
Isolation and Characterization of Antibacterial Conglutinins from Lupine Seeds

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of Lupine Protein Isolate (LPI), 7S and 11S Globulins, and Basic Subunit (BS) from Lupine Seeds
2.1.1. SDS-PAGE and Urea-PAGE of 7S and 11S Globulins and BS
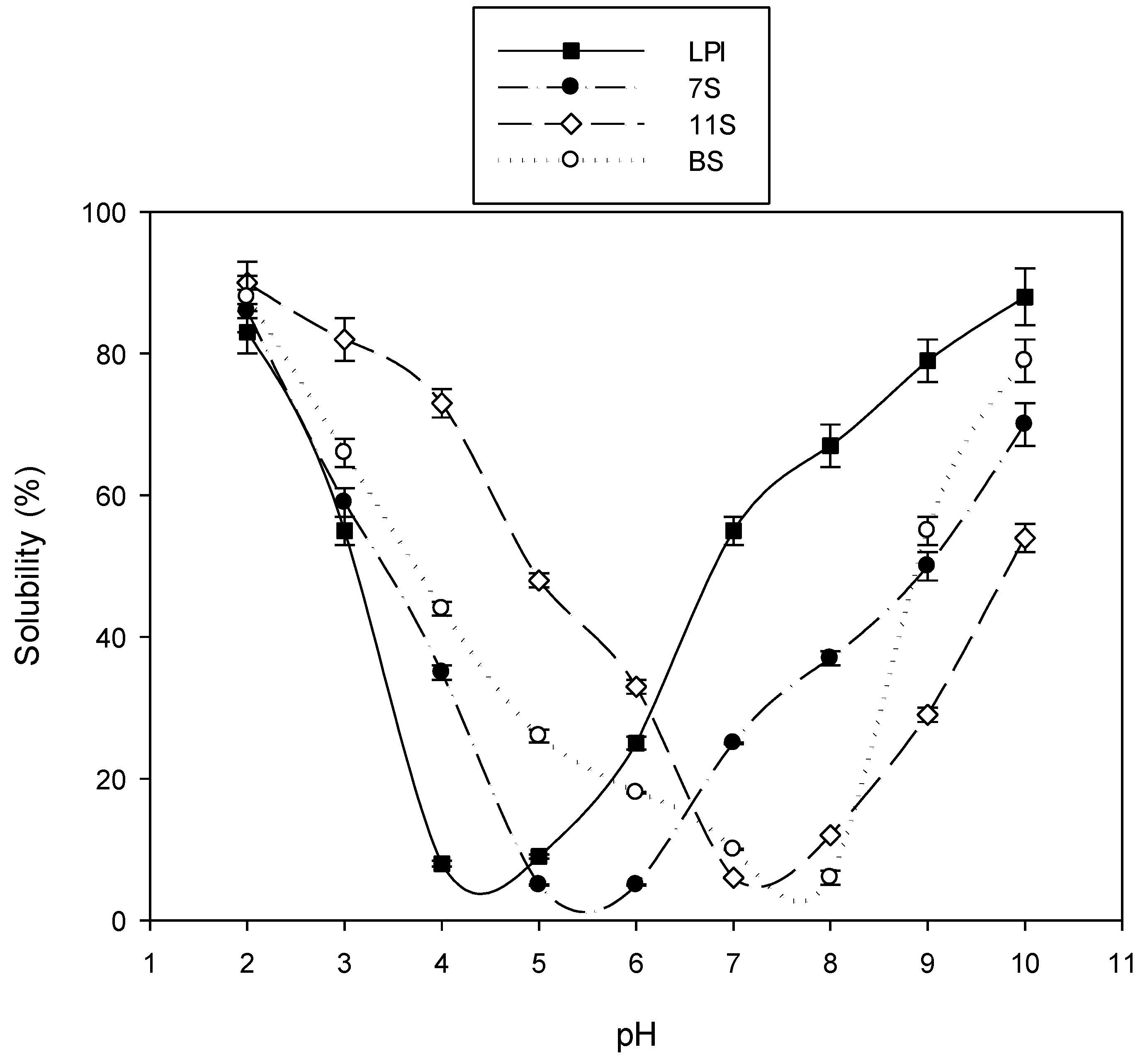
2.1.2. pH–Solubility Curve
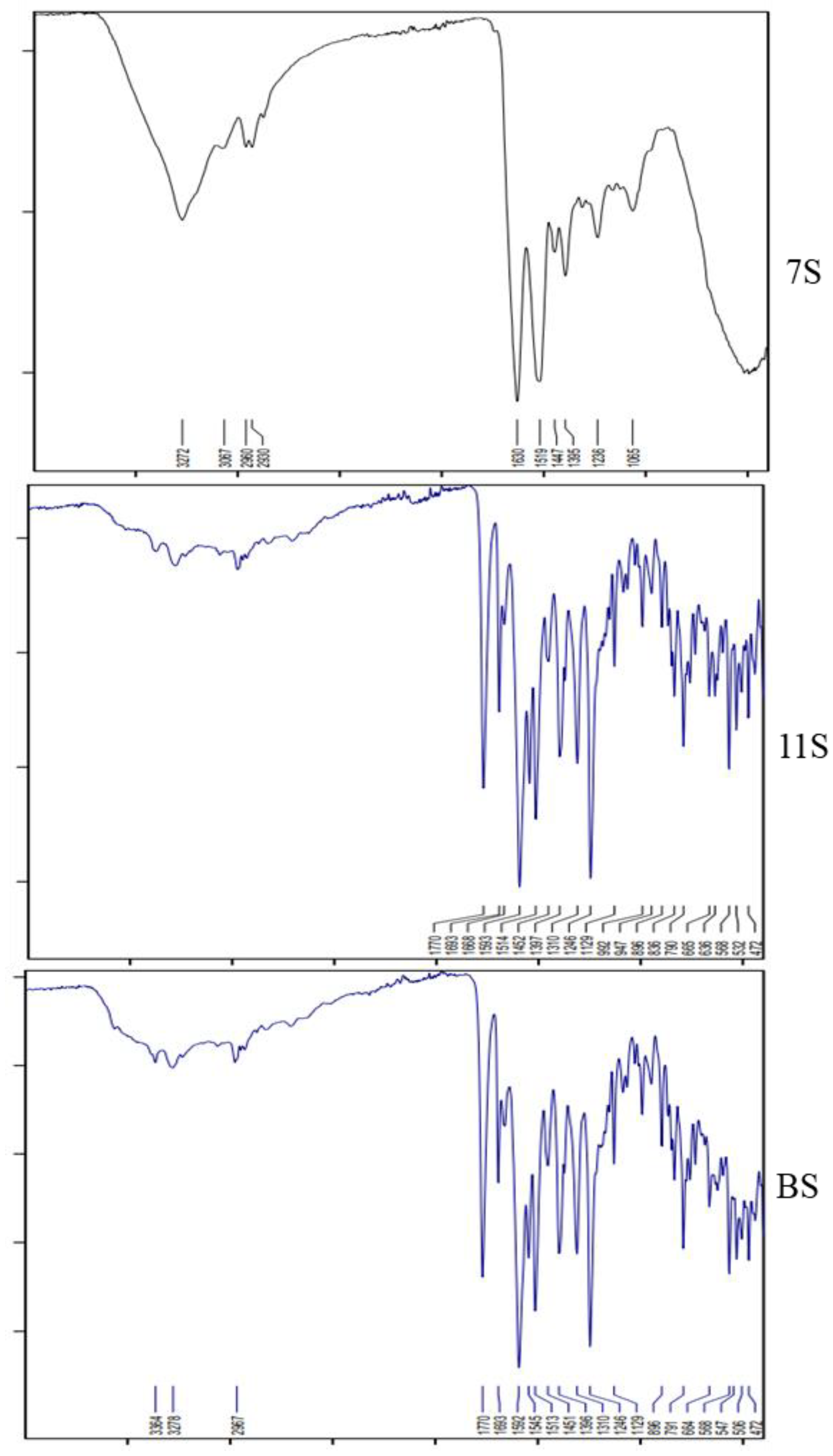
2.1.3. FT-IR Spectroscopy
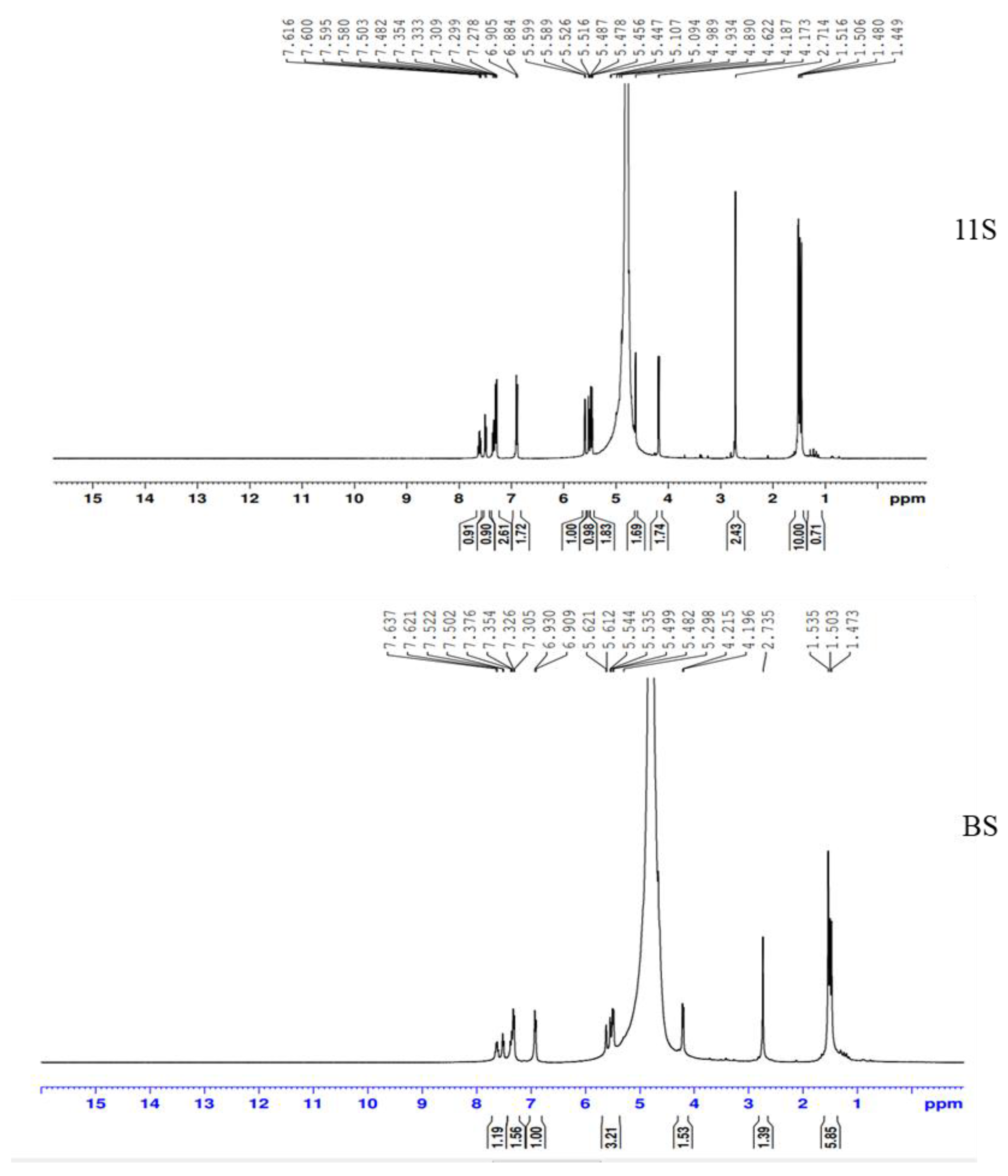
2.1.4. 1H NMR Spectroscopy
2.1.5. Total Phenolic Compounds
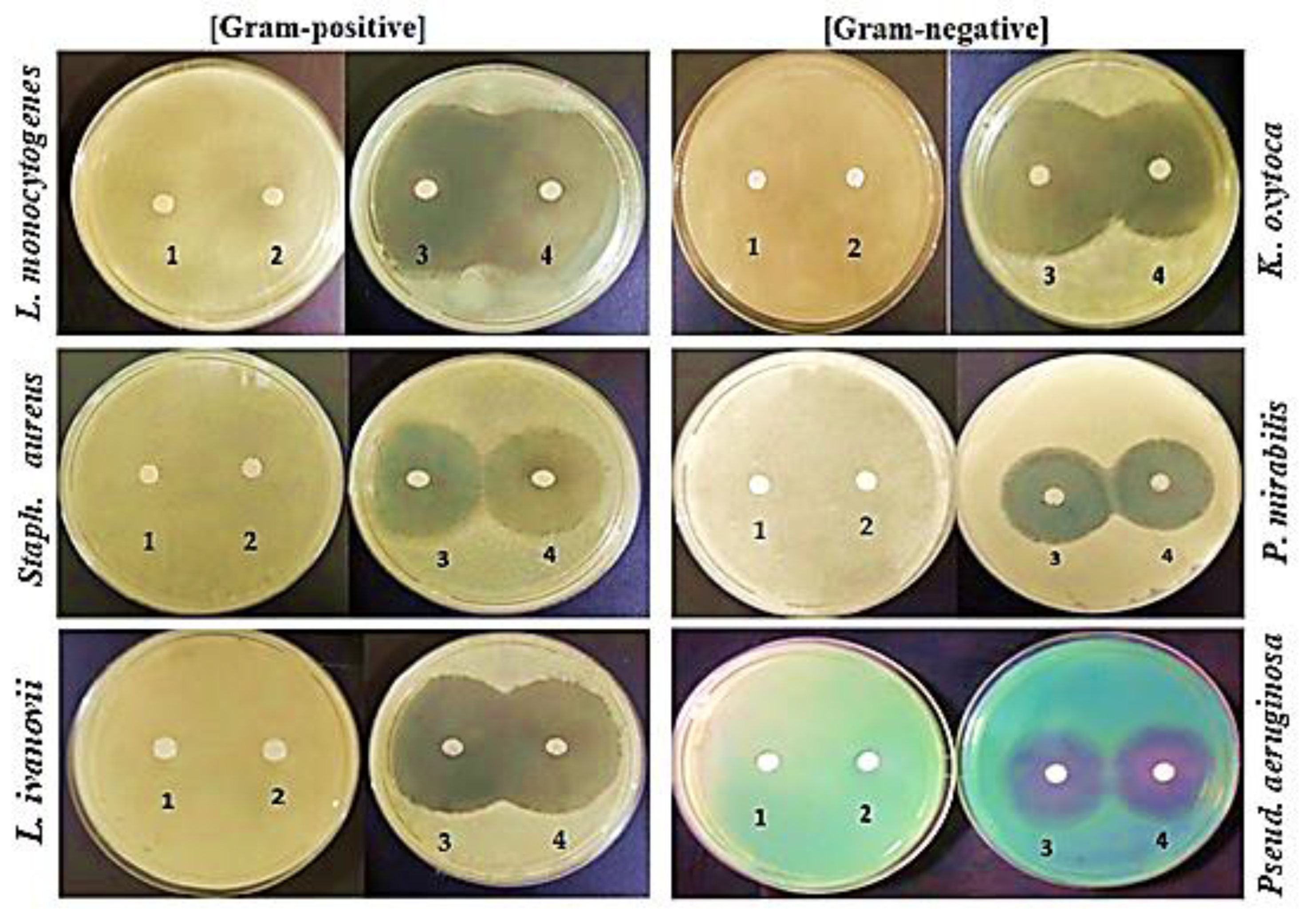
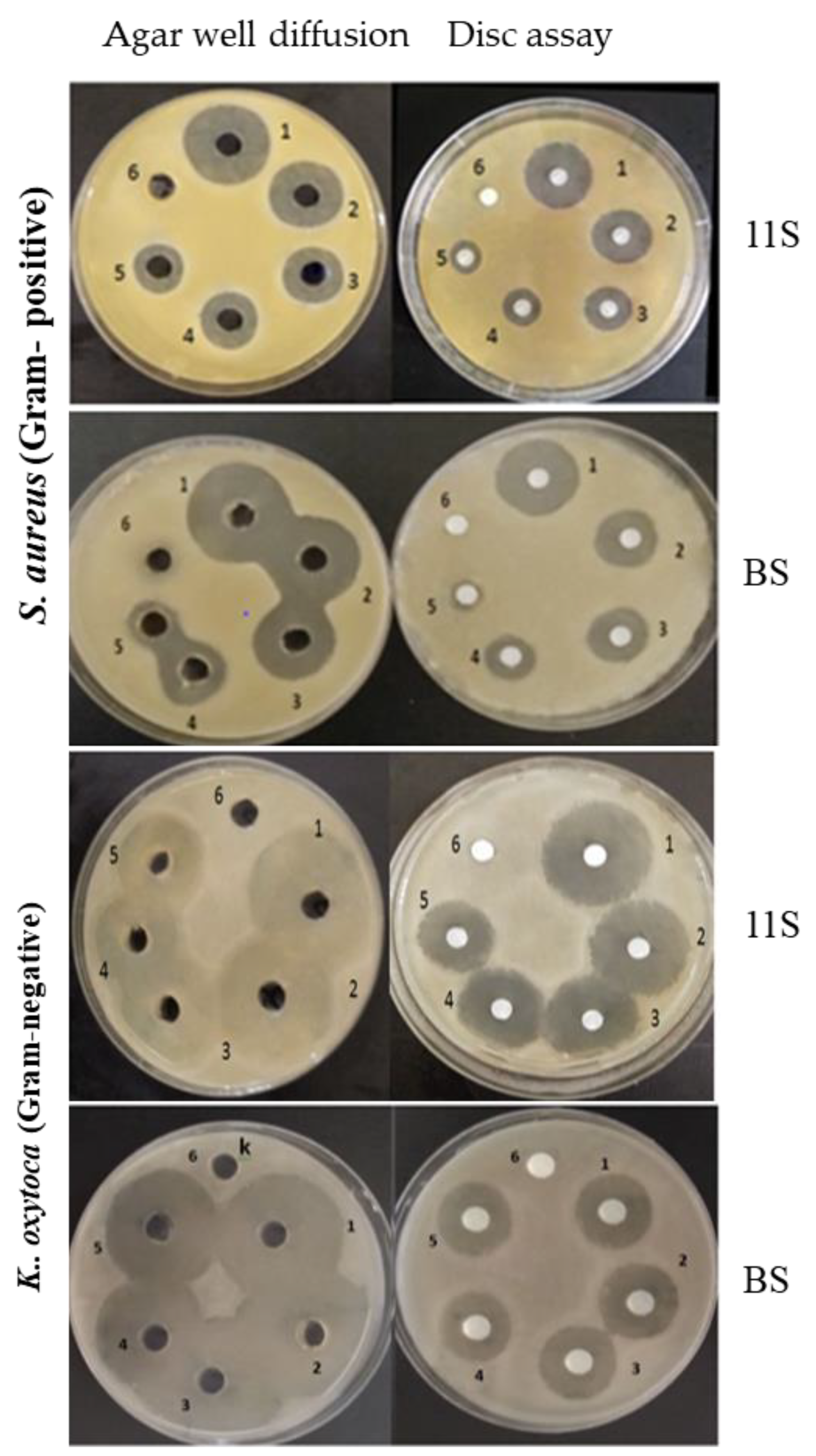
2.2. Antibacterial Activity of LPI, 7S and 11S Globulins, and BS against Gram-Positive and Gram-Negative Bacteria
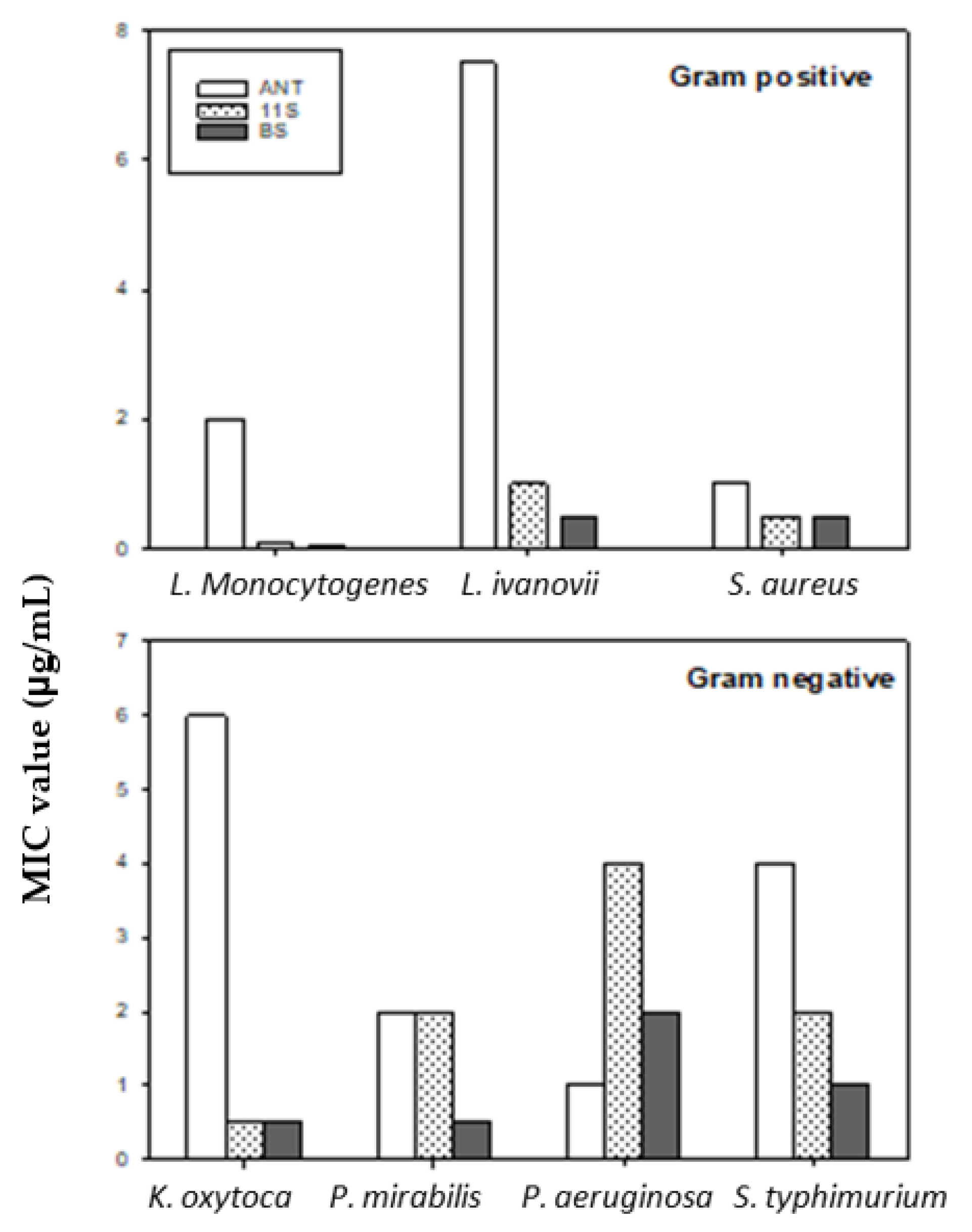
2.3. Minimum Inhibitory Concentration (MIC)
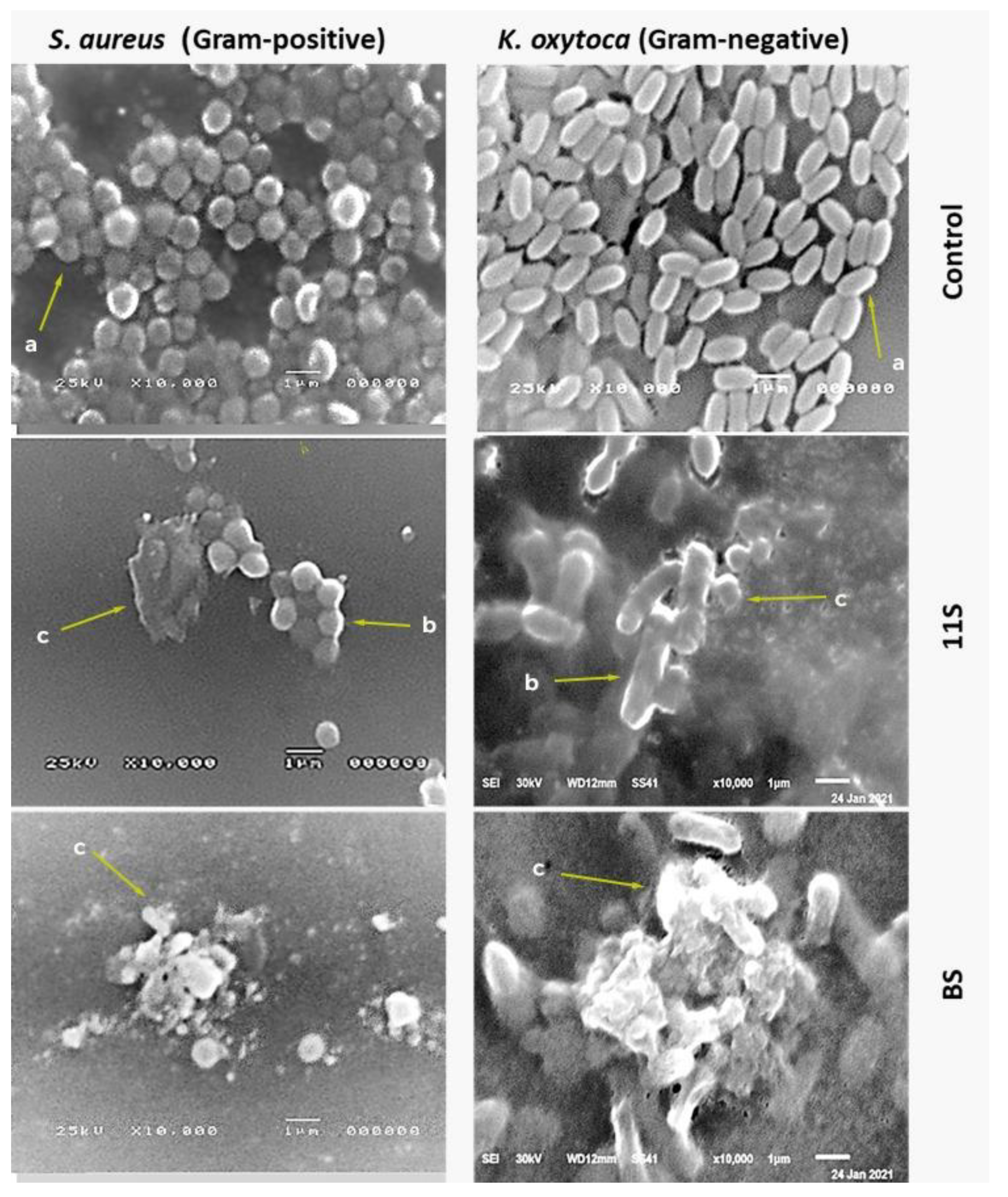
2.4. SEM Image Analysis
2.5. Comparison of the MIC of Tested Lupine (11S and BS) with Specific Antibiotics for Each Microorganism
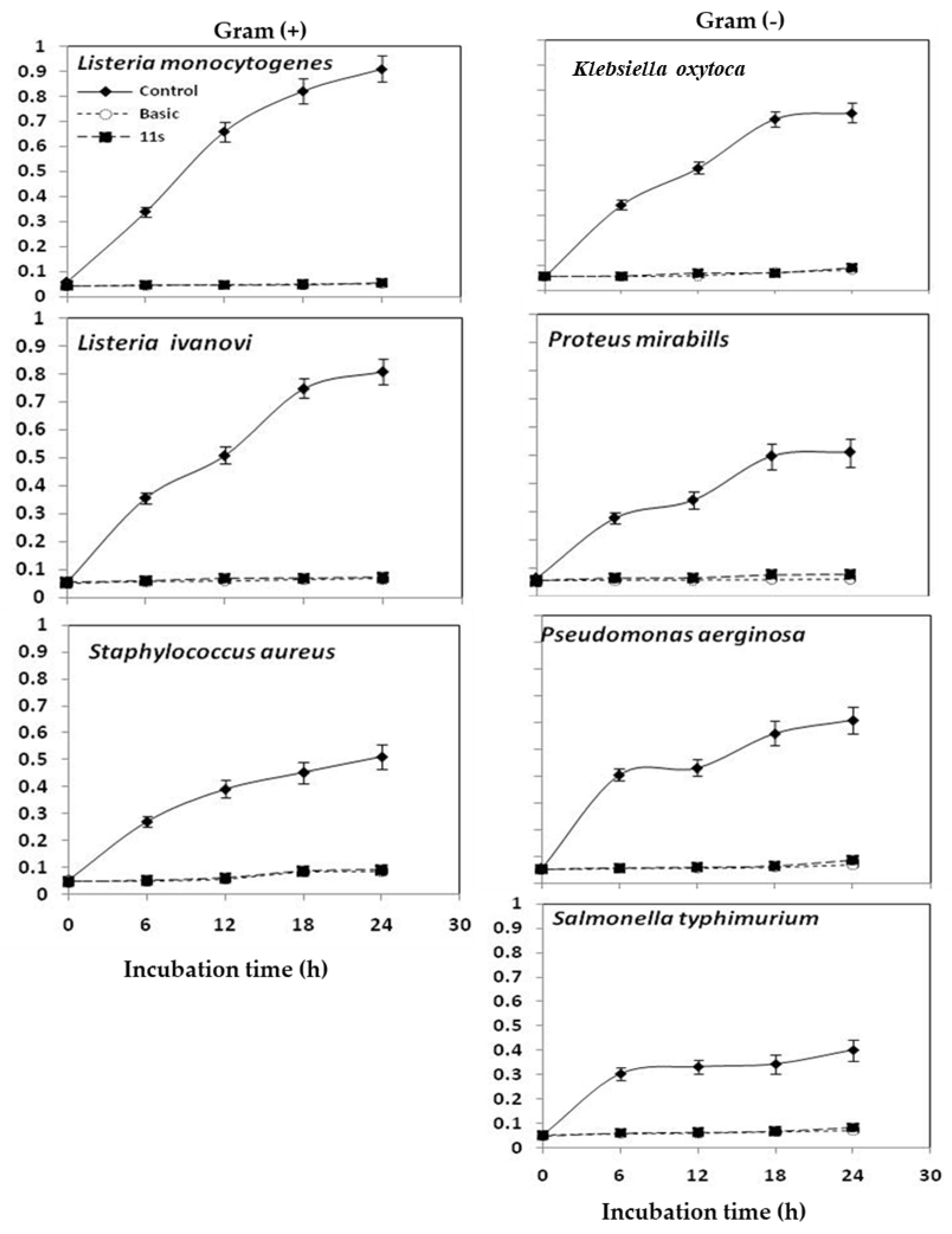
2.6. Quantitative Inhibition of Pathogenic Bacteria by 11S Globulin and BS
3. Discussion
4. Materials and Methods
4.1. Isolation and Chemical Characterization of Lupine Protein Isolate (LPI), 7S and 11S Globulins, and Basic Subunit (BS) from Lupine Seeds
4.1.1. Sample Preparation
4.1.2. Extraction of Protein Isolate
4.1.3. Isolation of 7S and 11S Globulins and Basic Subunit (BS) from Lupine
4.1.4. Sodium Dodecyl-Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) of Proteins
4.1.5. Urea-PAGE of 7S and 11S Globulins and BS
4.1.6. pH–Solubility Curve
4.1.7. Fourier-Transform Infrared (FT-IR) Spectroscopy
4.1.8. 1H Nuclear Magnetic Resonance (1H NMR) Spectroscopy
4.1.9. Total Phenolic Compounds (TPCs) and Total Flavonoid Content (TFC) Estimation
4.2. Antibacterial Activities of LPI, 7S and 11S Globulins, and BS against Gram-Negative and Gram-Positive Bacteria
4.2.1. Minimum Inhibitory Concentration (MIC)
Agar Well Diffusion Assay
Disk Diffusion Assay
4.2.2. Scanning Electron Microscopy (SEM)
4.2.3. Comparison of MICs of Tested Lupine (11S Globulin and BS) with Specific Antibiotics for Each Microorganism
4.2.4. Quantitative Inhibition of Pathogenic Bacteria by 11S Globulin and BS
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Golkar, Z.; Bagasra, O.; Pace, D.G. Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries 2014, 8, 129–136. [Google Scholar] [CrossRef]
- Adekanye, U.O.; Ekiri, A.B.; Galipó, E.; Muhammad, A.B.; Mateus, A.; La Ragione, R.M.; Wakawa, A.; Armson, B.; Mijten, E.; Alafiatayo, R. Knowledge, attitudes and practices of veterinarians towards antimicrobial resistance and stewardship in Nigeria. Antibiotics 2020, 9, 453. [Google Scholar] [CrossRef] [PubMed]
- Vivas, R.; Barbosa, A.A.T.; Dolabela, S.S.; Jain, S. Multidrug-resistant bacteria and alternative methods to control them: An overview. Microb. Drug Resist. 2019, 25, 890–908. [Google Scholar] [CrossRef] [PubMed]
- Maiyo, Z.; Ngure, R.; Matasyoh, J.; Chepkorir, R. Phytochemical constituents and antimicrobial activity of leaf extracts of three Amaranthus plant species. Afr. J. Biotechnol. 2010, 9, 3178–3182. [Google Scholar]
- Enan, G.; Abdel-Shafi, S.; Abdel-Haliem, M.; Negm, S. Characterization of probiotic lactic acid bacteria to be used as starter and protective cultures for dairy fermentations. Int. J. Probiotics Prebiotics 2013, 8, 157–164. [Google Scholar]
- Enan, G.; Al-Mohammadi, A.; El-Didamony, G.; Abdel-Haliem, M.; Zakaria, A. Antimicrobial activity of Enterococcus faecium NM2 isolated from urine: Purification, characterization and bactericidal action of enterocin NM2. Asian J. Appl. Sci. 2014, 7, 721–734. [Google Scholar] [CrossRef] [Green Version]
- Enan, G.; El-Didamony, G.; Mohamed, E.; Zakaria, A. Novel antibacterial activity of Enterococcus faecium NM2 isolated from urine of healthy people. Asian J. Appl. Sci. 2014, 7, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Erfan, A.M.; Marouf, S. Cinnamon oil downregulates virulence genes of poultry respiratory bacterial agents and revealed significant bacterial inhibition: An in vitro perspective. Veter-World 2019, 12, 1707. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.; Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Kamal, N.; Enan, G.; Sitohy, M. Catfish glycoprotein, a highly powerful safe preservative of minced beef stored at 4 C for 15 days. Foods 2020, 9, 1115. [Google Scholar] [CrossRef]
- Osman, A.; El-Didamony, G.; Sitohy, M.; Khalifa, M.; Enan, G. Soybean glycinin basic subunit inhibits methicillin resistant-vancomycin intermediate Staphylococcus aureus (MRSA-VISA) in vitro. Int. J. Appl. Res. Nat. Prod. 2016, 9, 17–26. [Google Scholar]
- Hudson, M.A.; Lockless, S.W. Elucidating the Mechanisms of Action of Antimicrobial Agents. mBio 2022, 13, e02240-02221. [Google Scholar] [CrossRef]
- Beuchat, L. Antimicrobial properties of spices and their essential oils. Food Preserv. 1994, 12, 257–262. [Google Scholar]
- Kumaravel, S.; Muthukumaran, P.; Thomas, N. Phytochemical, GC-MS and FT-IR analysis of Papver somniferum. J. Pharm. Biol. Sci. 2019, 7, 1–8. [Google Scholar]
- Arora, D.S.; Kaur, J. Antimicrobial activity of spices. Int. J. Antimicrob. Agents 1999, 12, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.; Bersani, C.; Comi, G. Antimicrobial activity of the essential oils of Thymus vulgaris L. measured using a bioimpedometric method. Food Prot. 1999, 62, 1017–1023. [Google Scholar] [CrossRef]
- Penalver, P.; Huerta, B.; Borge, C.; Astorga, R.; Romero, R.; Perea, A. Antimicrobial activity of five essential oils against origin strains of the Enterobacteriaceae family. Apmis 2005, 113, 1–6. [Google Scholar] [CrossRef]
- Mahgoub, S.; Osman, A.; Sitohy, M. Inhibition of growth of pathogenic bacteria in raw milk by legume protein esters. J. Food Prot. 2011, 74, 1475–1481. [Google Scholar] [CrossRef]
- Sitohy, M.; Mahgoub, S.; Osman, A.; El-Masry, R.; Al-Gaby, A. Extent and mode of action of cationic legume proteins against Listeria monocytogenes and Salmonella Enteritidis. Probiotics Antimicrob. Proteins 2013, 5, 195–205. [Google Scholar] [CrossRef]
- Osman, A.O.; Mahgoub, S.A.; Sitohy, M.Z. Preservative action of 11S (glycinin) and 7S (β-conglycinin) soy globulin on bovine raw milk stored either at 4 or 25 C. J. Dairy Res. 2013, 80, 174–183. [Google Scholar] [CrossRef]
- Osman, A.; Mahgoub, S.; Sitohy, M.Z. Hindering milk quality storage deterioration by mild thermization combined with methylated chickpea protein. Int. Food Res. J. 2014, 21, 693–701. [Google Scholar]
- Abdel-Hamid, M.; Goda, H.A.; De Gobba, C.; Jenssen, H.; Osman, A. Antibacterial activity of papain hydrolysed camel whey and its fractions. Int. Dairy J. 2016, 61, 91–98. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A. Antimicrobial activity of native and esterified legume proteins against Gram-negative and Gram-positive bacteria. Food Chem. 2010, 120, 66–73. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A.; Ghany, A.; Salama, A. Antibacterial phycocyanin from Anabaena oryzae SOS13. Int. J. Appl. Res. Nat. Prod. 2015, 8, 27–36. [Google Scholar]
- Osman, A.; Goda, H.A.; Abdel-Hamid, M.; Badran, S.M.; Otte, J. Antibacterial peptides generated by Alcalase hydrolysis of goat whey. LWT-Food Sci. Technol. 2016, 65, 480–486. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Osman, A.; Enan, G.; El-Nemer, M.; Sitohy, M. Antibacterial activity of methylated egg white proteins against pathogenic G+ and G-bacteria matching antibiotics. SpringerPlus 2016, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Osman, A.O. Enhancing milk preservation with esterified legume proteins. Probiotics Antimicrob. Proteins 2011, 3, 48–56. [Google Scholar] [CrossRef]
- Joray, M.; Rayas-Duarte, P.; Mohamed, A.; Van Santen, E. Coated lupin bean snacks. J. Food Qual. 2007, 30, 267–279. [Google Scholar] [CrossRef]
- Yeheyis, L.; Kijora, C.; Wink, M.; Peters, K.J. Effect of a traditional processing method on the chemical composition of local white lupin (Lupinus albus L.) seed in North-Western Ethiopia. Z. Nat. C 2011, 66, 403–408. [Google Scholar]
- Abd Elhamid, M.A.; Mandour, A.E.S.; Ismail, T.A.; Al-Zohairy, A.M.; Almowallad, S.; Alqahtani, L.S.; Osman, A. Powerful Antioxidants and Cytotoxic Activities of the Methanol Extracts from Eight Soybean Cultivars. Molecules 2022, 27, 2895. [Google Scholar] [CrossRef]
- Torres, A.; Frias, J.; Vidal-Valverde, C. Changes in chemical composition of lupin seeds (Lupinus angustifolius) after selective α-galactoside extraction. J. Sci. Food Agric. 2005, 85, 2468–2474. [Google Scholar] [CrossRef]
- Cerletti, P.; Fumagalli, A.; Venturin, D. Protein composition of seeds of Lupinus albus. J. Food Sci. 1978, 43, 1409–1411. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Mahgoub, S.A.; Osman, A.O. In vitro and in situ antimicrobial action and mechanism of glycinin and its basic subunit. Int. J. Food Microbiol. 2012, 154, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-Q.; Sun, X.-X.; Feng, J.-L.; Mo, H.-Z. Antibacterial activities and membrane permeability actions of glycinin basic peptide against Escherichia coli. Innov. Food Sci. Emerg. Technol. 2015, 31, 170–176. [Google Scholar] [CrossRef]
- Yang, J.; Sun, G.-J.; Li, Y.-Q.; Cui, K.-Y.; Mo, H.Z. Antibacterial characteristics of glycinin basic polypeptide against Staphylococcus aureus. Food Sci. Biotechnol. 2016, 25, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.-P.; Li, Y.-Q.; Sun, G.-J.; Mo, H.-Z. Antibacterial actions of glycinin basic peptide against Escherichia coli. J. Agric. Food Chem. 2017, 65, 5173–5180. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A. Bioactive compounds in soybean proteins and its applications in food systems. Sustain. Agric. Environ. Egypt Part I 2018, 147–160. [Google Scholar]
- Santiago Quiles, M.R.; Oquendo-Jiménez, I.; Herreño-Saénz, D.; Antoun, M.D. Genotoxicity of alkaloid-rich extract from Lupinus termis seeds. Pharm. Crops 2010, 1, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, E.M.; Qureshi, K.A.; Musa, K.H. Antimicrobial, antioxidant and phytochemical screening of Lupin seeds (Lupinus termis Forrsk.) from Sudan. CIBTech J. Microbiol. 2017, 6, 1–8. [Google Scholar]
- Hanania, M.; Radwan, S.; Karmi, E. Extraction method and evaluation of phenolics, flavonoids, antioxidant activity, antimicrobial activity and minerals of bitter Lupinus albus in Palestine. J. Biol. Act. Prod. Nat. 2018, 8, 137–143. [Google Scholar] [CrossRef]
- Krakowska-Sieprawska, A.; Rafińska, K.; Walczak-Skierska, J.; Buszewski, B. The influence of plant material enzymatic hydrolysis and extraction conditions on the polyphenolic profiles and antioxidant activity of extracts: A green and efficient approach. Molecules 2020, 25, 2074. [Google Scholar] [CrossRef] [PubMed]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major food-borne zoonotic bacterial pathogens. J. Trop. Med. 2020, 4674235. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Osman, A.; Enan, G.; Abdel-Hameid, S.; Sitohy, M. Characterization and antibacterial activity of 7S and 11S globulins isolated from cowpea seed protein. Molecules 2019, 24, 1082. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E. Mechanisms of action of newer antibiotics for Gram-positive pathogens. Lancet Infect. Dis. 2005, 5, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Enan, G.; Al-Mohammadi, A.-R.; Abdel-Shafi, S.; Abdel-Hameid, S.; Sitohy, M.Z.; El-Gazzar, N. Antibacterial peptides produced by Alcalase from cowpea seed proteins. Antibiotics 2021, 10, 870. [Google Scholar] [CrossRef]
- Johnson, E.A.; Brekke, C. Functional properties of acylated pea protein isolates. J. Food Sci. 1983, 48, 722–725. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.W.; Williams, J. The electrophoresis of transferrins in urea/polyacrylamide gels. Biochem. J. 1980, 189, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chobert, J.-M.; Touati, A.; Bertrand-Harb, C.; Dalgalarrondo, M.; Nicolas, M.-G.; Haertle, T. In vitro proteolysis and functional properties of reductively alkylated β-casein derivatives. J. Dairy Res. 1991, 58, 285–298. [Google Scholar] [CrossRef]
- Souillac, P.O.; Middaugh, C.R.; Rytting, J.H. Investigation of protein/carbohydrate interactions in the dried state. 2. Diffuse reflectance FTIR studies. Int. J. Pharm. 2002, 235, 207–218. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Ordonez, A.; Gomez, J.; Vattuone, M. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Nanda, A.; Saravanan, M. Biosynthesis of silver nanoparticles from Staphylococcus aureus and its antimicrobial activity against MRSA and MRSE. Nanomed. Nanotechnol. Biol. Med. 2009, 5, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- Mojsoska, B.; Carretero, G.; Larsen, S.; Mateiu, R.V.; Jenssen, H. Peptoids successfully inhibit the growth of gram negative E. coli causing substantial membrane damage. Sci. Rep. 2017, 7, 42332. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Inhibitory Action | |||||||||
| Lupine 11S Globulin Concentration (µg mL−1) | |||||||||
| 0.05 | 0.1 | 0.3 | 0.5 | 1 | 2 | 4 | 8 | ||
| Microorganism | Inhibition Zone Diameter (mm) ± SE | ||||||||
| Agar well diffusion | L. monocytogenes * | 0.000 q ± 0 | 16.000 o ± 0 | 28.00 l ± 0.58 | 30.00 ijk ± 0 | 29.333 jkl ± 0.33 | 33.000 g ± 1 | 46.000 c ± 0.58 | 49.000 a ± 1 |
| K. oxytoca | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 31.00 hi ± 0.58 | 33.000 g ± 1.73 | 47.000 bc ± 0 | 48.000 ab ± 0.58 | 49.000 a ± 0 | |
| L. ivanovii | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 13.000 p ± 0.58 | 29.333 jkl ± 0.88 | 36.000 f ± 0.58 | ||
| S. aureus | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 15.33 o ± 1.2 | 24.000 n ± 0 | 26.000 m ± 1.15 | 30.333 ij ± 0.67 | 39.667 e ± 0.33 | |
| P. mirabilis | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 28.000 l ± 0.58 | 30.000 ijk ± 0.58 | 32.000 gh ± 0.58 | |
| P. aeruginosa | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 28.667 kl ± 0.67 | 31.000 hi ± 0 | |
| S. typhimurium | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 0.000 q ± 0 | 24.000 n ± 0.58 | 25.000 mn ± 0 | 32.000 gh ± 1.15 | |
| Disk assay | L. monocytogenes | 0.000 p ± 0 | 5.000 no ± 0 | 15.67 ghij ± 0.33 | 12.00 jkl ± 0 | 13.00 ijkl ± 0.58 | 21.33 def ± 0.33 | 22.00 cde ± 0.58 | 25.00 abd ± 0.58 |
| K. oxytoca | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 18.83 efg ± 0.44 | 17.7 efgh ± 0.33 | 24.66 bcd ± 0.33 | 24.66 bcd ± 0.33 | 27.167 ab ± 0.17 | |
| L. ivanovii | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 9.00 lmn ± 0.58 | 13.00 ijkl ± 0.58 | 14.33 hijk ± 0.33 | 29.000 a ± 0 | |
| S. aureus | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 op ± 2 | 7.00 mn ± 1 | 14.00 hijk ± 3.51 | 22.00 cde ± 0.58 | 26.00 abc ± 0.58 | 18.000 efgh ± 9 | |
| P. mirabilis | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 14.00 hijk ± 0.58 | 15.000 ghijk ± 0 | 19.000 efg ± 0.58 | |
| P. aeruginosa | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 16.000 ghij ± 0 | 18.000 efgh ± 0.58 | |
| S. typhimurium | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 0.000 p ± 0 | 11.00 klm ± 0.58 | 14.00 hijk ± 1.15 | 17.000 fghi ± 0 | |
| (B) Statistical Significance of Microorganism and Concentration Effects | |||||||||
| Microorganism | Microorganism Effect | Concentration (µg mL−1) | Concentration Effect | ||||||
| Agar Well Dif | Disk Assay | Agar Well Dif | Disk Assay | ||||||
| L. monocytogenes | 28.917 a ± 3.06 | 14.250 a ± 1.69 | 0.05 | 0.000 h ± 0 | 0.000 g ± 0 | ||||
| K. oxytoca | 26.000 b ± 4.4 | 14.125 a ± 2.36 | 0.1 | 2.286 g ± 1.25 | 0.714 g ± 0.39 | ||||
| L. ivanovii | 15.333 d ± 3.62 | 8.167 c ± 2.04 | 0.3 | 4.000 f ± 2.19 | 2.524 f ± 1.23 | ||||
| S. aureus | 16.917 c ± 3.04 | 11.125 b ± 2.26 | 0.5 | 10.905 e ± 3.01 | 5.405 e ± 1.57 | ||||
| P. mirabilis | 11.250 e ± 3.04 | 6.000 d ± 1.64 | 1 | 14.190 d ± 3.04 | 7.667 d ± 1.63 | ||||
| P. aeruginosa | 7.458 g ± 2.7 | 4.250 e ± 1.54 | 2 | 26.762 c ± 2.91 | 15.143 c ± 1.76 | ||||
| S. typhimurium | 10.125 f ± 2.77 | 5.250 de ± 1.45 | 4 | 34.857 b ± 1.86 | 18.857 b ± 1.09 | ||||
| 8 | 39.571 a ± 1.68 | 21.881 a ± 1.5 | |||||||
| (A) Inhibitory Action | |||||||||
| Lupine BS Concentration (µg mL−1) | |||||||||
| 0.05 | 0.1 | 0.3 | 0.5 | 1 | 2 | 4 | 8 | ||
| Inhibition Zone Diameter (mm) ± SE | |||||||||
| Agar well diffusion | L. monocytogenes | 7.000 q ± 0 | 9.000 p ± 0.58 | 28.000 kl ± 0.58 | 38.000 gh ± 0.58 | 42.000 e ± 0 | 47.000 c ± 0.58 | 49.000 b ± 0 | 51.667 a ± 1.67 |
| K. oxytoca | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 27.000 l ± 0 | 35.000 i ± 0.58 | 37.333 h ± 0.88 | 45.000 d ± 0.58 | 49.000 b ± 0 | |
| L. ivanovii | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 25.000 m ± 0.58 | 34.000 i ± 0.58 | 16.000 o ± 0.58 | 27.000 l ± 0.58 | 40.000 f ± 0.58 | |
| S. aureus | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 8.000 pq ± 0.58 | 21.000 n ± 0.58 | 28.000 kl ± 0.58 | 37.000 h ± 0 | 39.000 fg ± 0 | |
| P. mirabilis | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 15.000 o ± 0 | 28.000 kl ± 0.58 | 28.000 kl ± 0 | 31.000 j ± 0.58 | 37.000 h ± 0.58 | |
| P. aeruginosa | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 31.000 j ± 0.58 | 32.000 j ± 0.58 | 34.000 i ± 0 | |
| S. typhimurium | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 0.000 r ± 0 | 24.000 m ± 0.58 | 28.000 kl ± 0.58 | 29.000 k ± 0 | 29.000 k ± 0.58 | |
| Disk assay | L. monocytogenes | 10.00 pq ± 0 | 14.00 m ± 0.58 | 15.66 l ± 1.45 | 18.333 jk ± 0.88 | 32.000 f ± 0 | 35.000 de ± 0 | 41.000 b ± 0.58 | 43.000 a ± 0.58 |
| K. oxytoca | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 17.000 kl ± 0 | 32.000 f ± 0.58 | 34.000 e ± 0.58 | 36.000 d ± 0 | 39.000 c ± 0.58 | |
| L. ivanovii | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 5.00 s ± 0.58 | 14.000 m ± 0 | 16.000 l ± 0.58 | 27.000 h ± 0.58 | 35.000 de ± 0.58 | |
| S. aureus | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 5.00 s ± 0 | 11.000 op ± 0.58 | 13.000 mn ± 0.58 | 29.000 g ± 0.58 | 30.000 g ± 0.58 | |
| P. mirabilis | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 10.00 pq ± 0.58 | 12.000 no ± 0.58 | 18.000 jk ± 0.58 | 19.000 j ± 0.58 | 25.000 i ± 0.58 | |
| P. aeruginosa | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 16.000 l ± 0.58 | 16.333 l ± 0.33 | 18.000 jk ± 0.58 | |
| S. typhimurium | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 0.000 t ± 0 | 8.000 r ± 0.58 | 9.000 qr ± 0.58 | 10.000 pq ± 0.58 | 17.000 kl ± 0.58 | |
| (B) Statistical Significance of Microorganism and Concentration Effects | |||||||||
| Microorganism | Microorganism Effect | Concentration (µg mL−1) | Concentration Effect | ||||||
| Agar Well Diffusion | Disk Assay | Agar Well Diffusion | Disk Assay | ||||||
| L. monocytogenes | 33.958 a ± 3.45 | 26.125 a ± 2.56 | 0.05 | 1.000 g ± 0.55 | 1.429 g ± 0.78 | ||||
| K. oxytoca | 24.167 b ± 4.11 | 19.750 b ± 3.43 | 0.1 | 1.286 g ± 0.71 | 2.000 f ± 1.1 | ||||
| L. ivanovii | 17.750 c ± 3.17 | 12.125 c ± 2.61 | 0.3 | 4.000 f ± 2.19 | 2.238 f ± 1.24 | ||||
| S. aureus | 16.625 d ± 3.27 | 11.000 d ± 2.43 | 0.5 | 16.143 e ± 3.01 | 7.905 e ± 1.56 | ||||
| P. mirabilis | 17.375 c ± 3.05 | 10.500 e ± 1.92 | 1 | 26.286 d ± 2.82 | 15.571 d ± 2.5 | ||||
| P. aeruginosa | 12.125 f ± 3.27 | 6.292 f ± 1.7 | 2 | 30.762 c ± 1.99 | 20.143 c ± 2.12 | ||||
| S. typhimurium | 13.750 e ± 2.89 | 5.500 g ± 1.27 | 4 | 35.714 b ± 1.74 | 25.476 b ± 2.29 | ||||
| 8 | 39.952 a ± 1.67 | 29.571 a ± 2.1 | |||||||
| Microorganism | MIC (µg/mL) | |
|---|---|---|
| 11S | BS | |
| L. monocytogenes | 0.1 | 0.05 |
| K. oxytoca | 0.5 | 0.5 |
| L. ivanovii | 1 | 0.5 |
| S. aureus | 0.5 | 0.5 |
| P. mirabilis | 2 | 0.5 |
| P. aeruginosa | 4 | 2 |
| S. typhimurium | 2 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Shafi, S.; El-Nemr, M.; Enan, G.; Osman, A.; Sitohy, B.; Sitohy, M. Isolation and Characterization of Antibacterial Conglutinins from Lupine Seeds. Molecules 2023, 28, 35. https://doi.org/10.3390/molecules28010035
Abdel-Shafi S, El-Nemr M, Enan G, Osman A, Sitohy B, Sitohy M. Isolation and Characterization of Antibacterial Conglutinins from Lupine Seeds. Molecules. 2023; 28(1):35. https://doi.org/10.3390/molecules28010035
Chicago/Turabian StyleAbdel-Shafi, Seham, Mona El-Nemr, Gamal Enan, Ali Osman, Basel Sitohy, and Mahmoud Sitohy. 2023. "Isolation and Characterization of Antibacterial Conglutinins from Lupine Seeds" Molecules 28, no. 1: 35. https://doi.org/10.3390/molecules28010035
APA StyleAbdel-Shafi, S., El-Nemr, M., Enan, G., Osman, A., Sitohy, B., & Sitohy, M. (2023). Isolation and Characterization of Antibacterial Conglutinins from Lupine Seeds. Molecules, 28(1), 35. https://doi.org/10.3390/molecules28010035