
Effectiveness of E-Beam Radiation against Saccharomyces cerevisiae, Brettanomyces bruxellensis, and Wild Yeast and Their Influence on Wine Quality


 and
and
Abstract
:1. Introduction
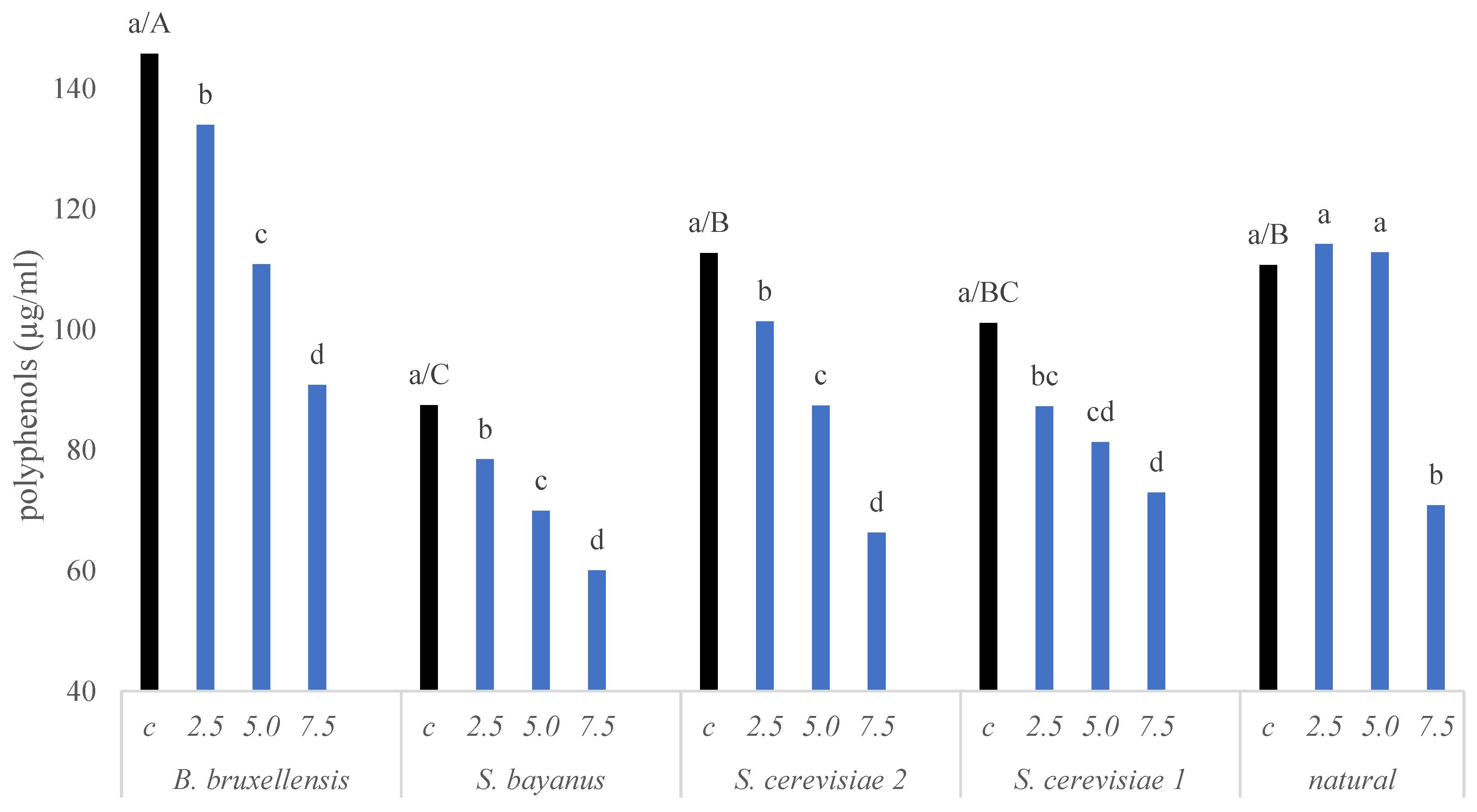
2. Results

2.1. Quality of Irradiated Wine—Selected Parameters



2.2. Influence of Yeast Bread on Color, Polyphenol Content, and Selected Chemical Compounds Included in the Wine Aroma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Byproducts | B. bruxellensis | S. bayanus | S. cerevisiae 1 (ES 181) | S. cerevisiae 2 (ES 123) | Wild Yeasts |
|---|---|---|---|---|---|
| Isoamyl (mg/L) | 177 d | 126 c | 88 a | 97 b | 252 e |
| Iso-butanol (mg/L) | 40.1 e | 26.3 c | 23.0 b | 19.6 a | 35.5 d |
| N-propanol (mg/L) | 52.3 c | 28.9 b | 18.7 a | 16.2 a | 78.6 d |
| Total | 269.4 C | 181.2 B | 129.7 A | 132.8 A | 366.1 D |
| Glycerol (g/L) | 4.53 d | 4.28 c | 3.90 b | 2.81 a | 5.34 e |
| Ethyl acetate (mg/L) | 47.1 c | 37.8 b | 26.5 a | 22.9 a | 53.2 d |
| Isoamyl acetate (mg/L) | 0.18 c | 0.14 b | 0.13 ab | 0.11 a | 0.22 d |
| Isobutyl acetate (mg/L) | 0.55 cd | 0.49 bc | 0.68 de | 0.72 e | 0.24 a |
| Acetoine (mg/L) | 0.31 cd | 0.35 d | 0.12 a | 0.17 b | 0.27 c |
| Acetaldehyde (mg/L) | 67.4 d | 49.7 c | 33.5 b | 29.6 a | 63.0 d |
| 2 and 3-Methyl-butanol (mg/L) | 98.4 c | 78.4 b | 66.8 a | 72.5 ab | 112.2 d |


3. Discussion
3.1. Effects of Ionizing Radiation on Yeast and Polyphenols
3.2. Influence of Yeast on the Bouquet of Wine
4. Materials and Methods
4.1. Characteristics of the Area of Research and Plant Material
4.2. Description of the Variety and Production of Wine
4.3. Yeast: Assessment of Their Numbers in the Wine
4.4. Irradiation
4.5. Color Measurement
4.6. Identification of Compounds in Wine
4.6.1. Phenolics
4.6.2. Glycerol
4.6.3. Volatile Compound Analysis
4.7. Sensory Evaluation
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. (Eds.) Handbook of Enology, Volume 1: The Microbiology of Wine and Vinifications; John Wiley & Sons: Hoboken, NJ, USA, 2006; Volume 1, pp. 73–124. [Google Scholar]
- Drtilová, T.; Ďurčanská, K.; Machyňáková, A.; Špánik, I.; Klempová, T.; Furdíková, K. Impact of different pure cultures of Saccharomyces cerevisiae on the volatile profile of Cabernet Sauvignonrosé wines. Czech J. Food Sci. 2020, 38, 94–102. [Google Scholar] [CrossRef]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage activities of Dekkera/Brettanomyces spp. In Food Spoilage Microorganisms; Blackburn, C., Ed.; Woodhead: Cambridge, UK, 2006; pp. 354–398. [Google Scholar]
- Barata, A.; Caldeira, J.; Botellheiro, R.; Pagliara, D.; Malfeito-Ferreira, M.; Loureiro, V. Survival patterns of Dekkera bruxellensis in wines and inhibitory effect of sulphur dioxide. Int. J. Food Microbiol. 2008, 121, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Sun, F.; Wang, W.; Liu, Y.; Wang, J.; Sun, J.; Mu, J.; Gao, Z. Effects of spontaneous fermentation on the microorganisms diversity and volatile compounds during ‘Marselan’ from grape to wine. LWT-Food Sci. Technol. 2020, 134, 110193. [Google Scholar] [CrossRef]
- Francesca, N.; Sannino, C.; Settanni, L.; Corona, O.; Barone, E.; Moschetti, G. Microbiological and chemical monitoring of Marsala base wine obtained by spontaneous fermentation during large-scale production. Ann. Microbiol. 2014, 64, 1643–1657. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, M.; Cortés, S.; Santiago, J.L.; Martínez, C.; Fernández, E. Aromatic Compounds in Wines Produced During Fermentation: Effect of Three Red Cultivars. Int. J. Food Prop. 2007, 10, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Malfeito-Ferreira, M. Yeasts and wine off-flavours: A technological perspective. Ann. Microbiol. 2011, 61, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Błaszak, M.; Nowak, A.; Lachowicz, S.; Migdał, W.; Ochmian, I. E-Beam Irradiation and Ozonation as an Alternative to the Sulphuric Method of Wine Preservation. Molecules 2019, 24, 3406. [Google Scholar] [CrossRef] [Green Version]
- Ward, L.R.; Van Schaik, E.; Samuel, J.; Pillai, S.D. Reduction in microbial infection risks from raw milk by Electron Beam Technology. Radiat. Phys. Chem. 2020, 168, 108567. [Google Scholar] [CrossRef]
- Migdał, W.; Owczarczyk, H. Radiation decontamination of meat lyophylized products. Radiat. Phys. Chem. 2002, 63, 371–373. [Google Scholar] [CrossRef]
- Migdał, W.; Owczarczyk, H.; Kędzia, B.; Hołderna-Kędzia, E.; Madajczyk, D. Microbiological decontamination of natural honey by irradiation. Radiat. Phys. Chem. 2000, 57, 285–288. [Google Scholar] [CrossRef]
- Barkai-Golan, R.; Follett, P.A. Irradiation for Quality Improvement, Microbial Safety and Phytosanitation of Fresh Produce; Elsevier Science: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Smith, B.; Shayanfar, S.; Walzem, R.; Alvarado, C.Z.; Pillai, S.D. Preserving fresh fruit quality by low-dose electron beam processing for vending distribution channels. Radiat. Phys. Chem. 2020, 168, 108540. [Google Scholar] [CrossRef]
- De Bruyn, I. Prospects of Radiation Sterilization of Self-Stable Food. In Irradiation for Food Safety and Quality; Loaharanu, P., Thomas, P., Eds.; Technomic Publishing Co. Inc.: Lancaster, PA, USA, 2001. [Google Scholar]
- Chmielewski, A.G.; Migdal, W. Radiation decontamination of herbs and spices. Nukleonika 2005, 50, 179–184. [Google Scholar]
- CODEX STAN 106-1983, REV.1-2003. General Standard for Irradiated Foods. 2003. Available online: http://www.codexalimentarius.net/download/standards/16/CXS_106e.pdf (accessed on 1 October 2016).
- Ehlermann, D.A. The early history of food irradiation. Radiat. Phys. Chem. 2016, 129, 10–12. [Google Scholar] [CrossRef]
- Roberts, P.B. Food irradiation: Standards, regulations and world-wide trade. Radiat. Phys. Chem. 2016, 129, 30–34. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Edelson, S.G.; Snipes, K.; Boyd, G. Inactivation of Escherichia coli O157:H7 in Apple Juice by Irradiation. Appl. Environ. Microbiol. 1998, 64, 4533–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlikowski, L.; Migdał, W.; Ptaszek, M.; Gryczka, U. Effectiveness of electron beam irradiation in the control of some soil-borne pathogens. Nukleonika 2011, 56, 357–362. [Google Scholar]
- Nieto-Sandoval, J.M.; Almela, L.; Fernández-López, J.A.; Muñoz, J.A. Effect of Electron Beam Irradiation on Color and Microbial Bioburden of Red Paprika. J. Food Prot. 2000, 63, 633–637. [Google Scholar] [CrossRef]
- Gryczka, U.; Kameya, H.; Kimura, K.; Todoriki, S.; Migdał, W.; Bułka, S. Efficacy of low energy electron beam on microbial decontamination of spices. Radiat. Phys. Chem. 2020, 170, 108662. [Google Scholar] [CrossRef]
- Rio Segade, S.; Orriols, I.; Gerbi, V.; Rolle, L. Phenolic characterization of thirteen red grape cultivars from Galicia by anthocyanin profile and flavanol composition. J. Int. Sci. Vigne Vin 2009, 43, 189–198. [Google Scholar] [CrossRef]
- Morata, A.; Bañuelos, M.A.; Tesfaye, W.; Loira, I.; Palomero, F.; Benito, S.; Callejo, M.J.; Villa, A.; González, M.C.; Suárez-Lepe, J.A. Electron Beam Irradiation of Wine Grapes: Effect on Microbial Populations, Phenol Extraction and Wine Quality. Food Bioprocess Technol. 2015, 8, 1845–1853. [Google Scholar] [CrossRef]
- Breitfellner, F.; Solar, S.; Sontag, G. Effect of γ-Irradiation on Phenolic Acids in Strawberries. J. Food Sci. 2002, 67, 517–521. [Google Scholar] [CrossRef]
- Sajilata, M.; Singhal, R. Effect of irradiation and storage on the antioxidative activity of cashew nuts. Radiat. Phys. Chem. 2006, 75, 297–300. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in wine-making. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Wei, R.; Wang, L.; Ding, Y.; Zhang, L.; Gao, F.; Chen, N.; Song, Y.; Li, H.; Wang, H. Natural and sustainable wine: A review. Crit. Rev. Food Sci. Nutr. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- de-la-Fuente-Blanco, A.; Sáenz-Navajas, M.P.; Ferreira, V. Levels of higher alcohols inducing aroma changes and modulating experts’ preferences in wine model solutions. Aust. J. Grape Wine Res. 2017, 23, 162–169. [Google Scholar] [CrossRef]
- Cioch, M.; Skotniczny, M.; Kuchta, T.; Satora, P. Characteristics of selected parameters of grape wines obtained from the vineyards of southern Poland. Postępy Tech. Przetwórstwa Spożywczego 2018, 1, 14–18. [Google Scholar]
- Remize, F.; Sablayrolles, J.M.; Dequin, S. Re-assessment of the influence of yeast strain and environmental factors on glycerol production in wine. J. Appl. Microbiol. 2000, 88, 371–378. [Google Scholar] [CrossRef]
- Gawel, R.; VAN Sluyter, S.; Waters, E.J. The effects of ethanol and glycerol on the body and other sensory characteristics of Riesling wines. Aust. J. Grape Wine Res. 2007, 13, 38–45. [Google Scholar] [CrossRef]
- Yalcin, S.K.; Ozbas, Z.Y. Effects of pH and temperature on growth and glycerol production kinetics of two indigenous wine strains of Saccharomyces cerevisiae from Turkey. Braz. J. Microbiol. 2008, 39, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Zhang, Z. Volatile Compounds of Young Wines from Cabernet Sauvignon, Cabernet Gernischet and Chardonnay Varieties Grown in the Loess Plateau Region of China. Molecules 2010, 15, 9184–9196. [Google Scholar] [CrossRef] [Green Version]
- Wzorek, W.; Pogorzelski, E. Technology of Grape and Fruit Wine-Making; SIGMA—NOT: Warsaw, Poland, 1998; pp. 56–71. [Google Scholar]
- Cortés-Diéguez, S.; Rodriguez-Solana, R.; Domínguez, J.M.; Díaz, E. Impact odorants and sensory profile of young red wines. J. Inst. Brew. 2015, 121, 628–635. [Google Scholar] [CrossRef]
- Cucciniello, R.; Forino, M.; Picariello, L. How acetaldehyde reacts with low molecular weight phenolics in white and red wines. Eur. Food Res. Technol. 2021, 247, 2935–2944. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma-A review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to wine-making. Ann. Microbiol. 2011, 61, 25–32. [Google Scholar] [CrossRef]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Spitaels, F.; Wieme, A.; Janssens, M.; Aerts, M.; Daniel, H.-M.; Van Landschoot, A. The Microbial Diversity of Traditional Spontaneously Fermented Lambic Beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Canonico, L.; Marinelli, V.; Ciani, M.; Comitini, F. Occurrence of Brettanomyces bruxellensis on Grape Berries and in Related Winemaking Cellar. Front. Microbiol. 2019, 10, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [Green Version]
- Lachowicz, S.; Oszmiański, J.; Rapak, A.; Ochmian, I. Profile and content of phenolic compounds in leaves, flowers, roots, and stalks of Sanguisorba officinalis L. determined with the LC-DAD-ESI-QTOF-MS/MS analysis and their in vitro anti-oxidant, antidiabetic, antiproliferative potency. Pharmaceuticals 2020, 13, 191. [Google Scholar] [CrossRef] [PubMed]
- Sterczyńska, M.; Machowski, M.; Jakubowski, M.; Wiśniewski, A. Effectiveness of selected methods of clarification of grape red wines fermented with wild and noble yeast. Inżynieria Przetwórstwa Spożywczego 2015, 4, 28–33. [Google Scholar]
- Mijowska, K.; Ochmian, I.; Oszmiański, J. Rootstock effects on polyphenol content in grapes of ‘Regent’ cultivated under cool climate conditio. J. Appl. Bot. Food Qual. 2017, 90, 159–164. [Google Scholar] [CrossRef]
- Ochmian, I.; Oszmiański, J.; Lachowicz, S.; Krupa-Małkiewicz, M. Rootstock effect on physicochemical properties and content of bioactive compounds of four cultivars Cornelian cherry fruits. Sci. Hortic. 2019, 256, 108588. [Google Scholar] [CrossRef]
- ISO 21527-1:2008; Horizontal Method for the Enumeration of Yeasts and Molds. Part 1: Colony Count Technique in Products with Water Activity Greater than 0.95. International Organization for Standardization: Geneva, Switzerland, 2008. Available online: https://www.iso.org/obp/ui/#iso:std:iso:21527:-1:ed-1:v1:en (accessed on 27 March 2019).
- Oszmiański, J.; Kolniak-Ostek, J.; Lachowicz, S.; Gorzelany, J.; Matłok, N. Effect of dried powder preparation process on polyphenolic content and antioxidant capacity of cranberry (Vaccinium macrocarpon L.). Ind. Crop. Prod. 2015, 77, 658–665. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Błaszak, M.; Jakubowska, B.; Lachowicz-Wiśniewska, S.; Migdał, W.; Gryczka, U.; Ochmian, I. Effectiveness of E-Beam Radiation against Saccharomyces cerevisiae, Brettanomyces bruxellensis, and Wild Yeast and Their Influence on Wine Quality. Molecules 2023, 28, 4867. https://doi.org/10.3390/molecules28124867
Błaszak M, Jakubowska B, Lachowicz-Wiśniewska S, Migdał W, Gryczka U, Ochmian I. Effectiveness of E-Beam Radiation against Saccharomyces cerevisiae, Brettanomyces bruxellensis, and Wild Yeast and Their Influence on Wine Quality. Molecules. 2023; 28(12):4867. https://doi.org/10.3390/molecules28124867
Chicago/Turabian StyleBłaszak, Magdalena, Barbara Jakubowska, Sabina Lachowicz-Wiśniewska, Wojciech Migdał, Urszula Gryczka, and Ireneusz Ochmian. 2023. "Effectiveness of E-Beam Radiation against Saccharomyces cerevisiae, Brettanomyces bruxellensis, and Wild Yeast and Their Influence on Wine Quality" Molecules 28, no. 12: 4867. https://doi.org/10.3390/molecules28124867
APA StyleBłaszak, M., Jakubowska, B., Lachowicz-Wiśniewska, S., Migdał, W., Gryczka, U., & Ochmian, I. (2023). Effectiveness of E-Beam Radiation against Saccharomyces cerevisiae, Brettanomyces bruxellensis, and Wild Yeast and Their Influence on Wine Quality. Molecules, 28(12), 4867. https://doi.org/10.3390/molecules28124867