
Luteolin Isolated from Juncus acutus L., a Potential Remedy for Human Coronavirus 229E

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
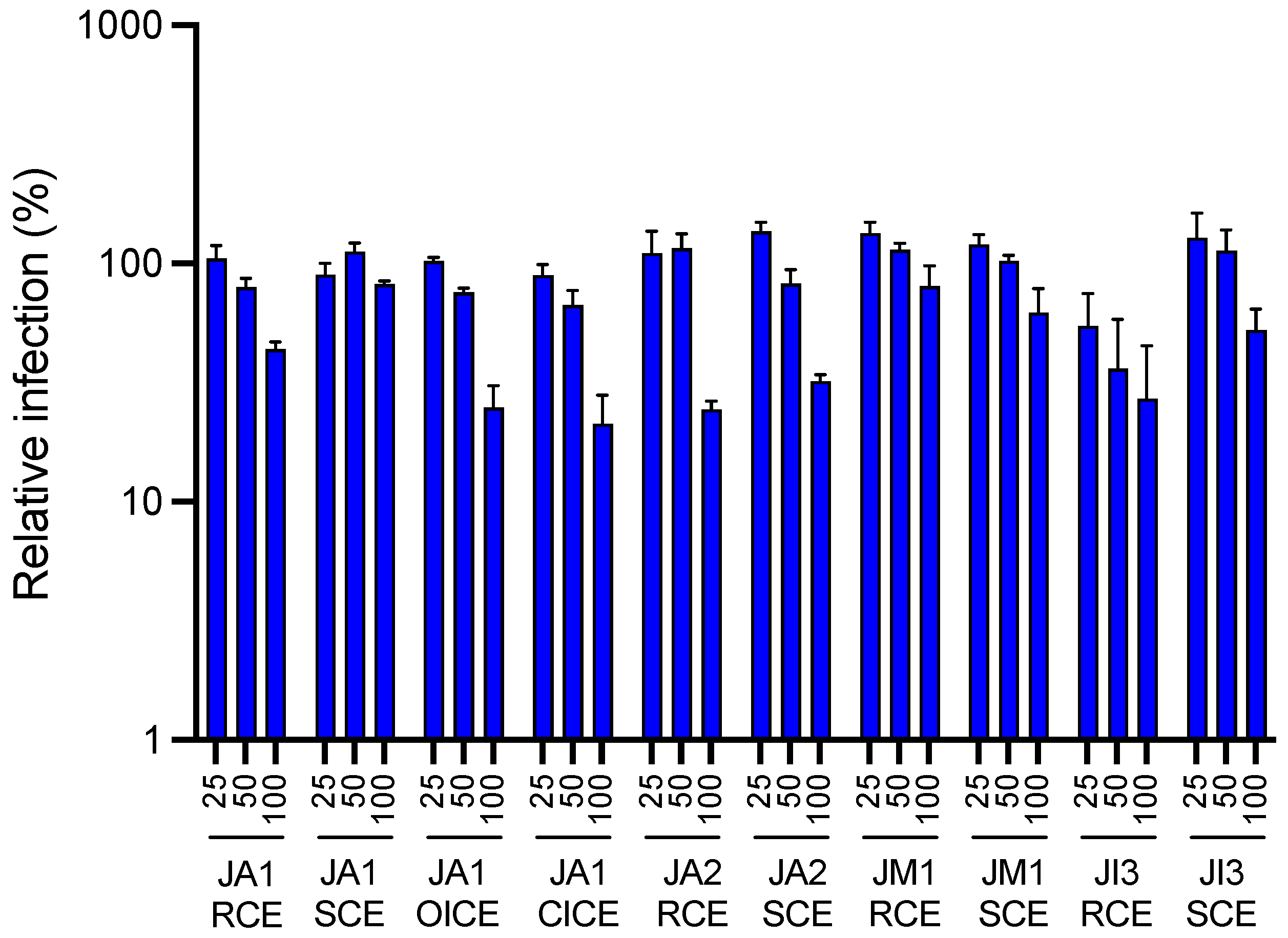
2.1. Antiviral Activity of Different Juncus Species against HCoV-229E
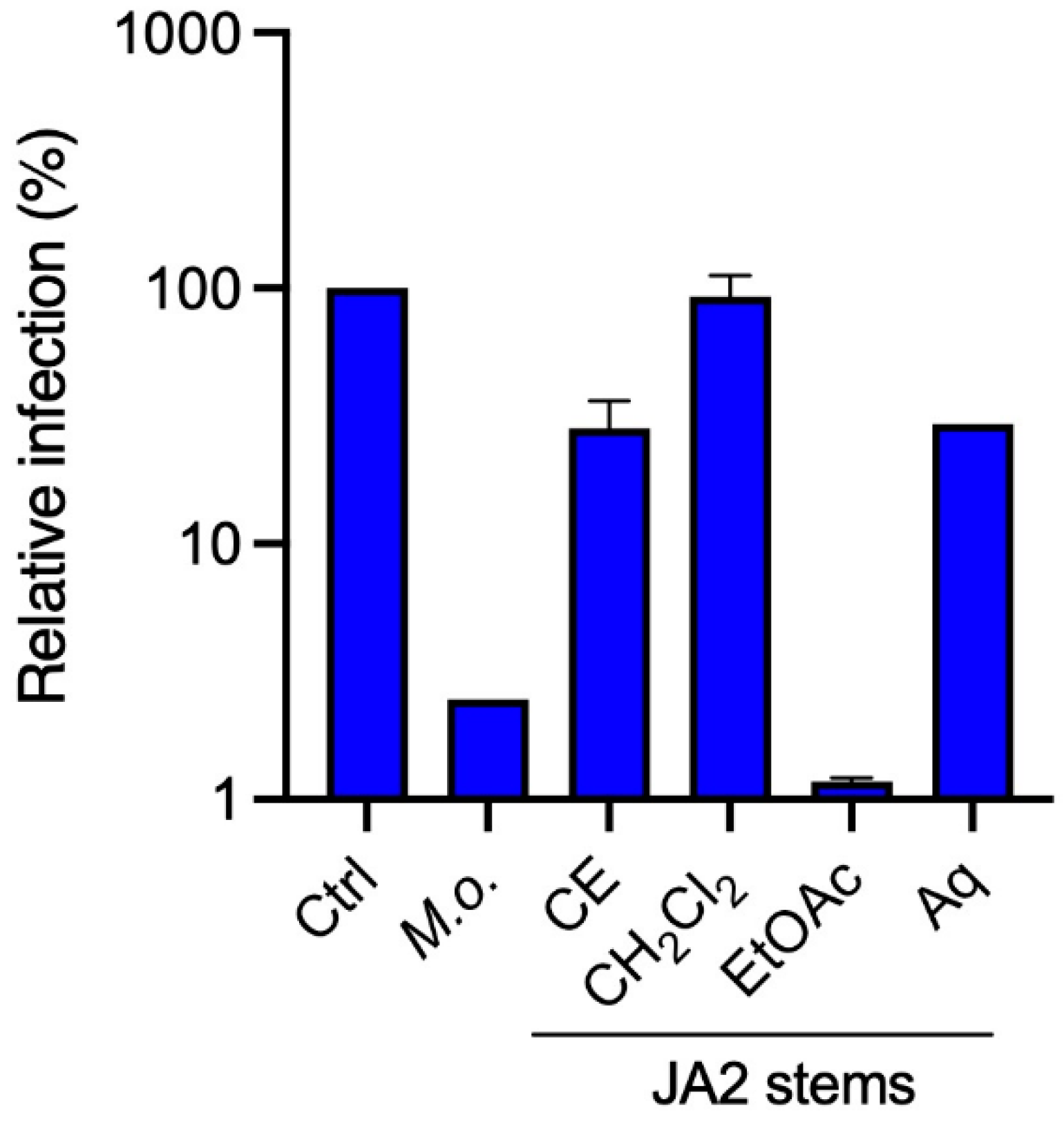
2.2. Antiviral Activity of Sub-Extracts of Juncus acutus Stems (JA2) against HCoV-229E
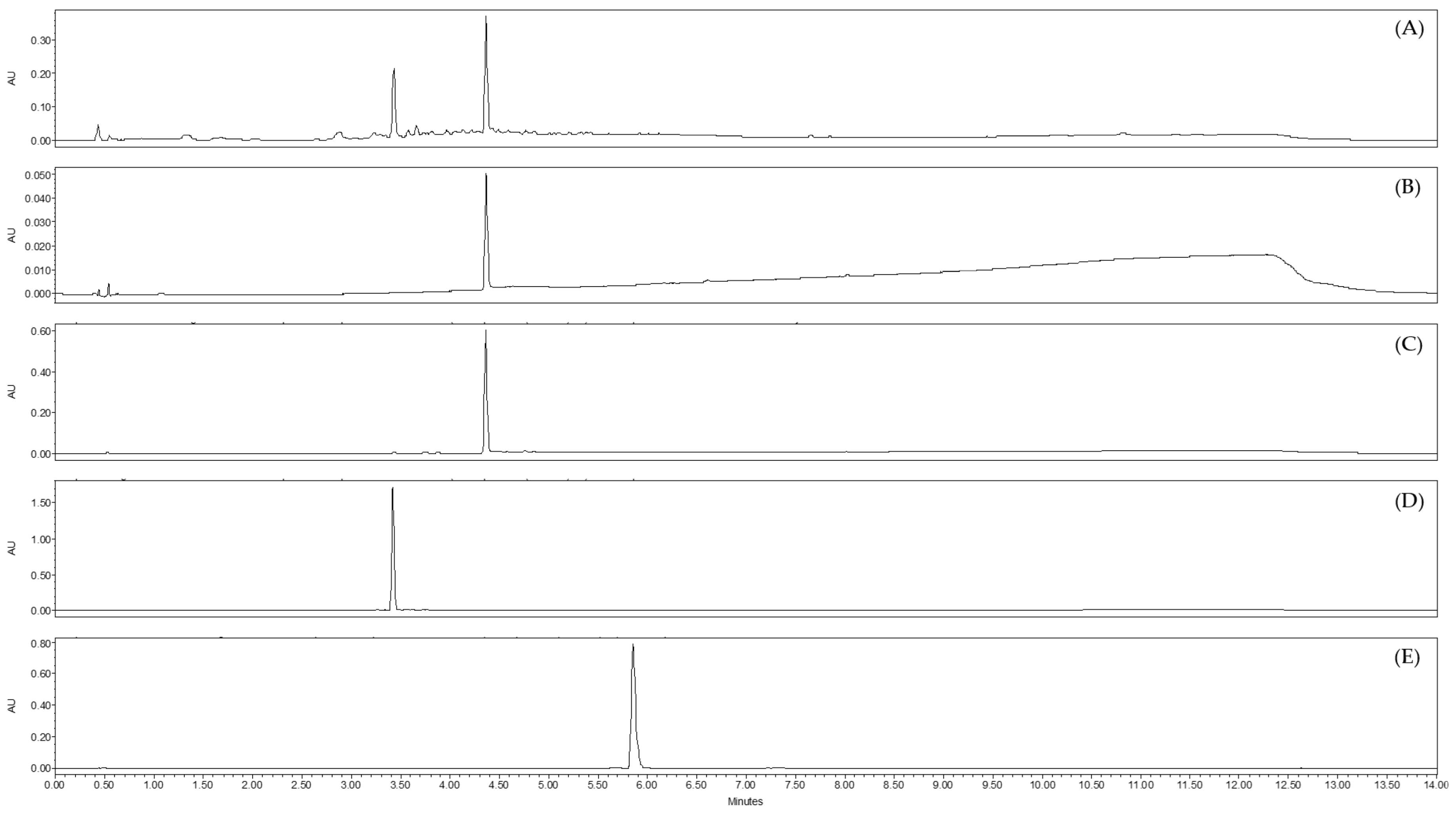
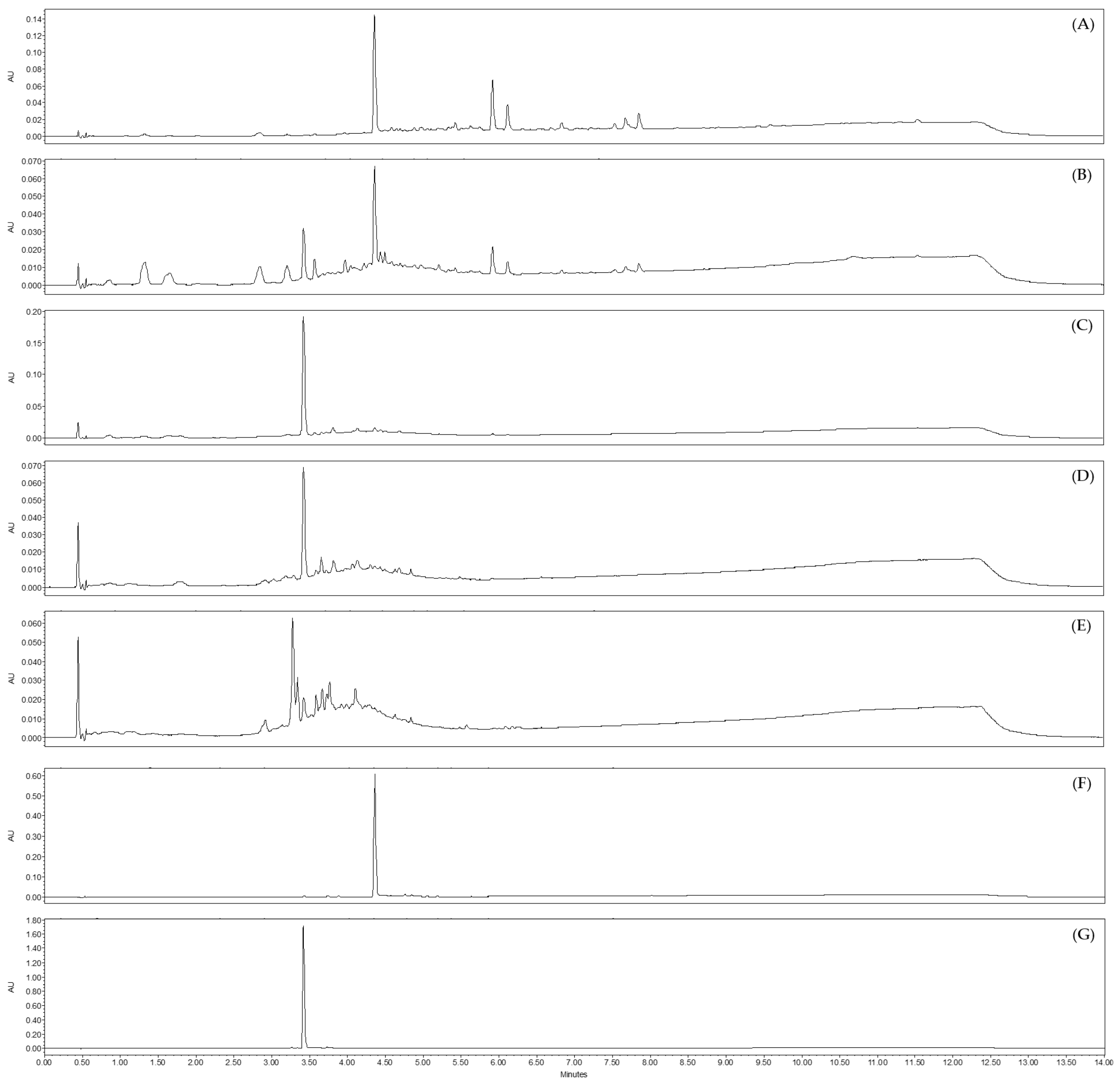
2.3. Analysis by UHPLC-UV-MS of Sub-Extracts of Juncus acutus Stems (JA2)
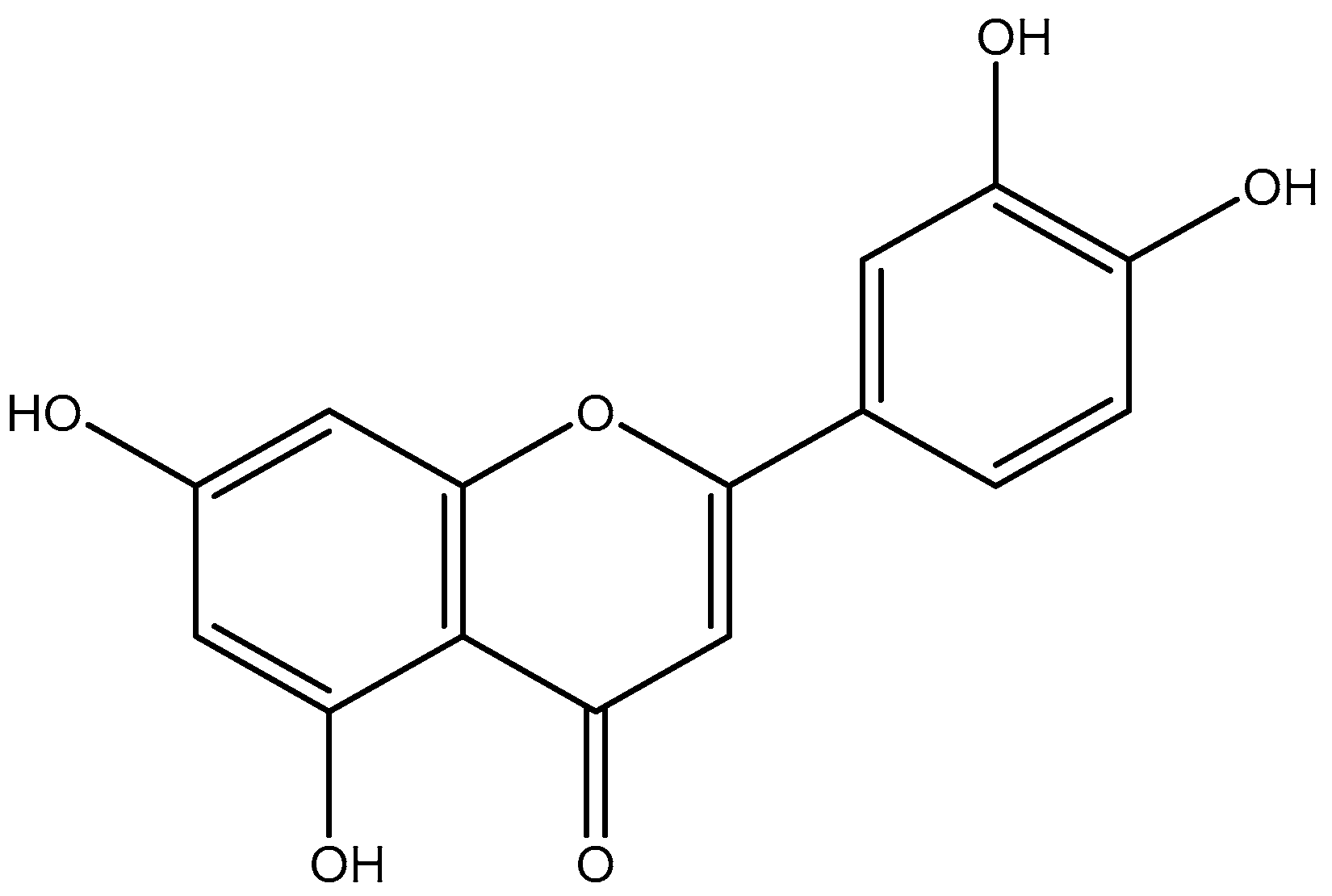
2.4. Identification of the Active Antiviral Compound(s) from the Ethyl Acetate and the Aqueous Sub-Extracts of Juncus acutus Stems (JA2) following Bioguided Fractionation
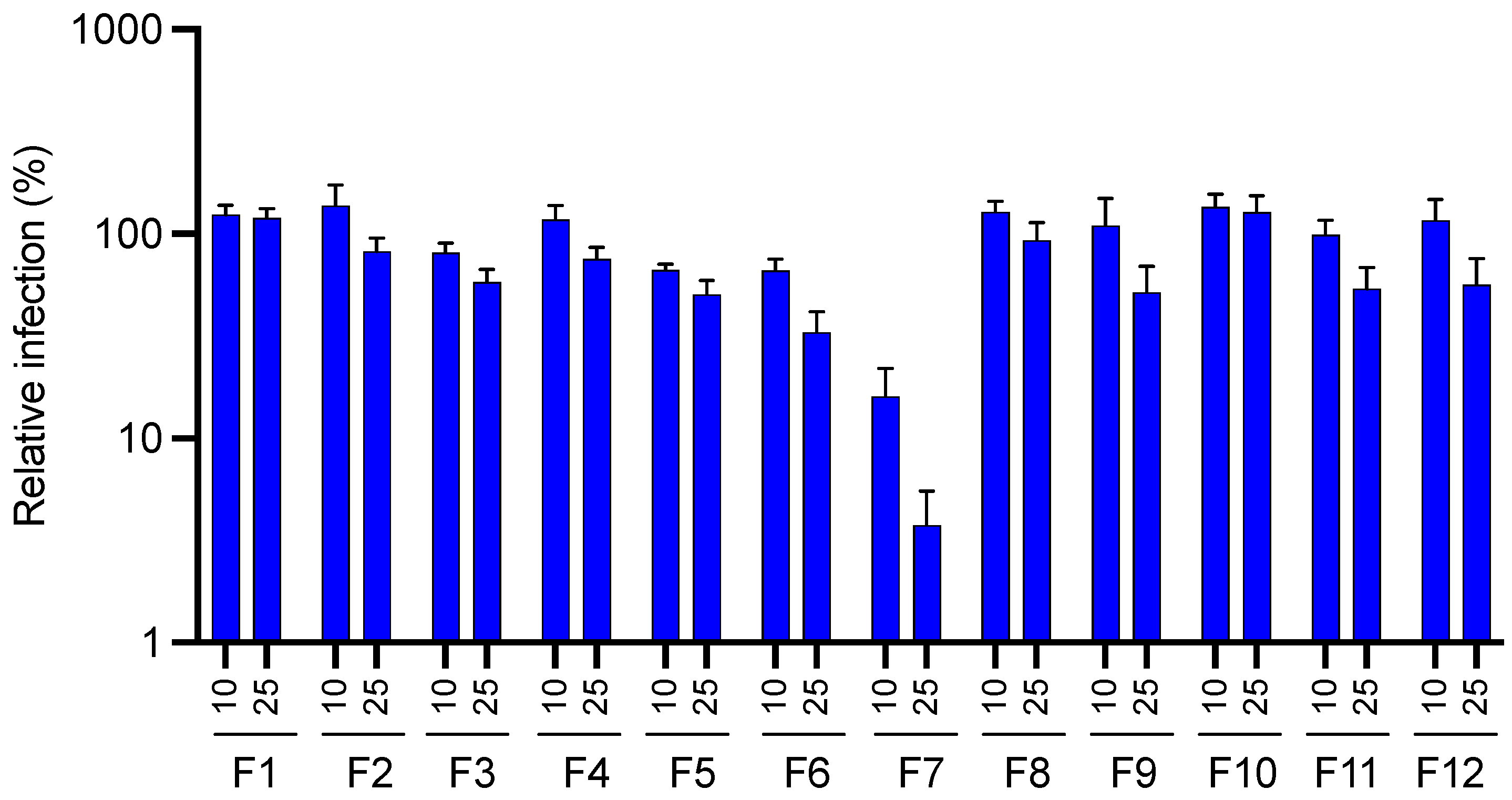
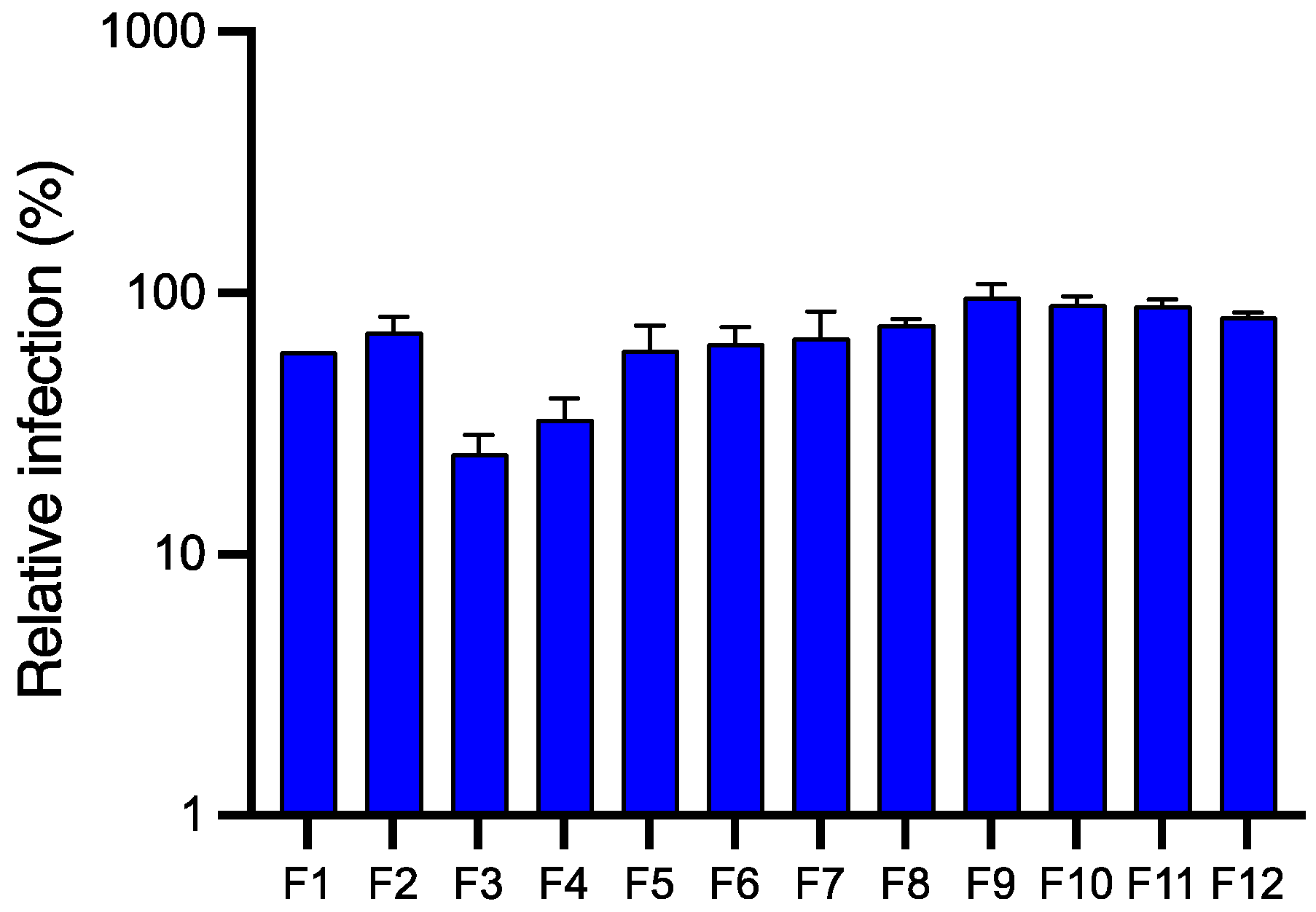
2.4.1. Antiviral Activity of Fractions Obtained from JA2 EtOAc Sub-Extract by Semi-Preparative HPLC
2.4.2. Antiviral Activity of Fractions Obtained from JA2 Aqueous Sub-Extract by CPC
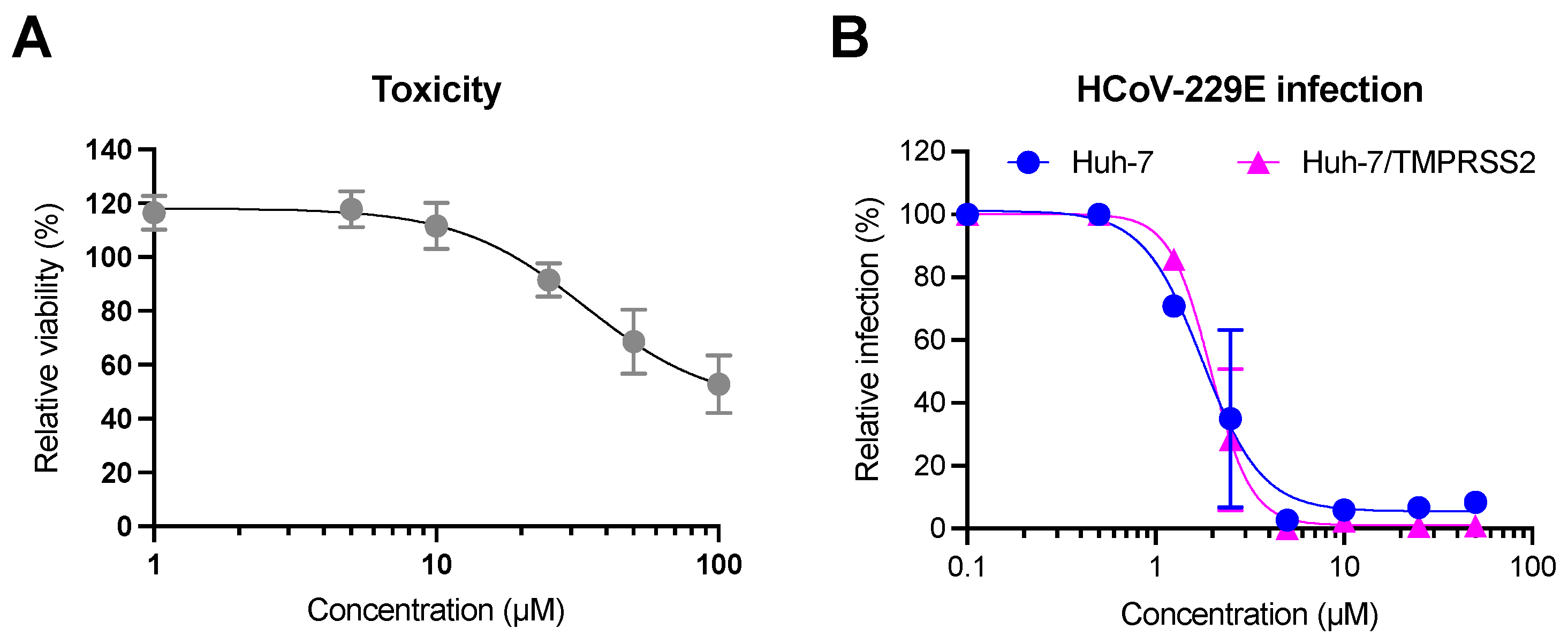
2.5. Antiviral Activity of Luteolin against HCoV-229E and Toxicity in Huh-7 Cells
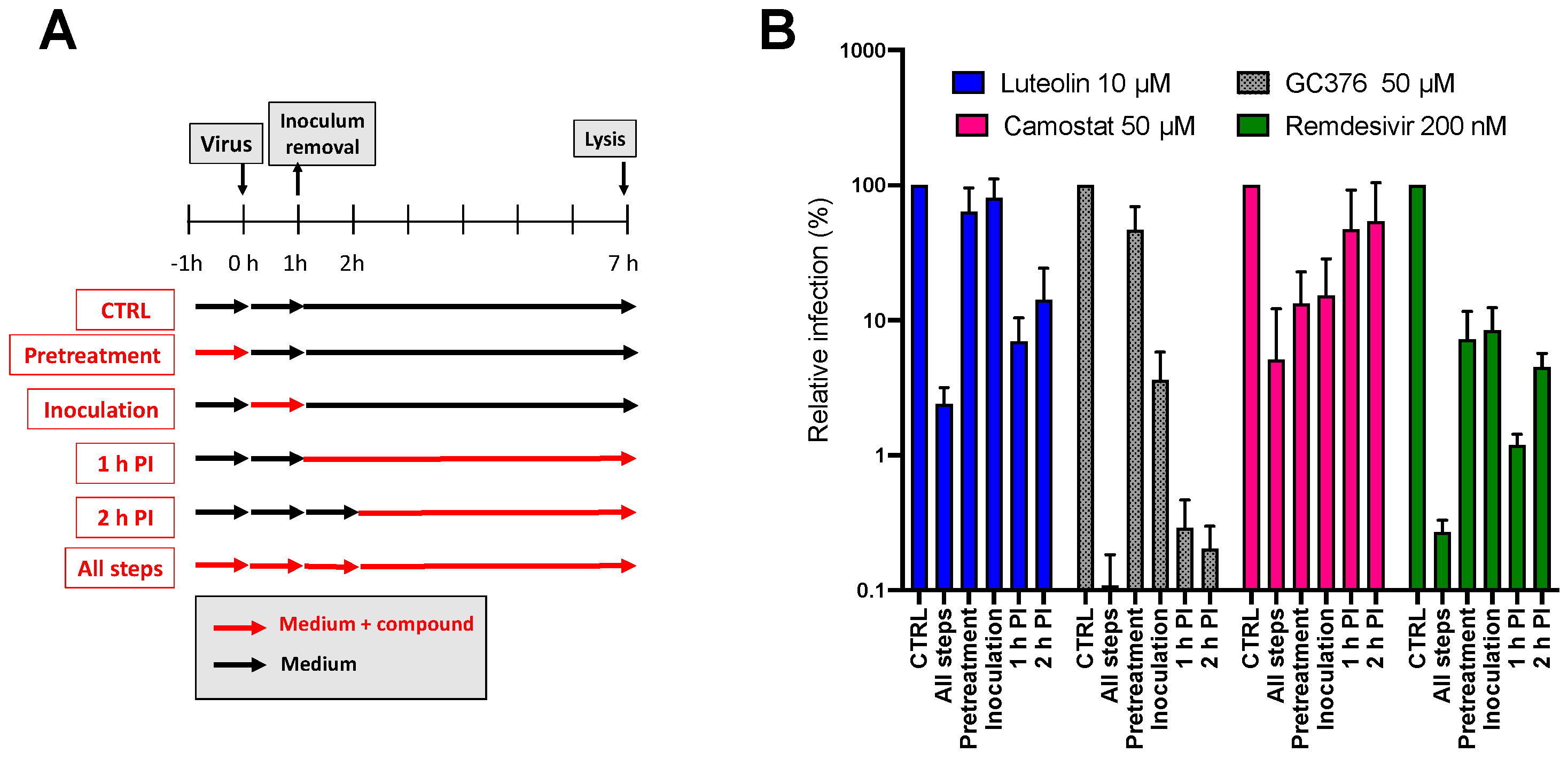
2.6. Mechanism of Action of Luteolin against HCoV-229E
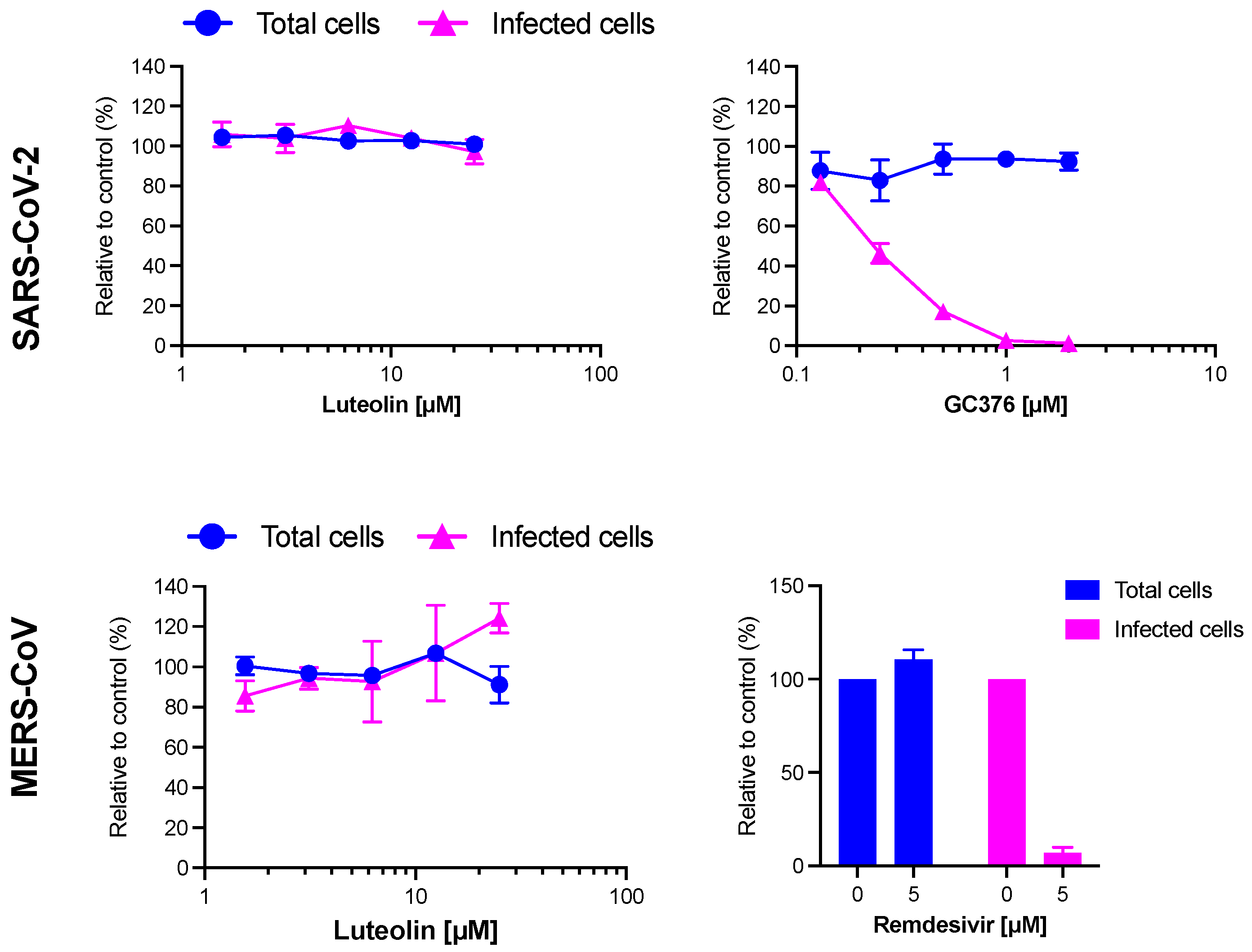
2.7. Antiviral Activity of Luteolin against SARS-CoV-2 and MERS-CoV
3. Discussion
4. Materials and Methods
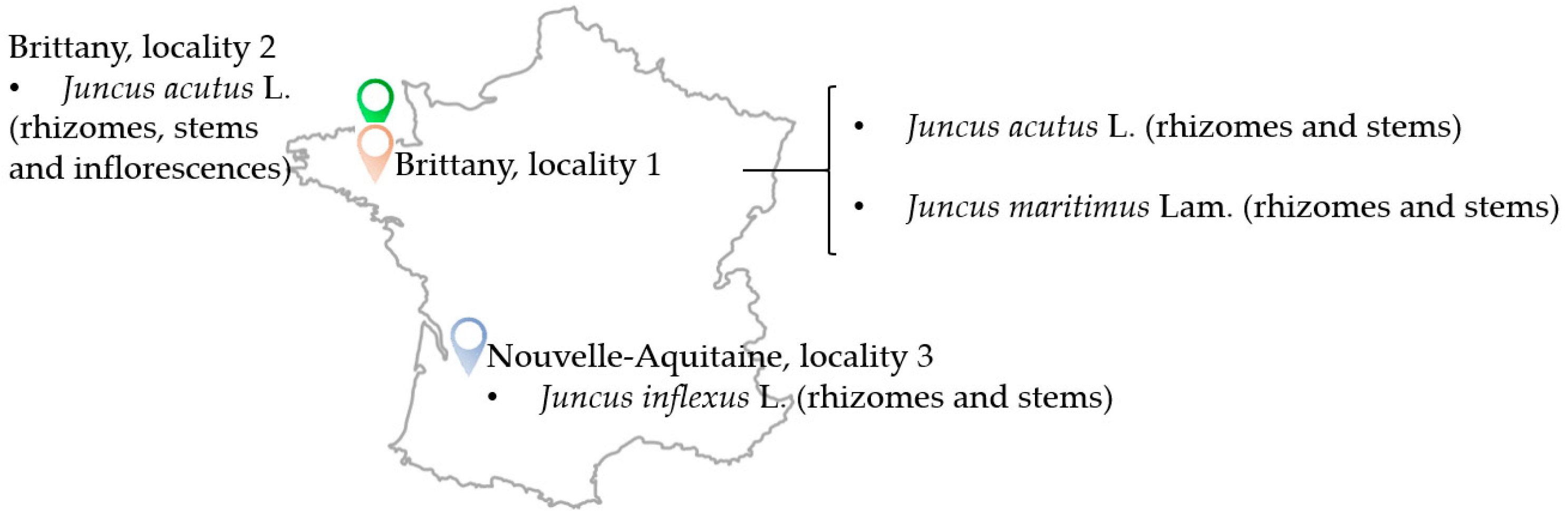
4.1. Plant Material
4.2. Plant Extract Preparation
4.3. Solvent-Solvent Partition of Juncus acutus Stems (JA2 SCE)
4.4. Fractionation of the EtOAc Sub-Extract of Juncus acutus Stems
4.5. Fractionation of the Aqueous Sub-Extract of Juncus acutus Stems
4.6. UHPLC-UV-MS Analysis
4.7. Cells and Culture Conditions
4.8. Viruses
4.9. Cell Toxicity Assay
4.10. Antiviral Activity Assay on HCoV-229E
4.11. SARS-CoV-2 Infection Assay
4.12. MERS-CoV Infection Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Sa, T.; Syed, N. The Future of Humanity and Microbes: Impact of Emerging Infectious Diseases on Global Health and Economies. IJSR 2013, 4, 2427–2442. [Google Scholar]
- Masters, P.S.; Perlman, S. Coronaviridae. Fields Virol. 2013, 1, 825–858. [Google Scholar]
- WHO. Report of the Twenty-First Meeting of Ministers of Health of Countries of the South-East Asia Region, New Delhi, India, 8–9 September 2003; WHO Regional Office for South-East Asia: New Delhi, India, 2003. [Google Scholar]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and Influenza Pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef]
- WHO. WHO Coronavirus (COVID-19) Dashboard 2023. Available online: https://covid19.who.int/ (accessed on 21 May 2023).
- Hamre, D.; Procknow, J.J. A New Virus Isolated from the Human Respiratory Tract. Exp. Biol. Med. 1966, 121, 190–193. [Google Scholar] [CrossRef]
- Farsani, S.M.J.; Dijkman, R.; Jebbink, M.F.; Goossens, H.; Ieven, M.; Deijs, M.; Molenkamp, R.; van der Hoek, L. The First Complete Genome Sequences of Clinical Isolates of Human Coronavirus 229E. Virus Genes 2012, 45, 433–439. [Google Scholar] [CrossRef]
- Corman, V.M.; Eckerle, I.; Memish, Z.A.; Liljander, A.M.; Dijkman, R.; Jonsdottir, H.; Juma Ngeiywa, K.J.Z.; Kamau, E.; Younan, M.; Al Masri, M.; et al. Link of a Ubiquitous Human Coronavirus to Dromedary Camels. Proc. Natl. Acad. Sci. USA 2016, 113, 9864–9869. [Google Scholar] [CrossRef]
- Sabir, J.S.M.; Lam, T.T.-Y.; Ahmed, M.M.M.; Li, L.; Shen, Y.; Abo-Aba, S.E.M.; Qureshi, M.I.; Abu-Zeid, M.; Zhang, Y.; Khiyami, M.A.; et al. Co-Circulation of Three Camel Coronavirus Species and Recombination of MERS-CoVs in Saudi Arabia. Science 2016, 351, 81–84. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Lee, J.-H.; Jung, Y.; Lee, S.-K.; Kim, J.; Lee, C.-S.; Kim, S.; Lee, J.-S.; Kim, N.-H.; Kim, H.-G. Rapid Biosensor of SARS-CoV-2 Using Specific Monoclonal Antibodies Recognizing Conserved Nucleocapsid Protein Epitopes. Viruses 2022, 14, 255. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, N.; Yang, L.; Song, X. Bioactive Natural Products in COVID-19 Therapy. Front. Pharmacol. 2022, 13, 926507. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, R.; Chapagain, P.; Siltberg-Liberles, J. Potential RNA-Dependent RNA Polymerase Inhibitors as Prospective Therapeutics against SARS-CoV-2. J. Med. Microbiol. 2020, 69, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.; Michot, J.-M.; Danlos, F.-X.; Ribas, A.; Soria, J.-C. Repurposing of Anticancer Drugs Expands Possibilities for Antiviral and Anti-Inflammatory Discovery in COVID-19. Cancer Discov. 2021, 11, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, L. Broad-spectrum Prodrugs with Anti-SARS-CoV-2 Activities: Strategies, Benefits, and Challenges. J. Med. Virol. 2022, 94, 1373–1390. [Google Scholar] [CrossRef]
- Deb, P.; Molla, M.M.A.; Saif-Ur-Rahman, K.M. An Update to Monoclonal Antibody as Therapeutic Option against COVID-19. Biosaf. Health 2021, 3, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Plichta, J.; Kuna, P.; Panek, M. Monoclonal Antibodies as Potential COVID-19 Therapeutic Agents. COVID 2022, 2, 599–620. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of Molnupiravir-Induced SARS-CoV-2 Mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- WHO. Solidarity Trial Consortium Remdesivir and Three Other Drugs for Hospitalised Patients with COVID-19: Final Results of the WHO Solidarity Randomised Trial and Updated Meta-Analyses. Lancet 2022, 399, 1941–1953. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Lee, S.T.; Gardner, D.R.; Panter, K.E.; James, L.F. Phytochemicals: The Good, the Bad and the Ugly? Phytochemistry 2007, 68, 2973–2985. [Google Scholar] [CrossRef] [PubMed]
- Mani, J.S.; Johnson, J.B.; Steel, J.C.; Broszczak, D.A.; Neilsen, P.M.; Walsh, K.B.; Naiker, M. Natural Product-Derived Phytochemicals as Potential Agents against Coronaviruses: A Review. Virus Res. 2020, 284, 197989. [Google Scholar] [CrossRef] [PubMed]
- Elmorsy, M.A.; El-Baz, A.M.; Mohamed, N.H.; Almeer, R.; Abdel-Daim, M.M.; Yahya, G. In Silico Screening of Potent Inhibitors against COVID-19 Key Targets from a Library of FDA-Approved Drugs. Environ. Sci. Pollut. Res. 2022, 29, 12336–12346. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a Database of Salt-Tolerant Plants: Helping put Halophytes to Work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef] [PubMed]
- Chapman, V.J. The New Perspective in the Halophytes. Q. Rev. Biol. 1942, 17, 291–311. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The Evolution of Halophytes, Glycophytes and Crops, and Its Implications for Food Security under Saline Conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Merchaoui, H.; Ksouri, R.; Abdelly, C.; Hanana, M. Halophytes.Tn: An Innovative Database for Tunisian Halophyte Plant Identification, Distribution and Characterization. Database 2022, 2022, baab082. [Google Scholar] [CrossRef]
- Sahli, R.; Rivière, C.; Neut, C.; Bero, J.; Sahuc, M.-E.; Smaoui, A.; Beaufay, C.; Roumy, V.; Hennebelle, T.; Rouillé, Y.; et al. An Ecological Approach to Discover New Bioactive Extracts and Products: The Case of Extremophile Plants. J. Pharm. Pharmacol. 2017, 69, 1041–1055. [Google Scholar] [CrossRef]
- Sahuc, M.-E.; Sahli, R.; Rivière, C.; Pène, V.; Lavie, M.; Vandeputte, A.; Brodin, P.; Rosenberg, A.R.; Dubuisson, J.; Ksouri, R.; et al. Dehydrojuncusol, a Natural Phenanthrene Compound Extracted from Juncus Maritimus, Is a New Inhibitor of Hepatitis C Virus RNA Replication. J. Virol. 2019, 93, e02009-18. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nagata, N.; Shirato, K.; Kawase, M.; Takeda, M. Taguchi Efficient Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein by the Transmembrane Protease TMPRSS2. J. Virol. 2010, 84, 12658–12664. [Google Scholar] [CrossRef]
- Shulla, A.; Heald-Sargent, T.; Subramanya, G.; Zhao, J.; Perlman, S.; Gallagher, T. A Transmembrane Serine Protease Is Linked to the Severe Acute Respiratory Syndrome Coronavirus Receptor and Activates Virus Entry. J. Virol. 2011, 85, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.; Uckeley, Z.M.; Doldan, P.; Stanifer, M.; Boulant, S.; Lozach, P.-Y. TMPRSS2 Expression Dictates the Entry Route Used by SARS-CoV-2 to Infect Host Cells. EMBO J. 2021, 40, e107821. [Google Scholar] [CrossRef] [PubMed]
- Meunier, T.; Desmarets, L.; Bordage, S.; Bamba, M.; Hervouet, K.; Rouillé, Y.; François, N.; Decossas, M.; Sencio, V.; Trottein, F.; et al. A Photoactivable Natural Product with Broad Antiviral Activity against Enveloped Viruses, Including Highly Pathogenic Coronaviruses. Antimicrob. Agents Chemother. 2022, 66, e01581-21. [Google Scholar] [CrossRef] [PubMed]
- El-Shamy, A.I.; Abdel-Razek, A.F.; Nassar, M.I. Phytochemical Review of Juncus L. Genus (Fam. Juncaceae). Arab. J. Chem. 2015, 8, 614–623. [Google Scholar] [CrossRef]
- Behery, F.A.A.; Naeem, Z.-E.M.; Maatooq, G.T.; Amer, M.M.A.; Ahmed, A.F. A Novel Antioxidant Phenanthrenoid Dimer from Juncus acutus L. Nat. Prod. Res. 2013, 27, 155–163. [Google Scholar] [CrossRef]
- Moustafa, S.; El-Alfy, T.; Hasan, N.; Abou El-Seoud, K. Inhibitory Effect of Flavonoids Isolated from Some Juncus Species (Flowers & Callus) on Blood Platelets Aggregation. Bull. Pharm. Sci. Assiut 2002, 25, 43–52. [Google Scholar] [CrossRef]
- Bús, C.; Tóth, B.; Stefkó, D.; Hohmann, J.; Vasas, A. Family Juncaceae: Promising Source of Biologically Active Natural Phenanthrenes. Phytochem. Rev. 2018, 17, 833–851. [Google Scholar] [CrossRef]
- Kovács, A.; Vasas, A.; Hohmann, J. Natural Phenanthrenes and Their Biological Activity. Phytochemistry 2008, 69, 1084–1110. [Google Scholar] [CrossRef]
- Murgolo, N.; Therien, A.G.; Howell, B.; Klein, D.; Koeplinger, K.; Lieberman, L.A.; Adam, G.C.; Flynn, J.; McKenna, P.; Swaminathan, G.; et al. SARS-CoV-2 Tropism, Entry, Replication, and Propagation: Considerations for Drug Discovery and Development. PLoS Pathog. 2021, 17, e1009225. [Google Scholar] [CrossRef]
- Chassagne, F.; Samarakoon, T.; Porras, G.; Lyles, J.T.; Dettweiler, M.; Marquez, L.; Salam, A.M.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. A Systematic Review of Plants with Antibacterial Activities: A Taxonomic and Phylogenetic Perspective. Front. Pharmacol. 2021, 11, 586548. [Google Scholar] [CrossRef]
- Vassilara, F.; Spyridaki, A.; Pothitos, G.; Deliveliotou, A.; Papadopoulos, A. A Rare Case of Human Coronavirus 229E Associated with Acute Respiratory Distress Syndrome in a Healthy Adult. Case Rep. Infect. Dis. 2018, 2018, 6796839. [Google Scholar] [CrossRef] [PubMed]
- Ishiuchi, K.; Kosuge, Y.; Hamagami, H.; Ozaki, M.; Ishige, K.; Ito, Y.; Kitanaka, S. Chemical Constituents Isolated from Juncus effusus Induce Cytotoxicity in HT22 Cells. J. Nat. Med. 2015, 69, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.A.; Harborne, J.B. Luteolin and Daphnetin Derivatives in the Juncaceae and Their Systematic Significance. Biochem. Syst. Ecol. 1975, 3, 181–190. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potential for Cancer Prevention and Therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- López-Lázaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Wang, S.; Ling, Y.; Yao, Y.; Zheng, G.; Chen, W. Luteolin Inhibits Respiratory Syncytial Virus Replication by Regulating the MiR-155/SOCS1/STAT1 Signaling Pathway. Virol. J. 2020, 17, 187. [Google Scholar] [CrossRef]
- Yan, H.; Ma, L.; Wang, H.; Wu, S.; Huang, H.; Gu, Z.; Jiang, J.; Li, Y. Luteolin Decreases the Yield of Influenza A Virus in Vitro by Interfering with the Coat Protein I Complex Expression. J. Nat. Med. 2019, 73, 487–496. [Google Scholar] [CrossRef]
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Zhang, H.; Luo, H.; Zhu, L.; Jiang, P.; et al. Small Molecules Blocking the Entry of Severe Acute Respiratory Syndrome Coronavirus into Host Cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef]
- Manvar, D.; Mishra, M.; Kumar, S.; Pandey, V.N. Identification and Evaluation of Anti Hepatitis C Virus Phytochemicals from Eclipta alba. J. Ethnopharmacol. 2012, 144, 545–554. [Google Scholar] [CrossRef]
- Bai, L.; Nong, Y.; Shi, Y.; Liu, M.; Yan, L.; Shang, J.; Huang, F.; Lin, Y.; Tang, H. Luteolin Inhibits Hepatitis B Virus Replication through Extracellular Signal-Regulated Kinase-Mediated Down-Regulation of Hepatocyte Nuclear Factor 4α Expression. Mol. Pharm. 2016, 13, 568–577. [Google Scholar] [CrossRef]
- Fan, W.; Qian, S.; Qian, P.; Li, X. Antiviral Activity of Luteolin against Japanese Encephalitis Virus. Virus Res. 2016, 220, 112–116. [Google Scholar] [CrossRef]
- Mehla, R.; Bivalkar-Mehla, S.; Chauhan, A. A Flavonoid, Luteolin, Cripples HIV-1 by Abrogation of Tat Function. PLoS ONE 2011, 6, e27915. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Su, W.; Jin, J.; Chen, J.; Li, X.; Zhang, X.; Sun, M.; Sun, S.; Fan, P.; An, D.; et al. Identification of Luteolin as Enterovirus 71 and Coxsackievirus A16 Inhibitors through Reporter Viruses and Cell Viability-Based Screening. Viruses 2014, 6, 2778–2795. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, W.; Perez-Lemus, G.R.; Menéndez, C.A.; Byléhn, F.; De Pablo, J.J. Molecular Characterization of COVID-19 Therapeutics: Luteolin as an Allosteric Modulator of the Spike Protein of SARS-CoV-2. Mol. Syst. Des. Eng. 2022, 7, 58–66. [Google Scholar] [CrossRef]
- Shawan, M.M.A.K.; Halder, S.K.; Hasan, M.d.A. Luteolin and Abyssinone II as Potential Inhibitors of SARS-CoV-2: An in Silico Molecular Modeling Approach in Battling the COVID-19 Outbreak. Bull. Natl. Res. Cent. 2021, 45, 27. [Google Scholar] [CrossRef]
- Munafò, F.; Donati, E.; Brindani, N.; Ottonello, G.; Armirotti, A.; De Vivo, M. Quercetin and Luteolin Are Single-Digit Micromolar Inhibitors of the SARS-CoV-2 RNA-Dependent RNA Polymerase. Sci. Rep. 2022, 12, 10571. [Google Scholar] [CrossRef]
- Sacan, A.; Ekins, S.; Kortagere, S. Applications and Limitations of In Silico Models in Drug Discovery. In Bioinformatics and Drug Discovery; Larson, R.S., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 910, pp. 87–124. ISBN 978-1-61779-964-8. [Google Scholar]
- Yasuda, M.T.; Fujita, K.; Hosoya, T.; Imai, S.; Shimoi, K. Absorption and Metabolism of Luteolin and Its Glycosides from the Extract of Chrysanthemum Morifolium Flowers in Rats and Caco-2 Cells. J. Agric. Food Chem. 2015, 63, 7693–7699. [Google Scholar] [CrossRef]
- Ali, F.; Siddique, Y.H. Bioavailability and Pharmaco-Therapeutic Potential of Luteolin in Overcoming Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets 2019, 18, 352–365. [Google Scholar] [CrossRef]
- Joel Mjiqiza, S.; Abraham Syce, J.; Chibuzo Obikeze, K. Pulmonary Effects and Disposition of Luteolin and Artemisia Afra Extracts in Isolated Perfused Lungs. J. Ethnopharmacol. 2013, 149, 648–655. [Google Scholar] [CrossRef]
- D’Ascanio, L.; Vitelli, F.; Cingolani, C.; Maranzano, M.; Brenner, M.J.; Di Stadio, A. Randomized Clinical Trial “Olfactory Dysfunction after COVID-19: Olfactory Rehabilitation Therapy vs. Intervention Treatment with Palmitoylethanolamide and Luteolin”: Preliminary Results. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4156–4162. [Google Scholar] [CrossRef]
- Aziz, N.; Kim, M.-Y.; Cho, J.Y. Anti-Inflammatory Effects of Luteolin: A Review of in Vitro, in Vivo, and in Silico Studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef] [PubMed]
- van den Worm, S.H.E.; Eriksson, K.K.; Zevenhoven, J.C.; Weber, F.; Züst, R.; Kuri, T.; Dijkman, R.; Chang, G.; Siddell, S.G.; Snijder, E.J.; et al. Reverse Genetics of SARS-Related Coronavirus Using Vaccinia Virus-Based Recombination. PLoS ONE 2012, 7, e32857. [Google Scholar] [CrossRef]
- Desmarets, L.; Callens, N.; Hoffmann, E.; Danneels, A.; Lavie, M.; Couturier, C.; Dubuisson, J.; Belouzard, S.; Rouillé, Y. A Reporter Cell Line for the Automated Quantification of SARS-CoV-2 Infection in Living Cells. Front. Microbiol. 2022, 13, 1031204. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Z. Natural Products, Alone or in Combination with FDA-Approved Drugs, to Treat COVID-19 and Lung Cancer. Biomedicines 2021, 9, 689. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Plant Part | Region | Geographical Coordinates |
|---|---|---|---|
| Juncus acutus L. | rhizomes, stems, inflorescences | Brittany, locality 1 | 48°48′44″ North, 3°05′27″ West |
| Juncus acutus L. | rhizomes, stems | Brittany, locality 2 | 48°49′48″ North, 3°04′40″ West |
| Juncus inflexus L. | rhizomes, stems | Nouvelle-Aquitaine, locality 3 | 44°59′44″ North, 0°26′41″ West |
| Juncus maritimus Lam | rhizomes, stems | Brittany locality 1 | 48°48′44″ North, 3°05′27″ West |
| Species | Plant Part | Abbreviation | Yield (%) |
|---|---|---|---|
| J. acutus (locality 1) | rhizomes | JA1 RCE | 9.6 |
| stems | JA1 SCE | 12.3 | |
| open inflorescences | JA1 OICE | 2.7 | |
| closed inflorescences | JA1 CICE | 8.5 | |
| J. acutus (locality 2) | stems | JA2 SCE | 11.85 |
| rhizomes | JA2 RCE | 6.95 | |
| J. maritimus (locality 1) | rhizomes | JM1 RCE | 11.25 |
| stems | JM1 SCE | 11.6 | |
| J. inflexus (locality 3) | rhizomes | JI3 RCE | 5.2 |
| stems | JI3 SCE | 11.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hakem, A.; Desmarets, L.; Sahli, R.; Malek, R.B.; Camuzet, C.; François, N.; Lefèvre, G.; Samaillie, J.; Moureu, S.; Sahpaz, S.; et al. Luteolin Isolated from Juncus acutus L., a Potential Remedy for Human Coronavirus 229E. Molecules 2023, 28, 4263. https://doi.org/10.3390/molecules28114263
Hakem A, Desmarets L, Sahli R, Malek RB, Camuzet C, François N, Lefèvre G, Samaillie J, Moureu S, Sahpaz S, et al. Luteolin Isolated from Juncus acutus L., a Potential Remedy for Human Coronavirus 229E. Molecules. 2023; 28(11):4263. https://doi.org/10.3390/molecules28114263
Chicago/Turabian StyleHakem, Asma, Lowiese Desmarets, Ramla Sahli, Rawen Ben Malek, Charline Camuzet, Nathan François, Gabriel Lefèvre, Jennifer Samaillie, Sophie Moureu, Sevser Sahpaz, and et al. 2023. "Luteolin Isolated from Juncus acutus L., a Potential Remedy for Human Coronavirus 229E" Molecules 28, no. 11: 4263. https://doi.org/10.3390/molecules28114263
APA StyleHakem, A., Desmarets, L., Sahli, R., Malek, R. B., Camuzet, C., François, N., Lefèvre, G., Samaillie, J., Moureu, S., Sahpaz, S., Belouzard, S., Ksouri, R., Séron, K., & Rivière, C. (2023). Luteolin Isolated from Juncus acutus L., a Potential Remedy for Human Coronavirus 229E. Molecules, 28(11), 4263. https://doi.org/10.3390/molecules28114263