.; Yang, F.; Sun, C. Dihydrohomoplantagin and Homoplantaginin, Major Flavonoid Glycosides from Salvia plebeia R. Br. Inhibit oxLDL-Induced Endothelial Cell Injury and Restrict Atherosclerosis via Activating Nrf2 Anti-Oxidation Signal Pathway. Molecules 2022, 27, 1990. https://doi.org/10.3390/molecules27061990
Meng N, Chen K, Wang Y, Hou J, Chu W, Xie S, Yang F, Sun C. Dihydrohomoplantagin and Homoplantaginin, Major Flavonoid Glycosides from Salvia plebeia R. Br. Inhibit oxLDL-Induced Endothelial Cell Injury and Restrict Atherosclerosis via Activating Nrf2 Anti-Oxidation Signal Pathway. Molecules. 2022; 27(6):1990. https://doi.org/10.3390/molecules27061990
Chicago/Turabian StyleMeng, Ning, Kai Chen, Yanhong Wang, Jiarong Hou, Wenhui Chu, Shan Xie, Fengying Yang, and Chunhui Sun. 2022. "Dihydrohomoplantagin and Homoplantaginin, Major Flavonoid Glycosides from Salvia plebeia R. Br. Inhibit oxLDL-Induced Endothelial Cell Injury and Restrict Atherosclerosis via Activating Nrf2 Anti-Oxidation Signal Pathway" Molecules 27, no. 6: 1990. https://doi.org/10.3390/molecules27061990
APA StyleMeng, N., Chen, K., Wang, Y., Hou, J., Chu, W., Xie, S., Yang, F., & Sun, C. (2022). Dihydrohomoplantagin and Homoplantaginin, Major Flavonoid Glycosides from Salvia plebeia R. Br. Inhibit oxLDL-Induced Endothelial Cell Injury and Restrict Atherosclerosis via Activating Nrf2 Anti-Oxidation Signal Pathway. Molecules, 27(6), 1990. https://doi.org/10.3390/molecules27061990