
Resolvin D5 (RvD5) Reduces Renal Damage Caused by LPS Endotoxemia in Female Mice
 , ,
, ,  , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
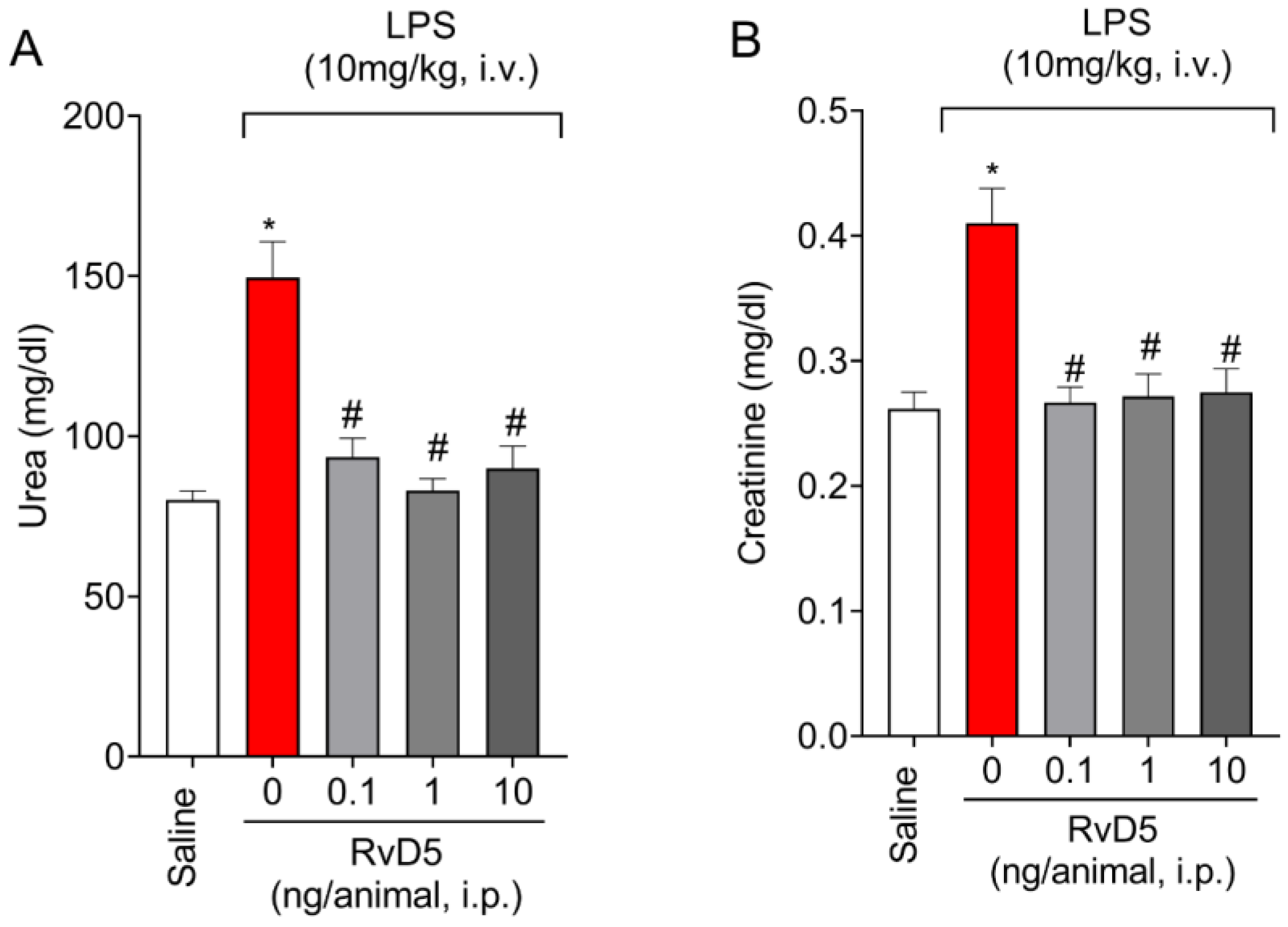
2.1. RvD5 Decreases the Levels of Renal Injury Markers in Plasma
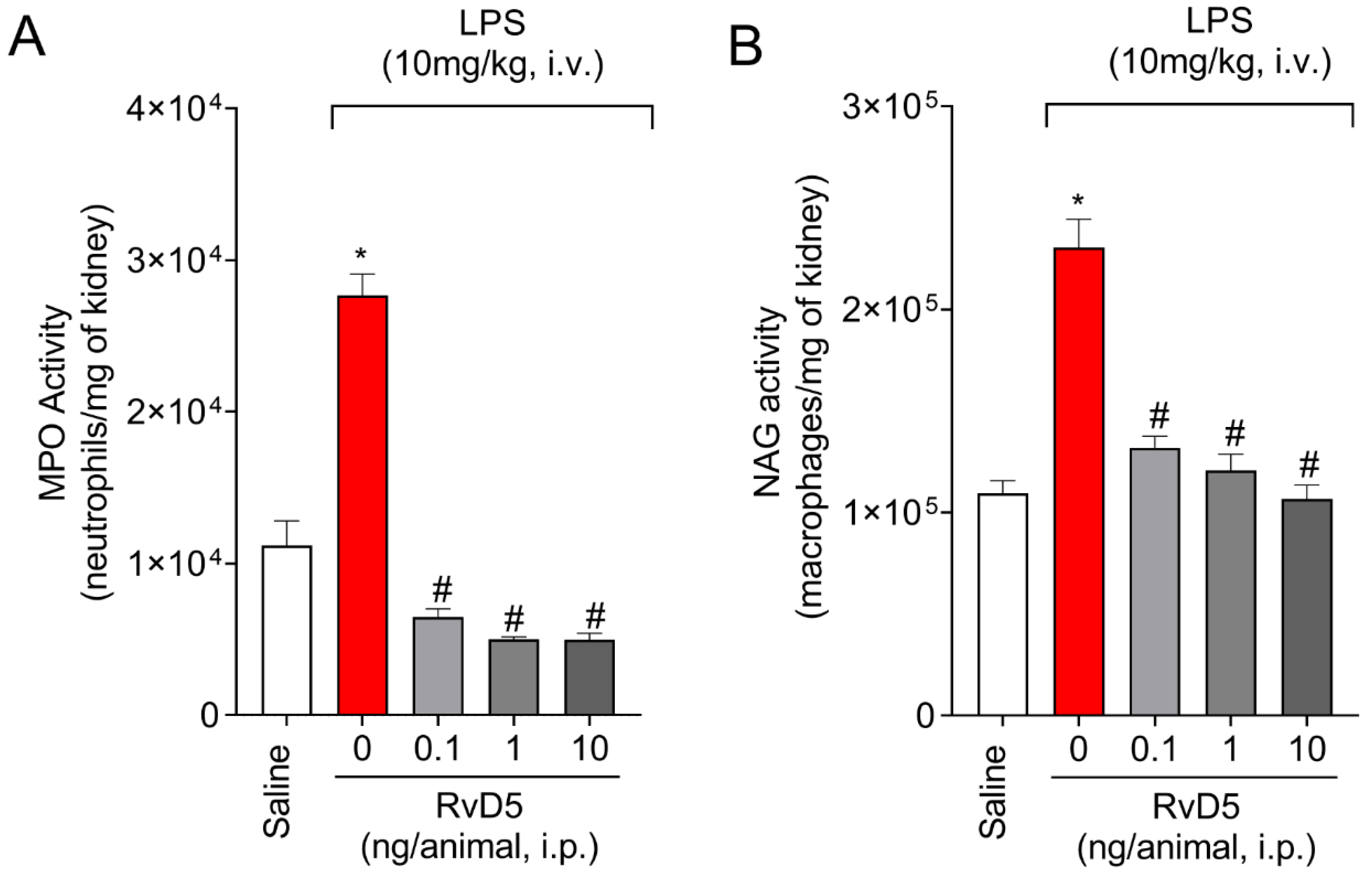
2.2. RvD5 Decreases MPO and NAG Activities in the Kidneys
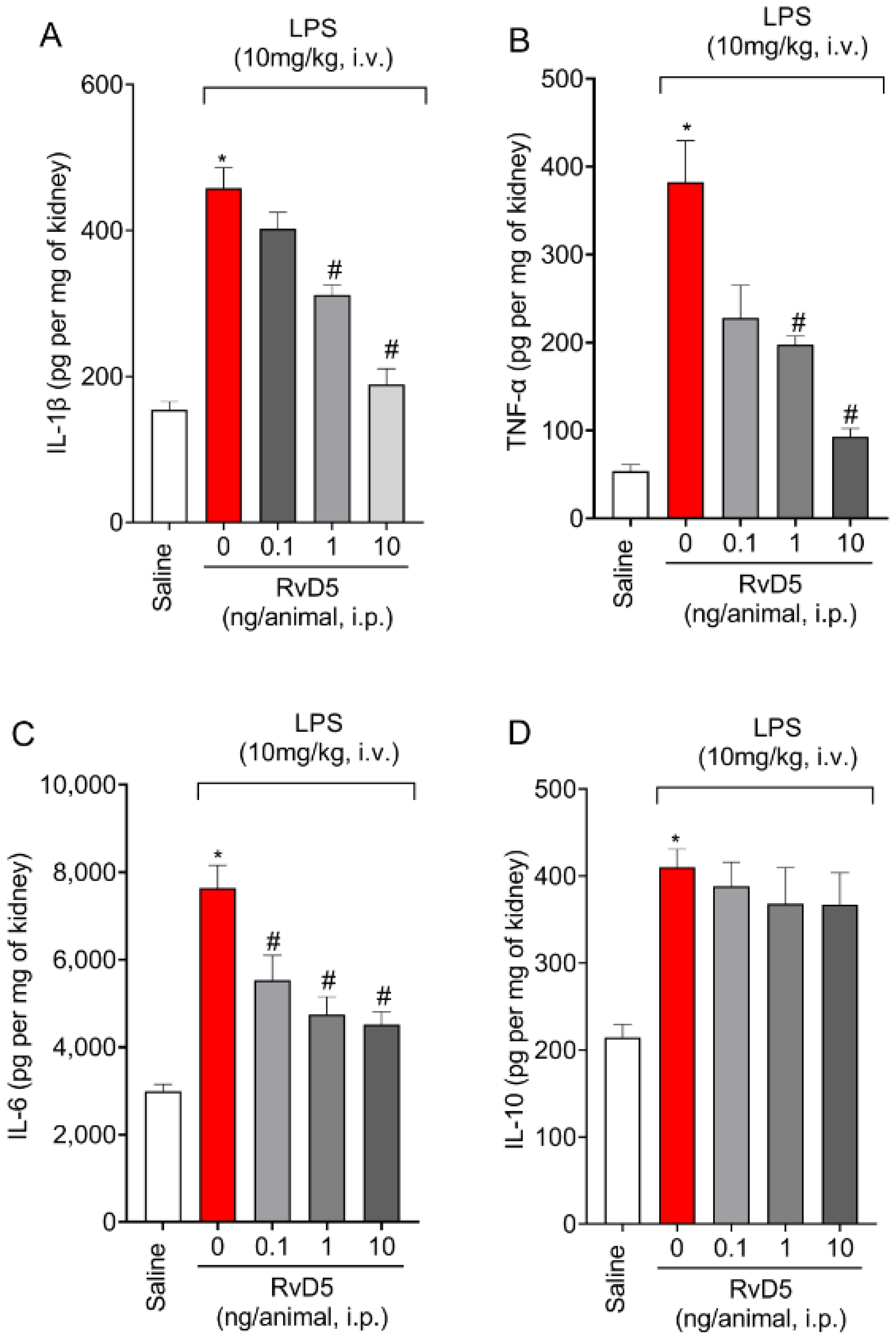
2.3. RvD5 Decreases the Production of Pro-Inflammatory Cytokines but Does Not Affect Anti-Inflammatory Cytokine Levels in The Kidneys
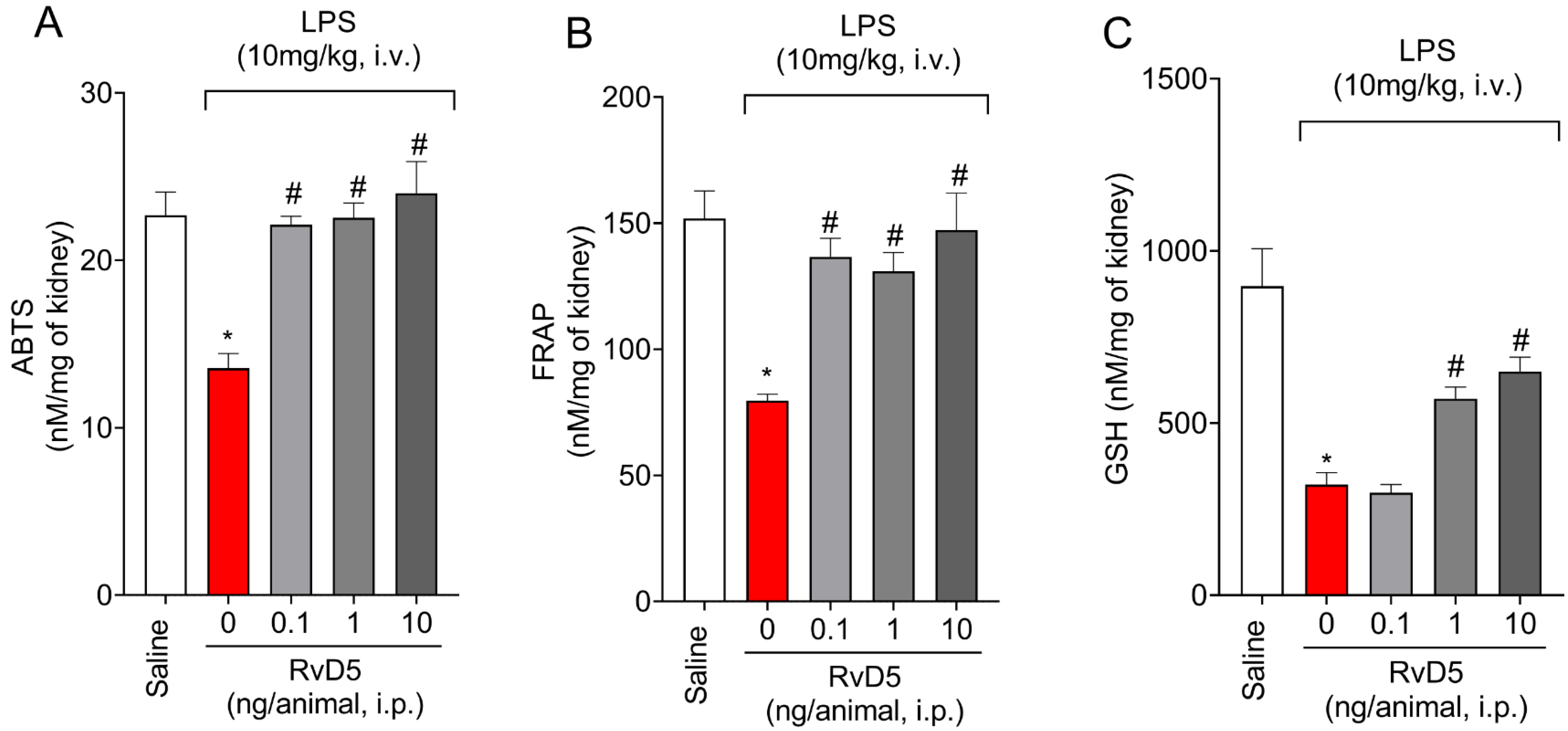
2.4. RvD5 Reestablishes Antioxidant Capacity
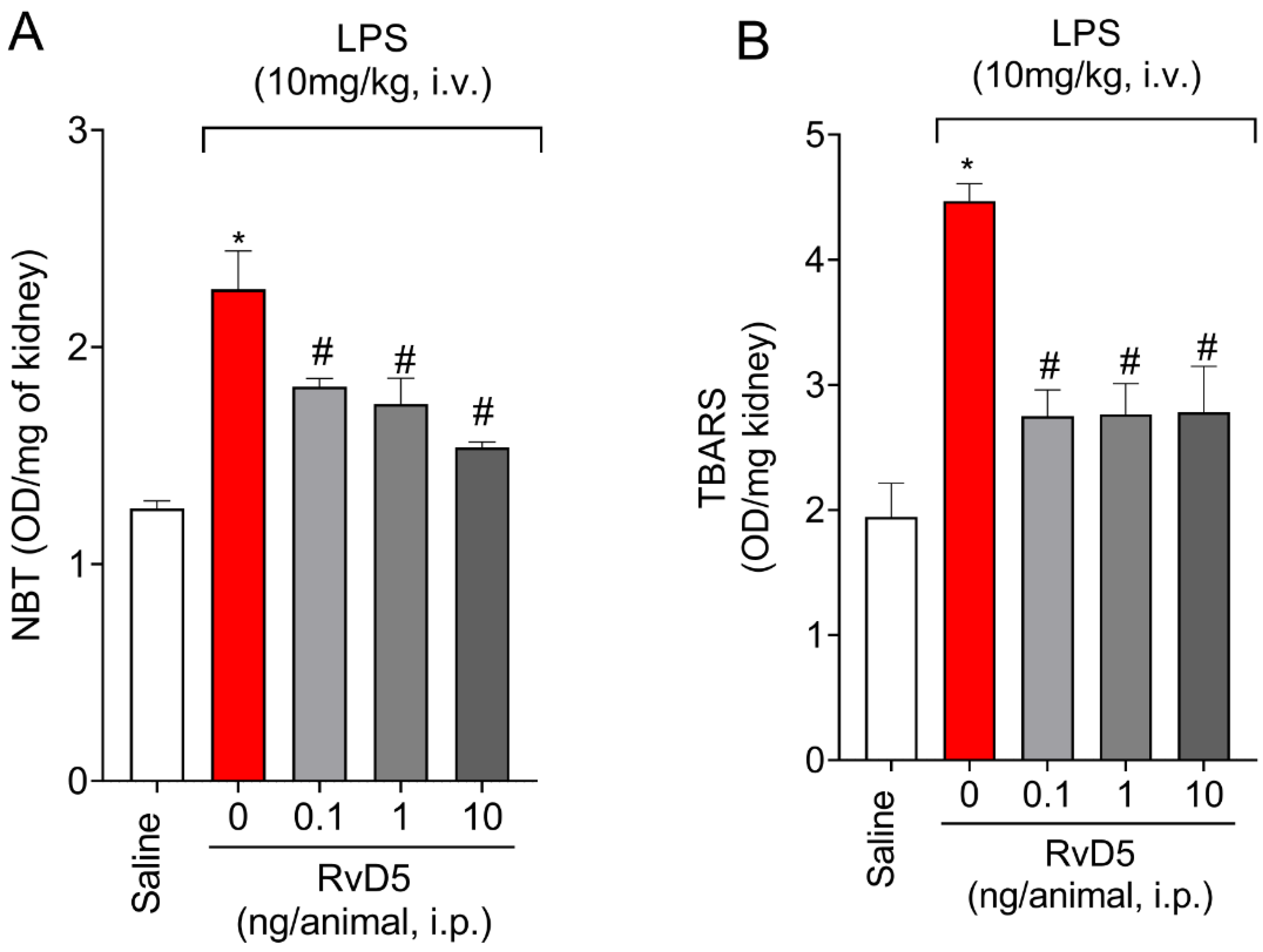
2.5. RvD5 Decreases Oxidative Stress
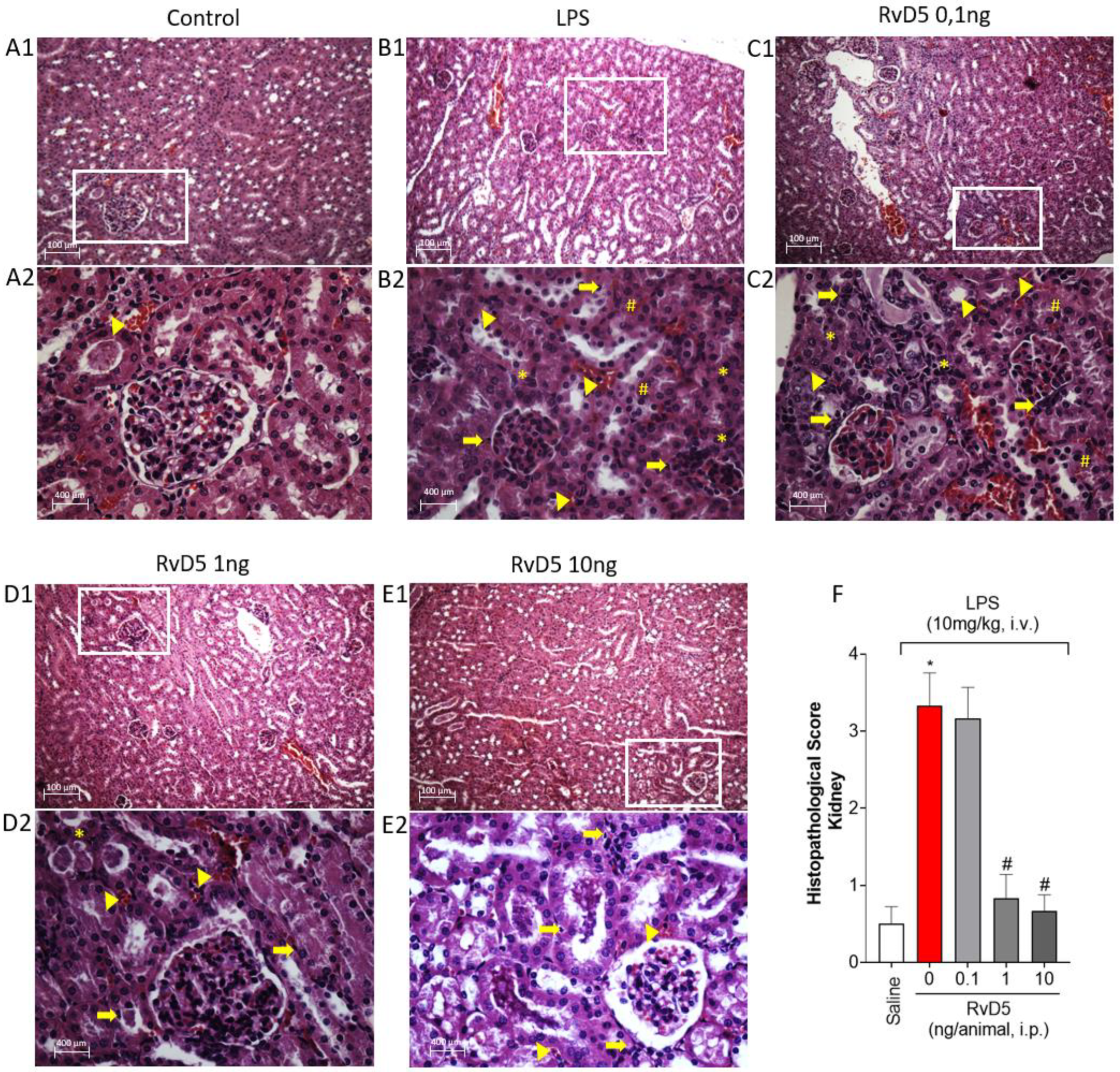
2.6. RvD5 Decreases Histopathological Changes, Leukocyte Infiltration, and Necrosis in the Kidneys
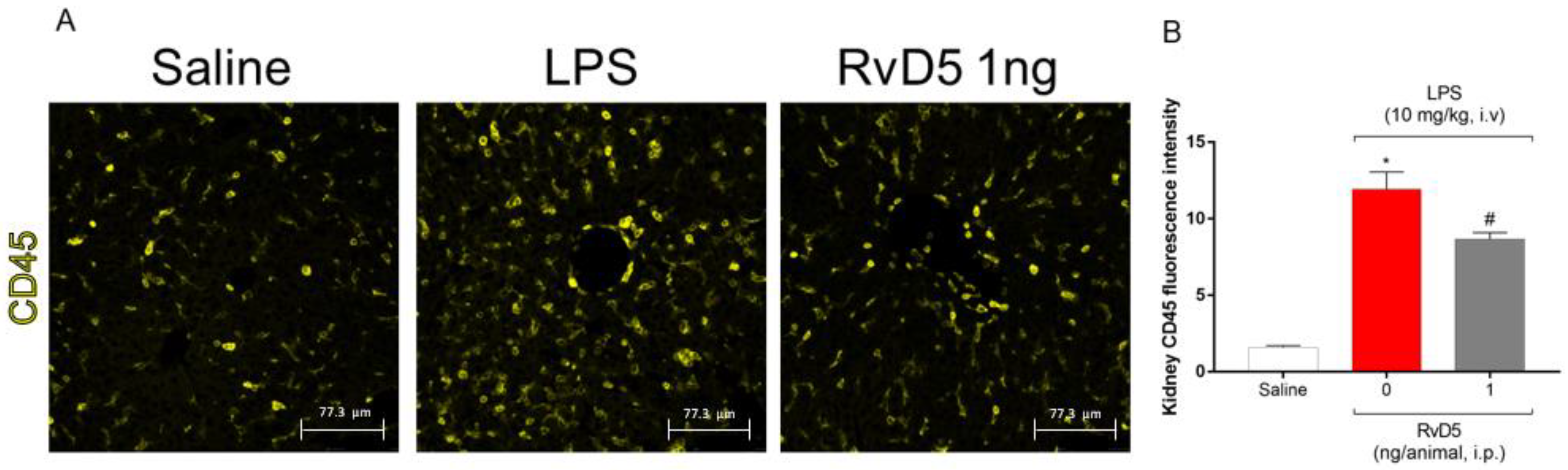
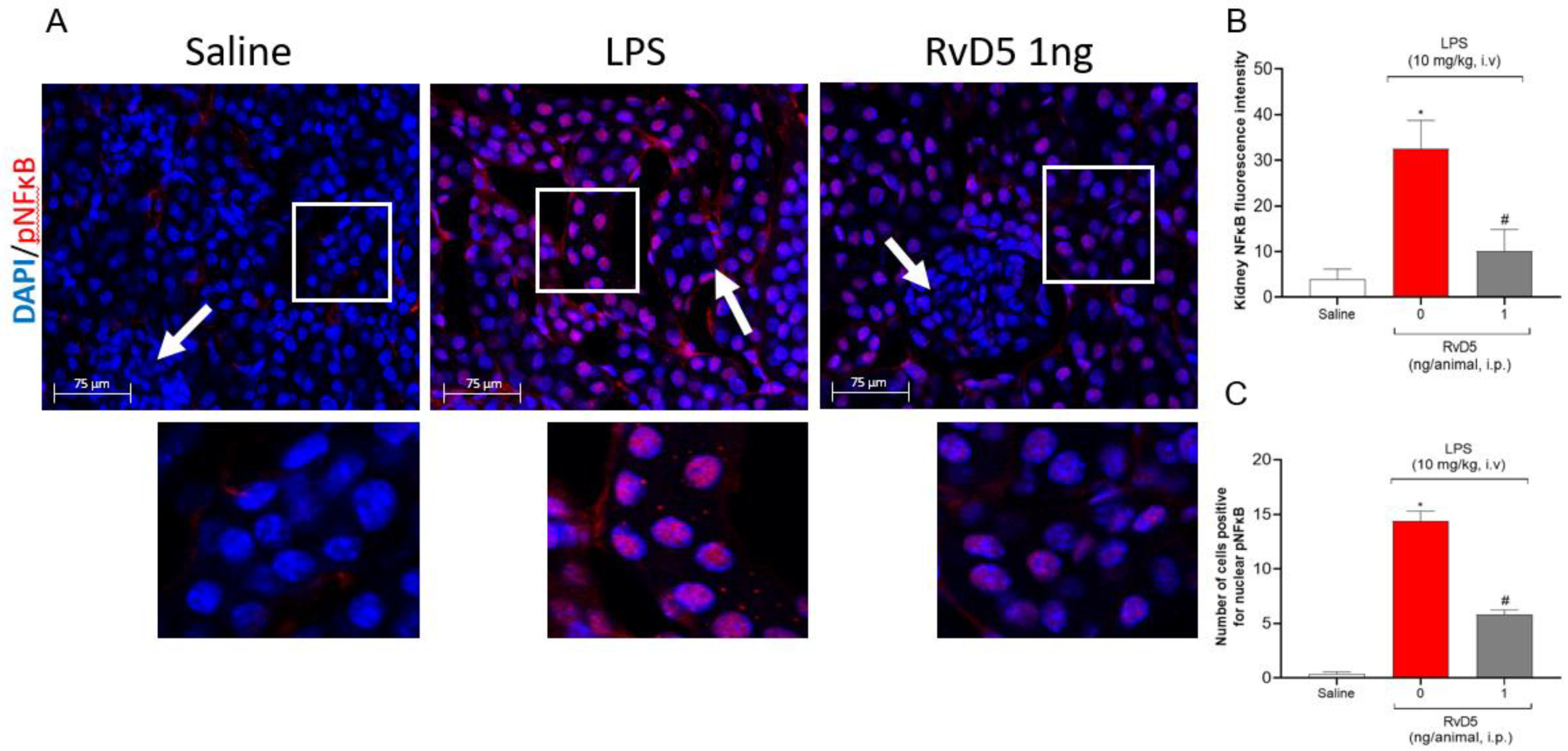
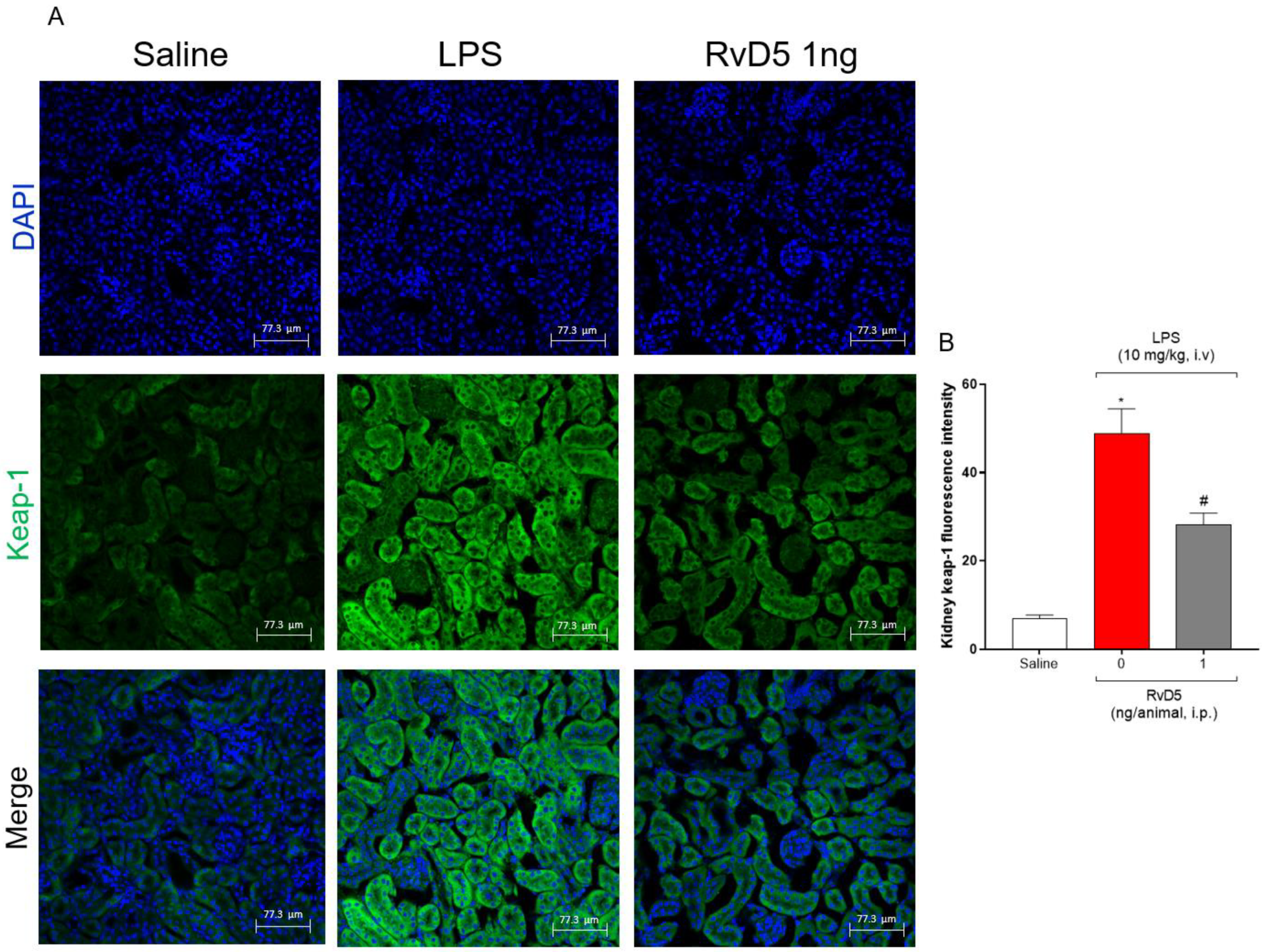
2.7. RvD5 Reduces the Infiltration of CD45+ Hematopoietic Leukocytes, Pnfkb and Keap-1 Staining in the Kidneys of Endotoxemic Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Procedures and Endotoxemia Model
4.3. Determination of Renal Injury Markers Levels in the Plasma
4.4. Myeloperoxidase (MPO) Activity Assay
4.5. N-acetylglucosaminidase (NAG) Activity Assay
4.6. Cytokine Measurement
4.7. ABTS and FRAP Assays
4.8. Determination of Thiobarbituric Acid Reactive Substances (TBARS) Levels
4.9. Nitroblue Tetrazolium (NBT) Assay
4.10. Determination of Reduced Glutathione (GSH) Level
4.11. Histopathological Analysis
4.12. Immunofluorescence Staining
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Font, M.D.; Thyagarajan, B.; Khanna, A.K. Sepsis and Septic Shock—Basics of diagnosis, pathophysiology and clinical decision making. Med. Clin. N. Am. 2020, 104, 573–585. [Google Scholar] [CrossRef]
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A. Sepsis and septic shock. Lancet 2018, 392, 75–87. [Google Scholar] [CrossRef]
- Bauer, M.; Gerlach, H.; Vogelmann, T.; Preissing, F.; Stiefel, J.; Adam, D. Mortality in sepsis and septic shock in Europe, North America and Australia between 2009 and 2019- results from a systematic review and meta-analysis. Crit. Care 2020, 24, 239. [Google Scholar] [CrossRef]
- Caraballo, C.; Jaimes, F. Organ Dysfunction in Sepsis: An Ominous Trajectory From Infection To Death. Yale J. Biol. Med. 2019, 92, 629–640. [Google Scholar]
- Pool, R.; Gomez, H.; Kellum, J.A. Mechanisms of Organ Dysfunction in Sepsis. Crit. Care Clin. 2018, 34, 63–80. [Google Scholar] [CrossRef]
- Peerapornratana, S.; Manrique-Caballero, C.L.; Gomez, H.; Kellum, J.A. Acute kidney injury from sepsis: Current concepts, epidemiology, pathophysiology, prevention and treatment. Kidney Int. 2019, 96, 1083–1099. [Google Scholar] [CrossRef]
- Poston, J.T.; Koyner, J.L. Sepsis associated acute kidney injury. BMJ 2019, 364, k4891. [Google Scholar] [CrossRef]
- Skube, S.J.; Katz, S.A.; Chipman, J.G.; Tignanelli, C.J. Acute Kidney Injury and Sepsis. Surg. Infect. 2018, 19, 216–224. [Google Scholar] [CrossRef]
- Hoste, E.A.J.; Kellum, J.A.; Selby, N.M.; Zarbock, A.; Palevsky, P.M.; Bagshaw, S.M.; Goldstein, S.L.; Cerda, J.; Chawla, L.S. Global epidemiology and outcomes of acute kidney injury. Nat. Rev. Nephrol. 2018, 14, 607–625. [Google Scholar] [CrossRef]
- Dickson, K.; Lehmann, C. Inflammatory Response to Different Toxins in Experimental Sepsis Models. Int. J. Mol. Sci. 2019, 20, 4341. [Google Scholar] [CrossRef] [Green Version]
- Proniewski, B.; Kij, A.; Sitek, B.; Kelley, E.E.; Chlopicki, S. Multiorgan Development of Oxidative and Nitrosative Stress in LPS-Induced Endotoxemia in C57Bl/6 Mice: DHE-Based In Vivo Approach. Oxidative Med. Cell. Longev. 2019, 2019, 7838406. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Z.; Wang, L.; Jiang, L.; Qin, Z.; Zhao, Y.; Su, B. Maresin 1 Attenuates Lipopolysaccharide-Induced Acute Kidney Injury via Inhibiting NOX4/ROS/NF-kappaB Pathway. Front. Pharmacol. 2021, 12, 782660. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Zhang, L.; Yang, Y.Y.; Tang, Y.; Zhou, J.J.; Feng, Y.Y.; Cui, T.L.; Liu, F.; Fu, P. Resolvin D1 Protects Lipopolysaccharide-induced Acute Kidney Injury by Down-regulating Nuclear Factor-kappa B Signal and Inhibiting Apoptosis. Chin. Med. J. 2016, 129, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Molaei, E.; Molaei, A.; Hayes, A.W.; Karimi, G. Resolvin D1, therapeutic target in acute respiratory distress syndrome. Eur. J. Pharmacol. 2021, 911, 174527. [Google Scholar] [CrossRef] [PubMed]
- Dartt, D.A.; Hodges, R.R.; Serhan, C.N. Immunoresolvent Resolvin D1 Maintains the Health of the Ocular Surface. Adv. Exp. Med. Biol. 2019, 1161, 13–25. [Google Scholar] [CrossRef]
- Demarquoy, J.; Borgne, F.L. Biosynthesis, metabolism, and function of protectins and resolvins. Clin. Lipidol. 2014, 9, 683–693. [Google Scholar] [CrossRef]
- Chiang, N.; Fredman, G.; Backhed, F.; Oh, S.F.; Vickery, T.; Schmidt, B.A.; Serhan, C.N. Infection regulates pro-resolving mediators that lower antibiotic requirements. Nature 2012, 484, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Diaz, L.A.; Altman, N.H.; Khan, W.N.; Serhan, C.N.; Adkins, B. Specialized Proresolving Mediators Rescue Infant Mice from Lethal Citrobacter rodentium Infection and Promote Immunity against Reinfection. Infect. Immun. 2017, 85, e00464.17. [Google Scholar] [CrossRef] [Green Version]
- Leao, F.F.; Waltrick, A.P.F.; Verri, W.A., Jr.; da Cunha, J.M.; Zanoveli, J.M. Resolvin D5 disrupts anxious- and depressive-like behaviors in a type 1 diabetes mellitus animal model. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 1269–1282. [Google Scholar] [CrossRef]
- Yamada, H.; Saegusa, J.; Sendo, S.; Ueda, Y.; Okano, T.; Shinohara, M.; Morinobu, A. Effect of resolvin D5 on T cell differentiation and osteoclastogenesis analyzed by lipid mediator profiling in the experimental arthritis. Sci. Rep. 2021, 11, 17312. [Google Scholar] [CrossRef]
- Luo, X.; Gu, Y.; Tao, X.; Serhan, C.N.; Ji, R.R. Resolvin D5 Inhibits Neuropathic and Inflammatory Pain in Male But Not Female Mice: Distinct Actions of D-Series Resolvins in Chemotherapy-Induced Peripheral Neuropathy. Front. Pharmacol. 2019, 10, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutz, S.; Ouyang, W. Regulation of Interleukin-10 Expression. Adv. Exp. Med. Biol. 2016, 941, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Tallon, J.; Browning, B.; Couenne, F.; Bordes, C.; Venet, F.; Nony, P.; Gueyffier, F.; Moucadel, V.; Monneret, G.; Tayakout-Fayolle, M. Dynamical modeling of pro- and anti-inflammatory cytokines in the early stage of septic shock. In Silico Biol. 2020, 14, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Lauw, F.N.; Pajkrt, D.; Hack, C.E.; Kurimoto, M.; van Deventer, S.J.; van der Poll, T. Proinflammatory effects of IL-10 during human endotoxemia. J. Immunol. 2000, 165, 2783–2789. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1-Nrf2-ARE Pathway As a Potential Preventive and Therapeutic Target: An Update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Hurley, J.C.; Nowak, P.; Ohrmalm, L.; Gogos, C.; Armaganidis, A.; Giamarellos-Bourboulis, E.J. Endotoxemia as a diagnostic tool for patients with suspected bacteremia caused by gram-negative organisms: A meta-analysis of 4 decades of studies. J. Clin. Microbiol. 2015, 53, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Copeland, S.; Warren, H.S.; Lowry, S.F.; Calvano, S.E.; Remick, D. Acute inflammatory response to endotoxin in mice and humans. Clin. Diagn. Lab Immunol. 2005, 12, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, J.; Kisdi, E.; Anttila, J.V. Model of bacterial toxin-dependent pathogenesis explains infective dose. Proc. Natl. Acad. Sci. USA 2018, 115, 10690–10695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boffa, J.J.; Just, A.; Coffman, T.M.; Arendshorst, W.J. Thromboxane receptor mediates renal vasoconstriction and contributes to acute renal failure in endotoxemic mice. J. Am. Soc. Nephrol. 2004, 15, 2358–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, S.; Wang, W.; Falk, S.; Schrier, R. Combination therapy with albumin and pentoxifylline protects against acute kidney injury during endotoxemic shock in mice. Ren. Fail. 2009, 31, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.K.; Hsu, C.Y. The Role of Acute Kidney Injury in Chronic Kidney Disease. Semin. Nephrol. 2016, 36, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Hung, Y.L.; Fang, S.H.; Wang, S.C.; Cheng, W.C.; Liu, P.L.; Su, C.C.; Chen, C.S.; Huang, M.Y.; Hua, K.F.; Shen, K.H.; et al. Corylin protects LPS-induced sepsis and attenuates LPS-induced inflammatory response. Sci. Rep. 2017, 7, 46299. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zeng, M.; Li, B.; Kan, Y.; Zhang, B.; Zheng, X.; Feng, W. Raw and salt-processed Achyranthes bidentata attenuate LPS-induced acute kidney injury by inhibiting ROS and apoptosis via an estrogen-like pathway. Biomed. Pharmacother. 2020, 129, 110403. [Google Scholar] [CrossRef]
- Ren, Q.; Guo, F.; Tao, S.; Huang, R.; Ma, L.; Fu, P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-kappaB p65 and MAPK signaling pathways in septic AKI mice. Biomed. Pharmacother. 2020, 122, 109772. [Google Scholar] [CrossRef]
- Gowda, S.; Desai, P.B.; Kulkarni, S.S.; Hull, V.V.; Math, A.A.; Vernekar, S.N. Markers of renal function tests. N. Am. J. Med. Sci. 2010, 2, 170–173. [Google Scholar]
- van Veldhuisen, D.J.; Ruilope, L.M.; Maisel, A.S.; Damman, K. Biomarkers of renal injury and function: Diagnostic, prognostic and therapeutic implications in heart failure. Eur. Heart J. 2016, 37, 2577–2585. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, S.; Thiemermann, C. Role of Metabolic Endotoxemia in Systemic Inflammation and Potential Interventions. Front. Immunol. 2020, 11, 594150. [Google Scholar] [CrossRef] [PubMed]
- Chun, H.W.; Lee, J.; Pham, T.H.; Lee, J.; Yoon, J.H.; Lee, J.; Oh, D.K.; Oh, J.; Yoon, D.Y. Resolvin D5, a Lipid Mediator, Inhibits Production of Interleukin-6 and CCL5 Via the ERK-NF-kappaB Signaling Pathway in Lipopolysaccharide-Stimulated THP-1 Cells. J. Microbiol. Biotechnol. 2020, 30, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Bussmann, A.J.C.; Zaninelli, T.H.; Saraiva-Santos, T.; Fattori, V.; Guazelli, C.F.S.; Bertozzi, M.M.; Andrade, K.C.; Ferraz, C.R.; Camilios-Neto, D.; Casella, A.M.B.; et al. The Flavonoid Hesperidin Methyl Chalcone Targets Cytokines and Oxidative Stress to Reduce Diclofenac-Induced Acute Renal Injury: Contribution of the Nrf2 Redox-Sensitive Pathway. Antioxidants 2022, 11, 1261. [Google Scholar] [CrossRef]
- Borghi, S.M.; Mizokami, S.S.; Carvalho, T.T.; Rasquel-Oliveira, F.S.; Ferraz, C.R.; Fattori, V.; Hayashida, T.H.; Peron, J.P.S.; Camilios-Neto, D.; Ambrosio, S.R.; et al. The diterpene from Sphagneticola trilobata (L.) Pruski, kaurenoic acid, reduces lipopolysaccharide-induced peritonitis and pain in mice. J. Ethnopharmacol. 2021, 273, 113980. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.M.; Fattori, V.; Saito, P.; Pinto, I.C.; Rodrigues, C.C.A.; Melo, C.P.B.; Bussmann, A.J.C.; Staurengo-Ferrari, L.; Bezerra, J.R.; Vignoli, J.A.; et al. The Lipoxin Receptor/FPR2 Agonist BML-111 Protects Mouse Skin Against Ultraviolet B Radiation. Molecules 2020, 25, 2953. [Google Scholar] [CrossRef] [PubMed]
- Bussmann, A.J.C.; Borghi, S.M.; Zaninelli, T.H.; Dos Santos, T.S.; Guazelli, C.F.S.; Fattori, V.; Domiciano, T.P.; Pinho-Ribeiro, F.A.; Ruiz-Miyazawa, K.W.; Casella, A.M.B.; et al. The citrus flavanone naringenin attenuates zymosan-induced mouse joint inflammation: Induction of Nrf2 expression in recruited CD45(+) hematopoietic cells. Inflammopharmacology 2019, 27, 1229–1242. [Google Scholar] [CrossRef]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Role of myeloperoxidase and oxidant formation in the extracellular environment in inflammation-induced tissue damage. Free Radic. Biol. Med. 2021, 172, 633–651. [Google Scholar] [CrossRef]
- Ow, C.P.C.; Trask-Marino, A.; Betrie, A.H.; Evans, R.G.; May, C.N.; Lankadeva, Y.R. Targeting Oxidative Stress in Septic Acute Kidney Injury: From Theory to Practice. J. Clin. Med. 2021, 10, 3798. [Google Scholar] [CrossRef]
- Kehrer, J.P.; Klotz, L.O. Free radicals and related reactive species as mediators of tissue injury and disease: Implications for Health. Crit. Rev. Toxicol. 2015, 45, 765–798. [Google Scholar] [CrossRef]
- Hattori, H.; Subramanian, K.K.; Sakai, J.; Jia, Y.; Li, Y.; Porter, T.F.; Loison, F.; Sarraj, B.; Kasorn, A.; Jo, H.; et al. Small-molecule screen identifies reactive oxygen species as key regulators of neutrophil chemotaxis. Proc. Natl. Acad. Sci. USA 2010, 107, 3546–3551. [Google Scholar] [CrossRef] [PubMed]
- Reshetnikov, V.; Hahn, J.; Maueroder, C.; Czegley, C.; Munoz, L.E.; Herrmann, M.; Hoffmann, M.H.; Mokhir, A. Chemical Tools for Targeted Amplification of Reactive Oxygen Species in Neutrophils. Front. Immunol. 2018, 9, 1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, M.S.N.; Longhi-Balbinot, D.T.; Guazelli, C.F.S.; Navarro, S.A.; Zarpelon, A.C.; Casagrande, R.; Arakawa, N.S.; Verri, W.A., Jr. Chapter 7—Sesquiterpene Lactones: Structural Diversity and Perspectives as Anti-Inflammatory Molecules. In Studies in Natural Products Chemistry; Atta ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 49, pp. 243–264. [Google Scholar]
- Staurengo-Ferrari, L.; Badaro-Garcia, S.; Hohmann, M.S.N.; Manchope, M.F.; Zaninelli, T.H.; Casagrande, R.; Verri, W.A., Jr. Contribution of Nrf2 Modulation to the Mechanism of Action of Analgesic and Anti-inflammatory Drugs in Pre-clinical and Clinical Stages. Front. Pharmacol. 2018, 9, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G.; Group, N.C.R.R.G.W. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, R.; Georgetti, S.R.; Verri, W.A., Jr.; Dorta, D.J.; dos Santos, A.C.; Fonseca, M.J. Protective effect of topical formulations containing quercetin against UVB-induced oxidative stress in hairless mice. J. Photochem. Photobiol. B 2006, 84, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, M.S.; Cardoso, R.D.; Pinho-Ribeiro, F.A.; Crespigio, J.; Cunha, T.M.; Alves-Filho, J.C.; da Silva, R.V.; Pinge-Filho, P.; Ferreira, S.H.; Cunha, F.Q.; et al. 5-lipoxygenase deficiency reduces acetaminophen-induced hepatotoxicity and lethality. Biomed Res. Int. 2013, 2013, 627046. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, J.W.; Cha, Y.N.; Kim, C. A quantitative nitroblue tetrazolium assay for determining intracellular superoxide anion production in phagocytic cells. J. Immunoass. Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, R.D.R.; Chambo, S.D.; Zaninelli, T.H.; Bianchini, B.H.S.; Silva, M.D.V.d.; Bertozzi, M.M.; Saraiva-Santos, T.; Franciosi, A.; Martelossi-Cebinelli, G.; Garcia-Miguel, P.E.; et al. Resolvin D5 (RvD5) Reduces Renal Damage Caused by LPS Endotoxemia in Female Mice. Molecules 2023, 28, 121. https://doi.org/10.3390/molecules28010121
Cardoso RDR, Chambo SD, Zaninelli TH, Bianchini BHS, Silva MDVd, Bertozzi MM, Saraiva-Santos T, Franciosi A, Martelossi-Cebinelli G, Garcia-Miguel PE, et al. Resolvin D5 (RvD5) Reduces Renal Damage Caused by LPS Endotoxemia in Female Mice. Molecules. 2023; 28(1):121. https://doi.org/10.3390/molecules28010121
Chicago/Turabian StyleCardoso, Renato D. R., Sandmary D. Chambo, Tiago H. Zaninelli, Beatriz H. S. Bianchini, Matheus Deroco Veloso da Silva, Mariana M. Bertozzi, Telma Saraiva-Santos, Anelise Franciosi, Geovana Martelossi-Cebinelli, Pamela E. Garcia-Miguel, and et al. 2023. "Resolvin D5 (RvD5) Reduces Renal Damage Caused by LPS Endotoxemia in Female Mice" Molecules 28, no. 1: 121. https://doi.org/10.3390/molecules28010121
APA StyleCardoso, R. D. R., Chambo, S. D., Zaninelli, T. H., Bianchini, B. H. S., Silva, M. D. V. d., Bertozzi, M. M., Saraiva-Santos, T., Franciosi, A., Martelossi-Cebinelli, G., Garcia-Miguel, P. E., Borghi, S. M., Casagrande, R., & Verri, W. A. (2023). Resolvin D5 (RvD5) Reduces Renal Damage Caused by LPS Endotoxemia in Female Mice. Molecules, 28(1), 121. https://doi.org/10.3390/molecules28010121