
Antioxidant and Antidiabetic Properties of Phlorotannins from Ascophyllum nodosum Seaweed Extracts



Abstract
:1. Introduction: The Importance of Macroalgae in the Food Industry
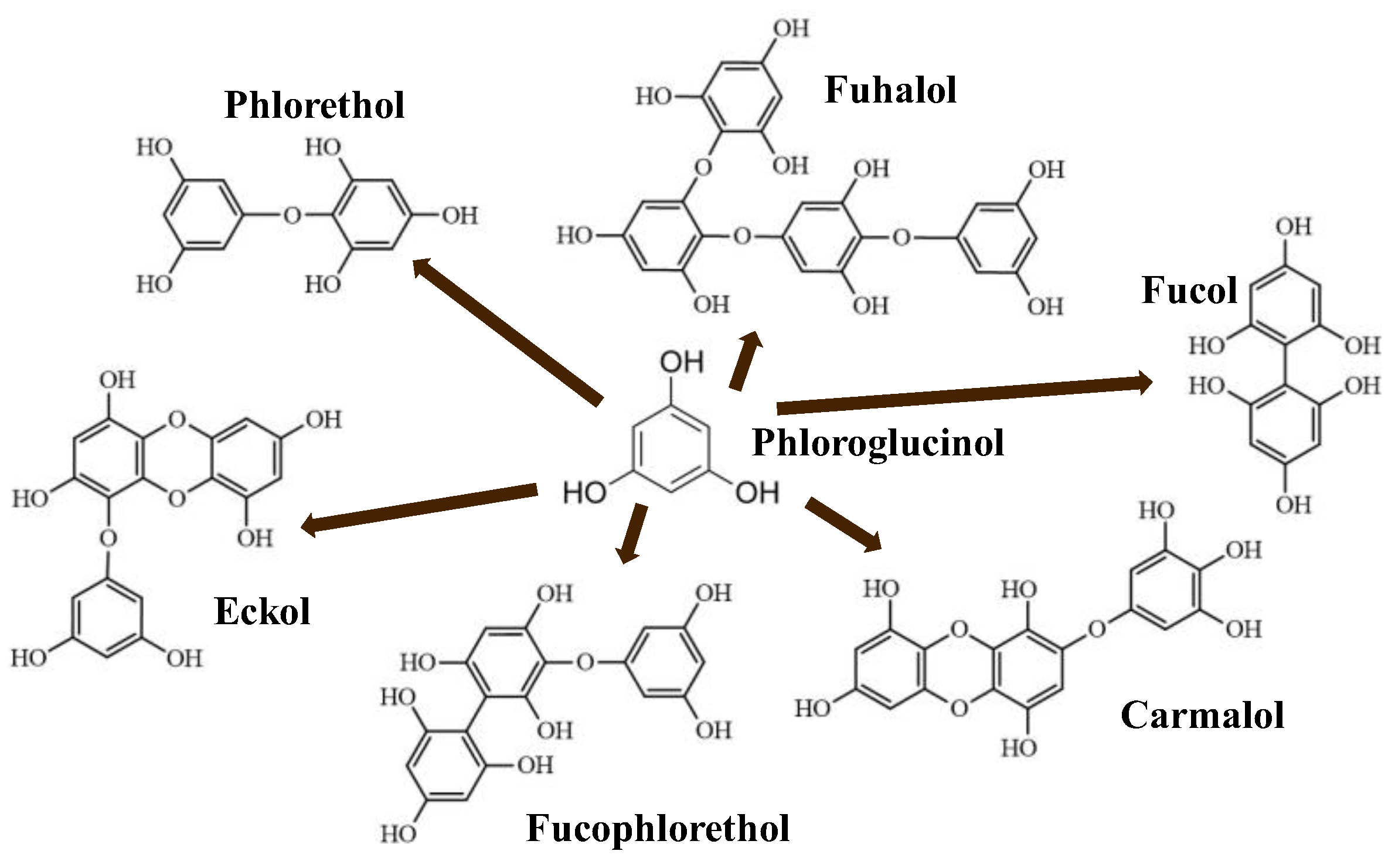
2. Phlorotannins
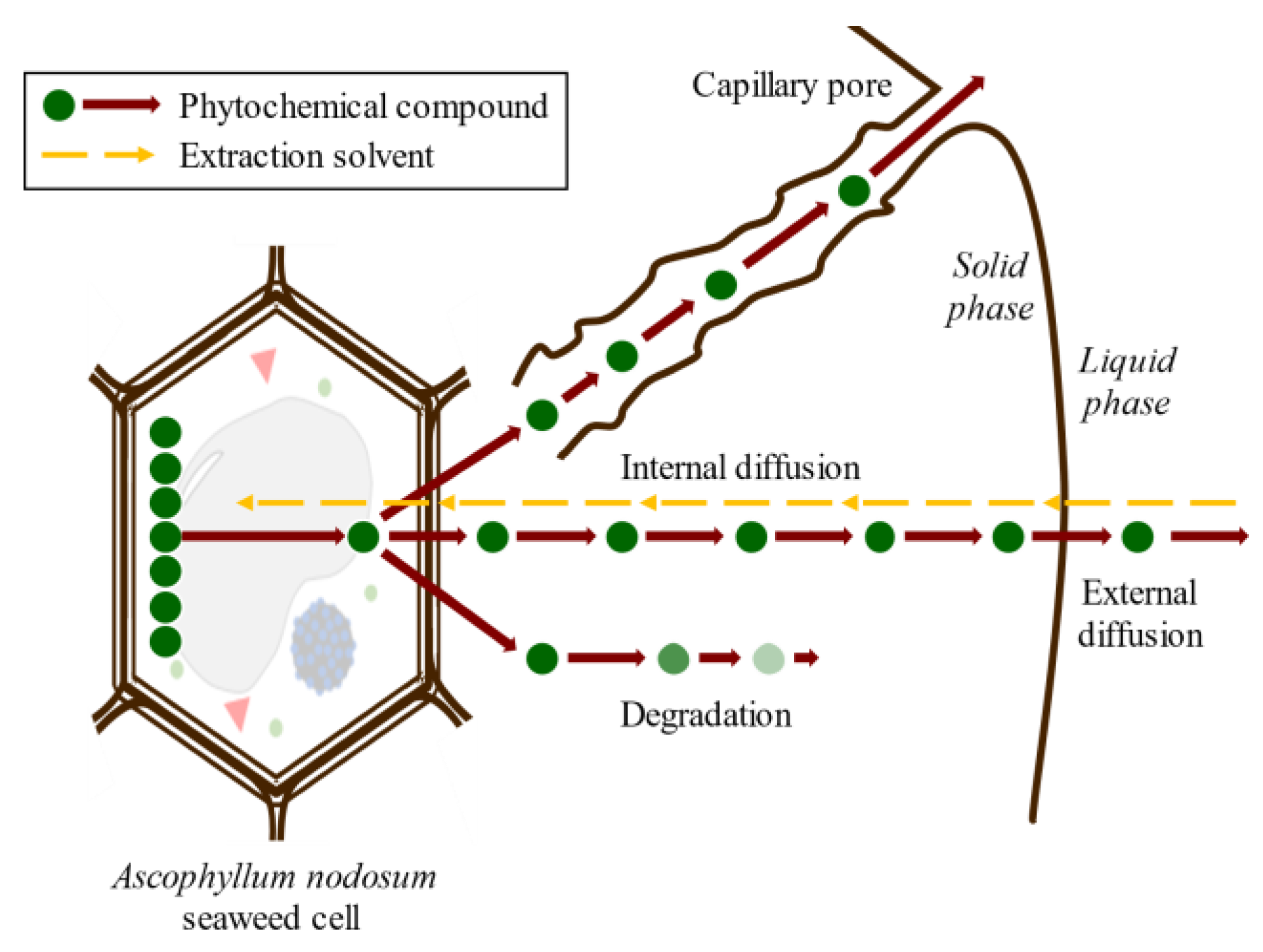
3. Phlorotannins’ Extraction and Isolation
3.1. Solid–Liquid Extraction
3.2. Ultrasound-Assisted Extraction
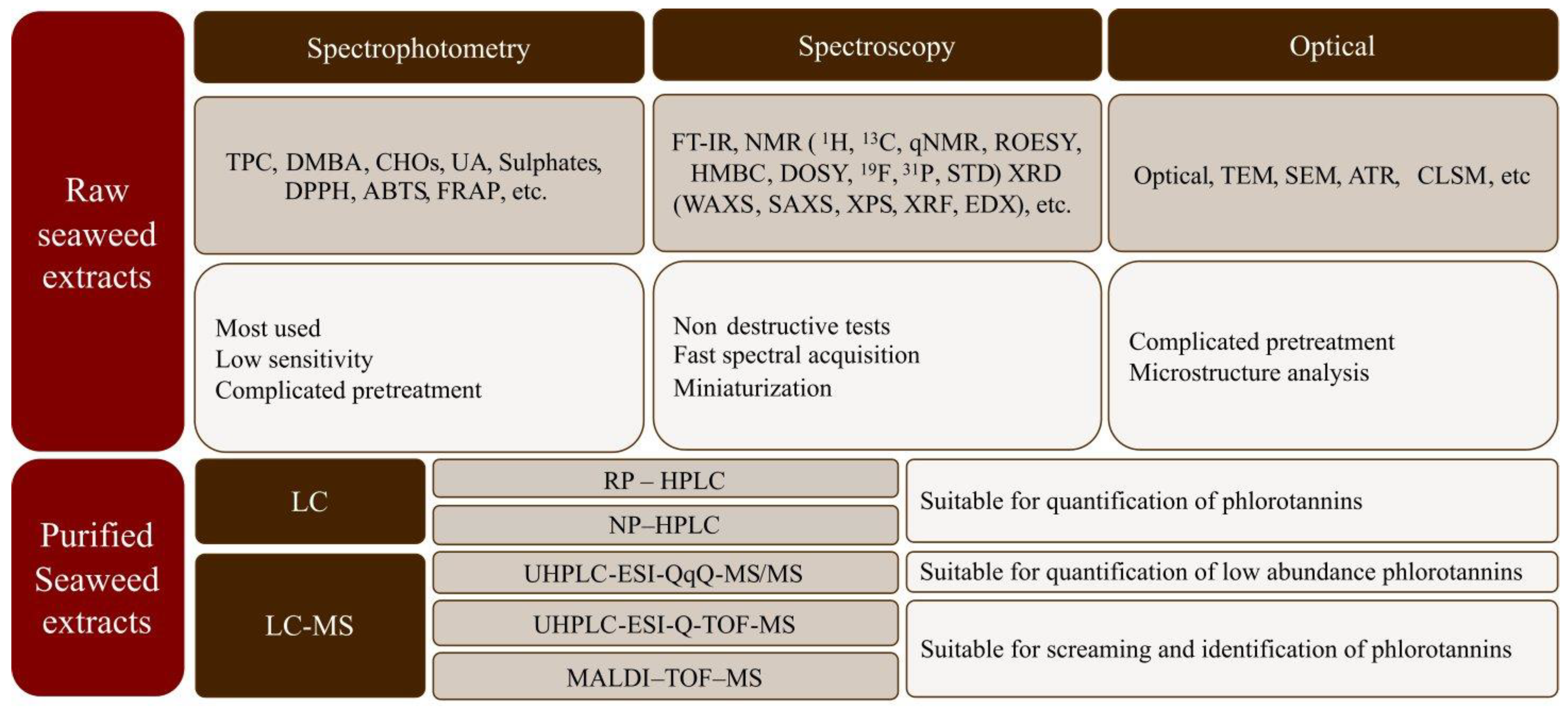
4. Characterization of Seaweed Phlorotannins
4.1. Antioxidant Activity Determination: Spectrophotometry Methods
4.1.1. Determination of Total Polyphenols Content (TPC)
4.1.2. Antioxidant Methods Based on the Transfer of Hydrogen Atoms
4.1.3. Antioxidant Methods Based on Electron Transfer
4.1.4. Antioxidant Methods Based on Proton and Electron Transfer
4.2. Chromatographic Methods
4.3. Fourier Transform Infrared Spectroscopy (FT-IR)
4.4. Nuclear Magnetic Resonance (NMR)
4.5. X-ray Diffraction (XRD)
4.6. Microscopy Techniques
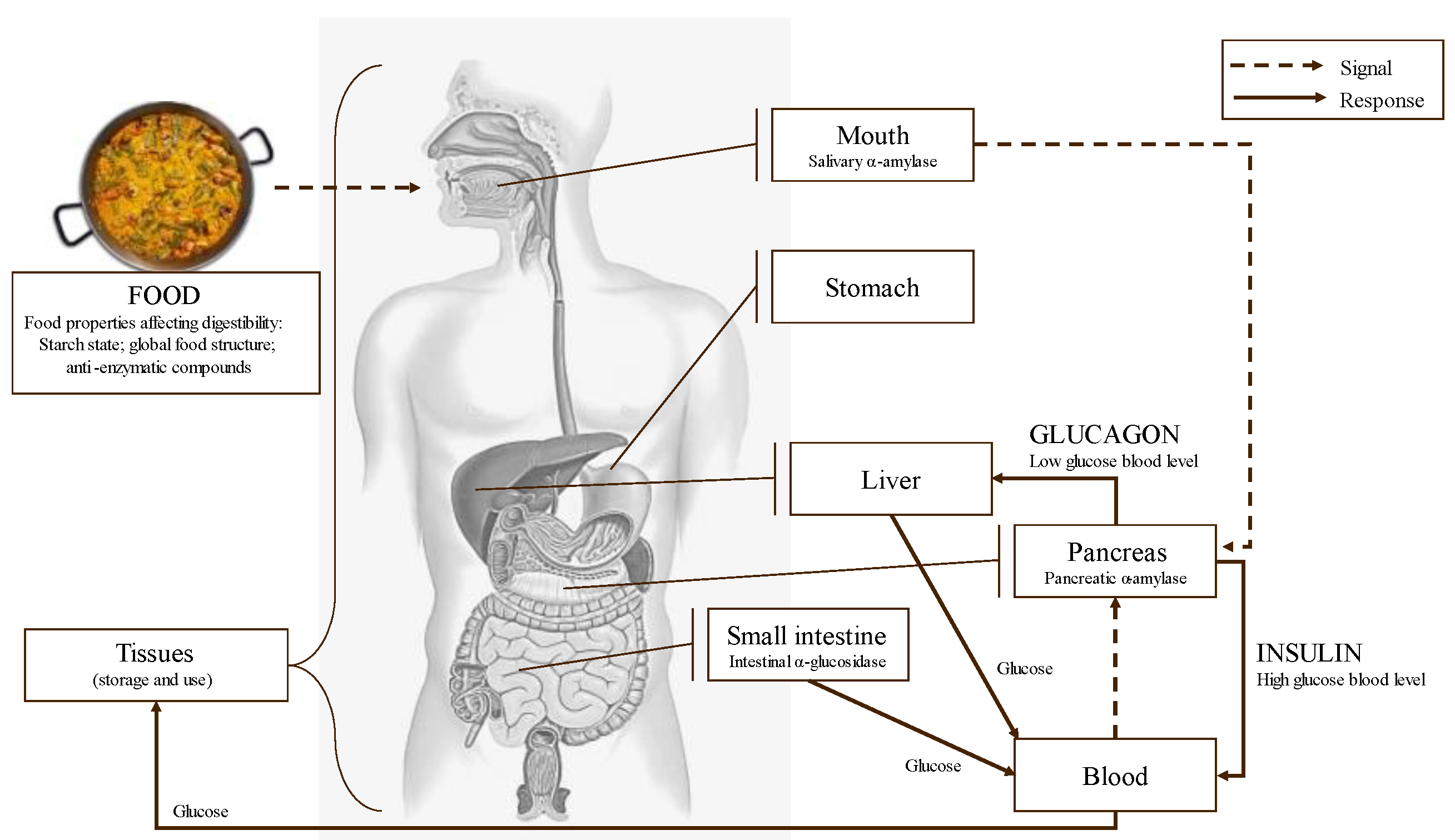
5. Potential Effects of Phlorotannins as Starch Digestive Enzymes Inhibitors in the Control of Glycemic Index
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valderrama, D. Social and economic dimensions of seaweed farming: A global review. In Proceedings of the Sixteenth Biennial Conference of the International Institute of Fisheries Economics and Trade, Dar es Salaam, Tanzania, 16–20 July 2012. [Google Scholar]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Graham, L.E.; Graham, J.; Wilcox, L.W. Introduction to algae. In Algae, 2nd ed.; Pearson: San Francisco, CA, USA, 2009; pp. 1–5. [Google Scholar]
- Lawton, R.J. The industrial ecology of freshwater macroalga for biomass applications. Algal Res. 2017, 24, 486–491. [Google Scholar] [CrossRef]
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M. Seaweed’s bioactive candidate compounds to food industry and global food security. Life 2020, 10, 140. [Google Scholar] [CrossRef]
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The Global Status of Seaweed Production, Trade, and Utilization; Food and Agriculture Organization of the United Nations: Roma, Italy, 2018; pp. 1–125. [Google Scholar]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Biological activities and potential cosmeceutical applications of bioactive components from brown seaweeds: A review. Phytochem. Rev. 2011, 10, 431–443. [Google Scholar] [CrossRef]
- Kilinç, B.; Cirik, S.; Turan, G.; Tekogul, H.; Koru, E. Seaweeds for Food and Industrial Applications. In Food Industry; Muzzalupo, I., Ed.; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Milledge, J.J.; Smith, B.; Dyer, P.W.; Harvey, P. Macroalgae-derived biofuel: A review of methods of energy extraction from seaweed biomass. Energies 2014, 7, 7194–7222. [Google Scholar] [CrossRef] [Green Version]
- Aderhold, D.; Williams, C.J.; Edyvean, R.G.J. The removal of heavy-metal ions by seaweeds and their derivatives. Bioresour. Technol. 1996, 58, 1–6. [Google Scholar] [CrossRef]
- McHugh, D.J. A Guide to the Seaweed Industry; FAO Fisheries Technical Paper Number; Food and Agriculture Organization of the United Nations: Roma, Italy, 2003; pp. 1–118. [Google Scholar]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Ford, L.; Theodoridou, K.; Sheldrake, G.N.; Walsh, P.J. A critical review of analytical methods used for the chemical characterisation and quantification of phlorotannin compounds in brown seaweeds. Phytochem. Anal. 2019, 30, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, C.; Shen, L.; Shin, H.C.; Lee, K.B.; Jif, B. Comparative analysis of oxidative mechanisms of phloroglucinol and dieckol by electrochemical, spectroscopic, cellular and computational methods. RSC Adv. 2018, 8, 1963–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajauria, G.; Jaiswal, A.K.; Abu-Ghannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown seaweed Himanthalia elongata from western coast of Ireland. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Kim, S.K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Wijesekara, I.; Yoon, N.Y.; Kim, S.K. Phlorotannins from Ecklonia cava (Phaeophyceae): Biological activities and potential health benefits. Biofactors 2010, 36, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic Potential of Marine Brown Algae—A Mini Review. J. Diabetes Res. 2020, 2020, 1230218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyon, M.; Labrecque, J. Functional foods: A conceptual definition. Br. Food J. 2008, 110, 1133–1149. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef] [PubMed]
- de Boer, A.; Urlings, M.J.E.; Bast, A. Active ingredients leading in health claims on functional foods. J. Funct. Foods 2016, 20, 587–593. [Google Scholar] [CrossRef]
- Apostolidis, E.; Lee, C.M. In vitro potential of Ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J. Food Sci. 2010, 75, 97–102. [Google Scholar] [CrossRef]
- Kim, K.T.; Rioux, L.E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef]
- Tierney, M.S.; Soler-Vila, A.; Rai, D.K.; Croft, A.K.; Brunton, N.P.; Smyth, T.J. UPLC-MS profiling of low molecular weight phlorotannin polymers in Ascophyllum nodosum, Pelvetia canaliculata and Fucus spiralis. Metabolomics 2014, 10, 524–535. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwai, K. Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK-Ay mice. Plant Foods Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef]
- Houchi, S.; Mahdadi, R.; Khenchouche, A.; Song, J.; Zhang, W.; Pang, X.; Zhang, L.; Sandalli, C.; Du, G. Investigation of common chemical components and inhibitory effect on GES-type β-lactamase (GES22) in methanolic extracts of Algerian seaweeds. Microb. Pathogenesis 2019, 126, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Alves, C.; Freitas, R.; Martins, A.; Pinteus, S.; Ribeiro, J.; Gaspar, H.; Alfonso, A.; Pedrosa, R. Antioxidant and neuroprotective potential of the brown seaweed Bifurcaria bifurcata in an in vitro Parkinson’s disease model. Mar. Drugs 2019, 17, 85. [Google Scholar] [CrossRef] [Green Version]
- Ragan, M.A.; Glombitzka, K.W. Phlorotannins, brown algal polyphenols. Prog. Phycol. Res. 1986, 4, 129–241. [Google Scholar]
- Kumar, L.R.G.; Paul, P.T.; Anas, K.K.; Tejpal, C.S.; Chatterjee1, N.; Anupama, T.K.; Mathew, S.; Ravishankar, C.N. Phlorotannins-bioactivity and extraction perspectives. J. Appl. Phycol. 2022, 34, 2173–2185. [Google Scholar] [CrossRef]
- Leyton, A.; Pezoa-Conte, P.; Barriga, A.; Buschmann, A.H.; Maki-Arvela, P.; Mikkola, J.P.; Lienqueo, M.E. Identification and efficient extraction method of phlorotannins from the brown seaweed Macrocystis pyrifera using an orthogonal experimental design. Algal Res. 2016, 16, 201–208. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Soria, A.C.; Villamiel, M. Effect of ultrasound on the technological properties and bioactivity of food: A review. Trends Food Sci. Technol. 2010, 21, 323–331. [Google Scholar] [CrossRef]
- Shahidi, F.; Naczk, M. Phenolics in Food and Nutraceuticals, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of phlorotannins extraction from Fucus vesiculosus and evaluation of their potential to prevent metabolic disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [Green Version]
- Singh, I.P.; Sidana, J. Phlorotannins. In Woodhead Publishing Series in Food Science, Technology and Nutrition, Functional Ingredients from Algae for Foods and Nutraceuticals; Dominguez, H., Ed.; Woodhead Publishing: Cambridge, MA, USA, 2013; pp. 181–204. [Google Scholar]
- Lopes, G.; Andrade, P.B.; Valentão, P. Phlorotannins: Towards new pharmacological interventions for diabetes mellitus type 2. Molecules 2017, 22, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermund, D.B.; Plaza, M.; Turner, C.; Jonsdottir, R.; Kristinsson, H.G.; Jacobsen, C.; Nielsen, K.F. Structure dependent antioxidant capacity of phlorotannins from Icelandic Fucus vesiculosus by UHPLCDAD-ECD-QTOFMS. Food Chem. 2018, 240, 904–909. [Google Scholar] [CrossRef] [Green Version]
- Machu, L.; Misurcova, L.; Ambrozova, J.V.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisbert, M.; Sineiro, J.; Moreira, R. Polyphenols extraction kinetics from Ascophyllum nodosum seaweed employing water and saltwater: Effect of ultrasound sonication. Algal Res. 2022, 66, 102773. [Google Scholar] [CrossRef]
- Tiwari, B.K. Ultrasound: A clean, green extraction technology. TrAC Trends Anal. Chem. 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurized liquid extraction and solid-liquid extraction methods on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Sci. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Sarkar, S.; Gayen, K.; Bhowmick, T.K. Green extraction of biomolecules from algae using subcritical and supercritical fluids. Biomass Conv. Bioref. 2022, 1–23. [Google Scholar] [CrossRef]
- Vissers, A.M.; Caligiani, A.; Sforza, S.; Vincken, J.P.; Gruppen, H. Phlorotannin composition of Laminaria digitata. Phytochem. Anal. 2017, 28, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Luo, G.; Wang, L.; Yuan, W. Optimization of antioxidant extraction from edible brown algae Ascophyllum nodosum using response surface methodology. Food Bioprod. Process. 2019, 114, 205–215. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, W.; Zhao, R. Extraction of antioxidants from brown algae Ascophyllum nodosum using a binary solvent extraction system. ACS Food Sci. Technol. 2021, 1, 1041–1049. [Google Scholar] [CrossRef]
- Sánchez-Camargo, A.D.; Montero, L.; Stiger-Pouvreau, T.; Tanniou, A.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Considerations on the use of enzyme-assisted extraction in combination with pressurized liquids to recover bioactive compounds from algae. Food Chem. 2016, 192, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, A.; Goosen, N. Green and integrated processing approaches for the recovery of high-value compounds from brown seaweeds. Adv. Bot. Res. 2019, 95, 369–413. [Google Scholar] [CrossRef]
- Flores-Fernandez, N.; Gonzalez-Munoz, M.J. Ultrasound-assisted extraction of bioactive carbohydrates. In Water Extraction of Bioactive Compounds; Dominguez-Gonzalez, H., Gonzalez-Munoz, M.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 317–331. [Google Scholar] [CrossRef]
- Picó, Y. Ultrasound-assisted extraction for food and environmental samples. TrAC Trends Anal. Chem. 2013, 43, 84–99. [Google Scholar] [CrossRef]
- Roselló-Soto, E.; Galanakis, C.M.; Brncic, M.; Orlien, V.; Trujillo, F.J.; Mawson, R.; Knoerzer, K.; Tiwari, B.K.; Barba, F.J. Clean recovery of antioxidant compounds from plant foods, by-products and algae assisted by ultrasounds processing. Modelling approaches to optimize processing conditions. Trends Food Sci. Technol. 2015, 42, 134–149. [Google Scholar] [CrossRef]
- Ummat, V.; Tiwari, B.K.; Jaiswal, A.K.; Condon, K.; Garcia-Vaquero, M.; O’Doherty, J.; O’Donnell, C.; Rajauria, G. Optimization of ultrasound frequency, extraction time and solvent for the recovery of polyphenols, phlorotannins and associated antioxidant activity of brown seaweeds. Mar. Drugs 2020, 18, 250. [Google Scholar] [CrossRef]
- Sardari, R.R.; Prothmann, J.; Gregersen, O.; Turner, C.; Karlsson, E.N. Identification of phlorotannins in the brown algae, Saccharina latissima and Ascophyllum nodosum by ultra-high-performance liquid chromatography coupled to high-resolution tandem mass spectrometry. Molecules 2021, 26, 43. [Google Scholar] [CrossRef]
- Poole, J.; Diop, A.; Rainville, L.C.; Barnabe, S. Bioextracting polyphenols from the brown seaweed Ascophyllum nodosum from Québec’s north shore coastline. Ind. Biotechnol. 2019, 15, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Barba, F.J.; Lorenzo, J.M. Antioxidant potential of extracts obtained from macro- (Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata) and micro-algae (Chlorella vulgaris and Spirulina platensis) assisted by ultrasound. Medicines 2018, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Cikoš, A.M.; Jokic, S.; Šubaric, D.; Jerkovic, I. Overview on the application of modern methods for the extraction of bioactive compounds from marine macroalgae. Mar. Drugs 2018, 16, 348. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Moreira, R.; Sineiro, J.; Chenlo, F.; Arufe, S.; Díaz-Varela, D. Aqueous extracts of Ascophyllum nodosum obtained by ultrasound-assisted extraction: Effects of drying temperature of seaweed on the properties of extracts. J. Appl. Phychol. 2017, 29, 3191–3200. [Google Scholar] [CrossRef]
- Kadam, S.U.; O’Donnell, C.P.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish brown seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound-assisted extraction, characterisation, and bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, S.; O’Donnell, C.P. Effect of ultrasound pretreatment on the extraction kinetics of bioactives from brown seaweed (Ascophyllum nodosum). Sep. Sci. Technol. 2015, 50, 670–675. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; Smyth, T.J.; O’Donnell, C.P. Optimization of ultrasound assisted extraction of bioactive components from brown seaweed Ascophyllum nodosum using surface methodology. Ultrason. Sonochem. 2015, 23, 308–316. [Google Scholar] [CrossRef]
- Pantidos, N.; Boath, A.; Lund, V.; Conner, S.; McDougall, G.J. Phenolic-rich extracts from the edible seaweed, Ascophyllum nodosum, inhibit α-amylase and α-glucosidase: Potential anti-hyperglycemic effects. J. Funct. Foods 2014, 10, 201–209. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanni, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Zhang, J.; Tiller, C.; Shen, J.; Wang, C.; Girouard, G.S.; Dennis, D.; Barrow, C.J.; Miao, M.; Stewart, H.S. Antidiabetic properties of polysaccharide- polyphenolic- enriched fractions from the brown seaweed Ascophyllum nodosum. Can. J. Physiol. Pharmacol. 2007, 85, 1116–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Jónsdóttir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Okada, Y.; Ishimaru, A.; Suzuki, R.; Okuyama, T. A new phloroglucinol derivative from the brown alga Eisenia bicyclis: Potential for the effective treatment of diabetic complications. J. Nat. Prod. 2004, 67, 103–105. [Google Scholar] [CrossRef]
- Jung, H.A.; Yoon, N.Y.; Woo, M.H.; Choi, J.S. Inhibitory activities of extract from several kinds of seaweeds and phlorotannins from brown alga Ecklonia stolonifera on glucose-mediated protein damage and rat lens aldose reductase. Fish Sci. 2008, 74, 1363–1365. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Lee, S.H.; Li, Y.; Kim, S.K. Phlorotannins from Ishige okamurae and acetyl- and butyrylcholinesterase inhibitory effects. J. Funct. Foods 2009, 1, 331–335. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Shibata, T.; Ishimaru, K.; Kawaguchi, S.; Yoshikawa, H.; Hama, Y. Antioxidant activities of phlorotannins isolated from Japanese Laminariaceae. J. Appl. Phycol. 2008, 20, 705–711. [Google Scholar] [CrossRef]
- Nakamura, T.; Nagayama, K.; Uchida, K.; Tanaka, R. Antioxidant activity of phlorotannins isolated from the brown alga Esinia bicyclis. Fish Sci. 1996, 62, 923–926. [Google Scholar] [CrossRef] [Green Version]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Han, J.S.; Kim, H.J.; Jeon, Y.J. Diphlorethohydroxycarmalol isolated from Ishige okamurae, a brown alga, a potent α-glucosidase and α-amylase inhibitor, alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2009, 615, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinzyo, Z.; Kido, M.; Mori, H.; Nakayama, Y.; Takahashi, M. Structure of an antiplasmin inhibitor, eckol, isolated from the brown algae Ecklonia kurome OKAMURA and inhibitory activities of its derivatives on plasma-plasmin inhibitor. Chem. Pharm. Bull. 1989, 37, 349–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, Y.M.; Baik, J.S.; Hyun, J.W.; Lee, N.H. Isolation of a new phlorotannins, fucodiphlorethol G, from a brown alga Ecklonia cava. BKCS 2007, 28, 1595–1597. [Google Scholar] [CrossRef] [Green Version]
- Sailler, B.; Glombitza, K.W. Phlorethols and fucophlorethols from the brown alga Cystophora retroflexa. Phytochemistry 1999, 50, 869–881. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Keusgen, M.; Hauperich, S. Fucophlorethols from the brown algae Sargassum spinuligerum and Cystophora torulosa. Phytochemistry 1997, 46, 1417–1422. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinzyo, Z.; Mori, H.; Nakayama, Y.; Takahashi, M. Anti-plasmin inhibitor. VI.: Structure of phlorofucofuroeckol A, a novel phlorotannin with both dibenzo-1, 4-dioxin and dibenzofuran elements, from Ecklonia kurome OKAMURA. Chem. Pharm. Bull. 1990, 38, 133–135. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, Y.; Matsuda, K.; Yamada, Y.; Nishikawa, M.; Shioya, K.; Katsumaki, H.; Imai, K.; Amano, H. Isolation of a new antiallergic phlorotannins, phlorofucofuroeckol-B, from an edible brown alga, Eiseenia arborea. Biosci. Biotechnol. Biochem. 2006, 70, 2807–2811. [Google Scholar] [CrossRef] [Green Version]
- Glombitza, K.W.; Rösener, H.U. Bifuhalol: Ein diphenyläther aus Bifurcaria bifurcata. Phytochemistry 1974, 13, 1245–1247. [Google Scholar] [CrossRef]
- Kang, K.A.; Lee, K.H.; Chae, S.W.; Koh, Y.S.; Yoo, B.S.; Kim, J.H.; Ham, Y.M.; Baik, J.S.; Lee, N.H.; Hyun, J.W. Triphlorethol-A from Ecklonia cava protects V79-4 lung fibroblast against hydrogen peroxide induced cell damage. Free Radic. Res. 2005, 39, 883–892. [Google Scholar] [CrossRef]
- Tanniou, A.; Vandanjon, L.; Incera, M.; Leon, E.S.; Husa, V.; Le Grand, J.; Nicolas, J.L.; Poupart, N.; Kervarec, N.; Engelen, A.; et al. Assessment of the spatial variability of phenolic contents and associated bioactivities in the invasive alga Sargassum muticum sampled along its European range from Norway to Portugal. J. Appl. Phycol. 2014, 26, 1215–1230. [Google Scholar] [CrossRef] [Green Version]
- Pulido, R.; Bravo, L.; Calixto, F.D.S. Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/ antioxidant power assay. J. Agric. Food Chem. 2000, 48, 3396–3402. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.A.O.; Félix, R.; Pais, A.C.S.; Rocha, S.M.; Silvestre, A.J.D. The quest for phenolic compounds from macroalgae: A Review of extraction and identification methodologies. Biomolecules 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the Folin-Ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [Green Version]
- Apak, R.; Çapanoglu, E.; Shahidi, F. Measurement of Antioxidant Activity and Capacity: Recent Trends and Applications; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 278–283. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. AJEV 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Becker, E.M.; Nissen, L.R.; Skibsted, L.H. Antioxidant evaluation protocols: Food quality or health effects. Eur. Food Res. Technol. 2004, 219, 561–571. [Google Scholar] [CrossRef]
- Ciz, M.; Cížová, H.; Denev, P.; Kratchanova, M.; Slavov, A.; Lojek, A. Different methods for control and comparison of the antioxidant properties of vegetables. Food Control 2010, 21, 518–523. [Google Scholar] [CrossRef]
- Regoli, F.; Winston, G.W. Quantification of total oxidant scavenging capacity of antioxidants for peroxynitrite, peroxyl radicals, and hydroxyl radicals. Toxicol. Appl. Pharmacol. 1999, 156, 96–105. [Google Scholar] [CrossRef]
- Antolovich, M.; Prenzler, P.D.; Patsalides, E.; McDonald, S.; Robards, K. Methods for testing antioxidant activity. Analyst 2002, 127, 183–198. [Google Scholar] [CrossRef]
- Williams, D.E. Structure of 2,2-diphenyl-1-picrylhydrazyl free radical. J. Am. Chem. Soc. 1966, 88, 5665–5666. [Google Scholar] [CrossRef]
- Staško, A.; Brezová, V.; Biskupic, S.; Mišík, V. The potential pitfalls of using 1,1-diphenyl-2-picrylhydrazyl to characterize antioxidants in mixed water solvents. Free Radic. Res. 2007, 41, 379–390. [Google Scholar] [CrossRef]
- Miller, N.J.; Diplock, A.T.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Lemanska, K.; Szymusiak, H.; Tyrakowska, B.; Zielinski, R.; Soffers, A.E.; Rietjens, I.M. The influence of pH on antioxidant properties and the mechanism of antioxidant action of hydroxyflavones. Free Radic. Biol. Med. 2001, 31, 869–881. [Google Scholar] [CrossRef]
- Walker, R.B.; Everette, J.D. Comparative reaction rates of various antioxidants with ABTS radical cation. J. Agric. Food Chem. 2009, 57, 1156–1161. [Google Scholar] [CrossRef]
- Yoon, J.S.; Yadunandam, A.K.; Kim, S.J.; Woo, H.C.; Kim, H.R.; Kim, G.D. Dieckol, isolated from Ecklonia stolonifera, induces apoptosis in human hepatocellular carcinoma Hep3B cells. J. Nat. Med. 2013, 67, 519–527. [Google Scholar] [CrossRef]
- Novakova, L.; Matysova, L.; Solich, P. Advantages of application of UPLC in pharmaceutical analysis. Talanta 2006, 68, 908–918. [Google Scholar] [CrossRef]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Motilva, M.J.; Serra, A.; Macia, A. Analysis of food polyphenols by ultra high-performance liquid chromatography coupled to mass spectrometry: An overview. J. Chromatogr. A 2013, 1292, 66–82. [Google Scholar] [CrossRef]
- Allwood, J.W.; Evans, H.; Austin, C.; McDougall, G.J. Extraction, enrichment, and LC-MSn-based characterization of phlorotannins and related phenolics from the brown seaweed, Ascophyllum nodosum. Mar. Drugs 2020, 18, 448. [Google Scholar] [CrossRef] [PubMed]
- Konermann, L.; Ahadi, E.; Rodriguez, A.D.; Vahidi, S. Unravelling the mechanism of electrospray ionization. Anal. Chem. 2013, 85, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.; Cai, Z. The latest developments and applications of mass spectrometry in food-safety and quality analysis. TrAC Trends Anal. Chem. 2013, 52, 170–185. [Google Scholar] [CrossRef]
- Dong, D.; Jiao, L.; Li, C.; Zhao, C. Rapid and real-time analysis of volatile compounds released from food using infrared and laser spectroscopy. TrAC Trends Anal. Chem. 2019, 110, 410–416. [Google Scholar] [CrossRef]
- Lim, S.N.; Cheung, P.C.K.; Ooi, V.E.C.; Ang, P.O. Evaluation of antioxidative activity of extracts from a brown seaweed, Sargassum siliquastrum. J. Agric. Food Chem. 2002, 50, 3862–3866. [Google Scholar] [CrossRef]
- Parys, S.; Kehraus, S.; Pete, R.; Küpper, F.C.; Glombitza, K.W.; König, G.M. Seasonal variation of polyphenolics in Ascophyllum nodosum (Phaeophyceae). Eur. J. Phycol. 2009, 44, 331–338. [Google Scholar] [CrossRef]
- Hesse, M.; Meier, H.; Zeeh, B. Spektroskopiche Methoden in der Organischen Chemie, 7th ed.; Georg Thieme: Stuttgart, Germany, 2005; pp. 44–52. [Google Scholar]
- Igoumenidis, P.E.; Zoumpoulakis, P.; Karathanos, V.T. Physicochemical interactions between rice starch and caffeic acid during boiling. Int. Food Res. J. 2018, 109, 589–595. [Google Scholar] [CrossRef]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Ar Gall, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical-scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Shimanuki, N.; Imai, M.; Nagai, K. Effects of counter cations on the water vapor sorption properties of alginic acid and alginates. J. Appl. Polym. Sci. 2020, 137, e49326. [Google Scholar] [CrossRef] [Green Version]
- Milinovic, J.; Figueiredo, F.M.L.; Noronha, J.P.; Sardinha, J. Application of ED-XRF spectra for determination of macroelements in edible seaweeds. J. Food Compost. Anal. 2022, 110, 104559. [Google Scholar] [CrossRef]
- André, R.; Guedes, L.; Melo, R.; Ascensão, L.; Pacheco, R.; Vaz, P.D.; Serralheiro, M.L. Effect of food preparations on in vitro bioactivities and chemical components of Fucus vesiculosus. Foods 2020, 9, 955. [Google Scholar] [CrossRef]
- Hutzler, P.; Fischbach, R.; Heller, W.; Jungblut, T.P.; Reuber, S.; Schmitz, R.; Veit, M.; Weissenböck, G.; Schnitzler, J.P. Tissue localization of phenolic compounds in plants by confocal laser scanning microscopy. J. Exp. Bot. 1998, 49, 953–965. [Google Scholar] [CrossRef]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant starch—A review. CRFSFS 2006, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch structure and digestibility enzyme-substrate relationship. World’s Poult. Sci. J. 2004, 60, 186–195. [Google Scholar] [CrossRef]
- Parada, J.; Aguilera, J.M. Review: Starch matrices and the glycemic response. FSTI 2011, 17, 187–204. [Google Scholar] [CrossRef]
- Santos, J.L.; Saus, E.; Smalley, S.V.; Cataldo, L.R.; Alberti, G.; Parada, J.; Gratacòs, M.; Estivill, X. Copy-number polymorphism of the salivary amylase gene: Implications in human nutrition research. J. Nutr. Nutr. 2012, 5, 117–131. [Google Scholar] [CrossRef]
- Parada, J.; Santos, J.L. Interactions among starch, lipids, and proteins in foods: Microstructure control for glycemic response modulation. Crit. Rev. Food Sci. Nutr. 2016, 56, 2362–2369. [Google Scholar] [CrossRef]
- Cummings, J.H.; Englyst, H.N. Gastrointestinal effects of food carbohydrate. AJCN 1995, 61, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Díez-Sánchez, E.; Quiles, A.; Hernando, I. Interactions between blackcurrant polyphenols and food macronutrients in model systems: In vitro digestion studies. Foods 2021, 10, 847. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, S.; Bai, F.; Cao, J.; Sun, L. The physical adsorption of gelatinized starch with tannic acid decreases the inhibitory activity of the polyphenol against α-amylase. Foods 2021, 10, 1233. [Google Scholar] [CrossRef] [PubMed]
- Deladino, L.; Teixeira, A.S.; Navarro, A.S.; Álvarez, I.; Molina-García, A.D.; Martino, M. Corn starch systems as carriers for yerba mate (Ilex paraguariensis) antioxidants. Food Bioprod. Process. 2015, 94, 463–472. [Google Scholar] [CrossRef]
- Hall, A.C.; Fairclough, A.C.; Mahadevan, K.; Paxman, J.R. Ascophyllum nodosum enriched bread reduces subsequent energy intake with no effect on post-prandial glucose and cholesterol in healthy, overweight males. A pilot study. Appetite 2012, 58, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamat, H.; Matanjun, P.; Ibrahim, S.; Amin, S.F.; Abdul-Hamid, M.; Rameli, A.S. The effect of seaweed composite our on the textural properties of dough and bread. J. Appl. Phycol. 2014, 26, 1057–1062. [Google Scholar] [CrossRef]
- Menezes, B.S.; Coelho, M.S.; Meza, S.L.R.; Salas-Mellado, M.; Souza, M.R.A.Z. Macroalgal biomass as an additional ingredient of bread. Int. Food Res. J. 2015, 22, 812–817. [Google Scholar]
- Allsopp, P.; Crowe, W.; Bahar, B.; Harnedy, P.A.; Brown, E.S.; Taylor, S.S.; Smyth, T.J.; Soler-Vila, A.; Magee, P.J.; Gill, C.; et al. The effect of consuming Palmaria palmata enriched bread on inflammatory markers, antioxidant status, lipid profile and thyroid function in a randomised placebo-controlled intervention trial in healthy adults. Eur. J. Nutr. 2016, 55, 1951–1962. [Google Scholar] [CrossRef]
- Rózylo, R.; Hassoon, W.H.; Gawlik-Dziki, U.; Siasta, M.; Dziki, D. Study on the physical and antioxidant properties of gluten-free bread with brown algae. CyTA-J. Food Sci. 2017, 15, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Parada, J.; Pérez-Correa, J.; Pérez-Jiménez, J. Design of low glycemic response foods using polyphenols from seaweed. J. Funct. Foods 2019, 56, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Tanaka, T.; Tamura, S.; Toshima, S.; Tamaya, K.; Miyata, Y.; Tanaka, K.; Matsumoto, K. α-Glucosidase inhibitory profile of catechins and theaflavins. J. Agric. Food Chem. 2007, 55, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural products for the treatment of type 2 diabetes mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zheng, Y.; Shen, Y. Voglibose (Basen®, AO-128), one of the most important α-glucosidase inhibitors. Curr. Med. Chem. 2006, 13, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. In vitro studies of eggplant (Solanum melongena) phenolics as inhibitors of key enzymes relevant for type 2 diabetes and hypertension. Bioresour. Technol. 2008, 99, 2981–2988. [Google Scholar] [CrossRef]
- Williamson, G. Possible effects of dietary polyphenols on sugar absorption and digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef]
- Flores, F.P.; Singh, R.K.; Kerr, W.L.; Pegg, R.B.; Kong, F. Antioxidant and enzyme inhibitory activities of blueberry anthocyanins prepared using different solvents. J. Agric. Food Chem. 2013, 61, 4441–4447. [Google Scholar] [CrossRef]
- Gauer, J.S.; Tumova, S.; Lippiat, J.D.; Kerimi, A.; Williamson, G. Differential patterns of inhibition of the sugar transporters GLUT2, GLUT5 and GLUT7 by flavonoids. Biochem. Pharmacol. 2018, 152, 11–20. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Renard, C.M.G.C. Interactions between polyphenols and macromolecules: Quantification methods and mechanisms. Crit. Rev. Food Sci. Nutr. 2012, 52, 213. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- Lee, S.; Jeon, Y. Anti-diabetic effects of brown algae derived phlorotannins, marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef]
- Liu, B.; Kongstad, K.T.; Wiese, S.; Jäger, A.K.; Staerk, D. Edible seaweed as future functional food: Identification of α-glucosidase inhibitors by combined use of high-resolution a α-glucosidase inhibition profiling and HPLC-HRMS-SPE-NMR. Food Chem. 2016, 203, 16–22. [Google Scholar] [CrossRef]
- Roy, M.C.; Anguenot, R.; Fillion, C.; Beaulieu, M.; Bérubé, J.; Richard, D. Effect of a commercially-available algal phlorotannins extract on digestive enzymes and carbohydrate absorption In Vivo. Food Res. Int. 2011, 44, 3026–3029. [Google Scholar] [CrossRef]
- Kellogg, J.; Grace, M.H.; Lila, M.A. Phlorotannins from Alaskan seaweed inhibit carbolytic enzyme activity. Mar. Drugs 2014, 12, 5277–5294. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Li, Y.; Karadeniz, F.; Kim, M.M.; Kima, S.K. α-Glucosidase and α-amylase inhibitory activities of phloroglucinol derivatives from edible marine brown alga, Ecklonia cava. J. Sci. Food Agric. 2009, 89, 1552–1558. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, M.H.; Heo, S.J.; Kang, S.M.; Ko, S.C.; Han, J.S.; Jeon, Y.J. Dieckol isolated from Ecklonia cava inhibits α-glucosidase and α-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef] [PubMed]
- You, H.N.; Lee, H.A.; Park, M.H.; Lee, J.H.; Han, J.S. Phlorofucofuroeckol A isolated from Ecklonia cava alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2015, 752, 92–96. [Google Scholar] [CrossRef]
- Moon, H.E.; Islam, M.N.; Ahn, B.R.; Chowdhury, S.S.; Sohn, H.S.; Jung, H.A.; Cho, J.S. Protein tyrosine phosphatase 1B and α-glucosidase inhibitory phlorotannins from edible brown algae, Ecklonia stolonifera and Eisenia bicyclis. Biosci. Biotechnol. Biochem. 2011, 75, 1472–1480. [Google Scholar] [CrossRef] [Green Version]
- Eom, S.H.; Lee, S.H.; Yoon, N.Y.; Jung, W.K.; Jeon, Y.J.; Kim, S.K.; Leeh, M.S.; Kima, Y.M. α-Glucosidase- and α-amylase-inhibitory activities of phlorotannins from Eisenia bicyclis. J. Sci. Food Agric. 2012, 92, 2084–2090. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, S.M.; Ko, S.C.; Moon, S.H.; Jeon, B.T.; Lee, D.H.; Jeon, Y.J. Octaphlorethol A: A potent a-glucosidase inhibitor isolated from Ishige foliacea shows an antihyperglycemic effect in mice with streptozotocin-induced diabetes. Food Funct. 2014, 5, 2602–2609. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Hang, J.S. Hypoglycemic Effect of Sargassum ringgoldianum Extract in STZ-induced Diabetic Mice. Prev. Nutr. Food Sci. 2012, 17, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Firdaus, M.; Prihanto, A.A. α-Amylase and α-glucosidase inhibition by brown seaweed (Sargassum sp) extracts. J. Life Sci. 2014, 1, 6–11. [Google Scholar] [CrossRef]
- Kawamura-Konishi, Y.; Watanabe, N.; Saito, M.; Nakajima, N.; Sakaki, T.; Katayama, T.; Enomoto, T. Isolation of a new phlorotannin, a potent inhibitor of carbohydrate-hydrolyzing enzymes, from the brown alga Sargassum patens. J. Agric. Food Chem. 2012, 60, 5565–5570. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Extractant | LS (gsol/galgae) | Time | Temperature | TPC | Reference |
|---|---|---|---|---|---|---|
| SLE | Ethanol | 90 | 30 min | 30 °C | 0.7 mgPE/mgextract | [47] |
| SLE | Water | 100 | 1 h | 65 °C | 7.3 mgPE/gextract | [54] |
| Ethanol (30% v/v) | 5 | 30 min | 25 °C | 4.1 mgPE/gextract | ||
| Ethanol (80% v/v) | 10 | 20 (+5) h | 25 (+65) °C | 3.4 mgPE/gextract | ||
| SLE | Acetone (70% v/v) | 20 | 3 h | rt | 24.5 mgPE/gextract. | [13] |
| SLE | Ethanol (50% v/v) | 15 | 4 h | 20 °C | 0.2 gPE/gextract | [53] |
| UAE | 10 | 30 min | NS (35 kHz) | 0.4 gPE/gextract | ||
| UAE | Ethanol (40% v/v) | 50 | 30 min | 60 °C | 0.5 gPE/gextract | [46] |
| SLE | Water | 10 | 24 h | 4 °C | 52 mgPE/gextract | [55] |
| Methanol (50% v/v) | 77 mgPE/gextract | |||||
| Ethanol (75% v/v) | 95.4 mgPE/gextract | |||||
| Dioxolane (75% v/v) | 90 mgPE/gextract | |||||
| 1,3-propanediol | 98.5 mgPE/gextract | |||||
| UAE | Ethanol (50% v/v) | 10 | 30 min | rt | 46.6 mg/gextract | [56] |
| SLE | Methanol (70% v/v) | 10 | 4 h | rt | 0.5 gGAE/gextract | [57] |
| MAE (2.45 GHz) | 15 min | 110 °C | 1.4 gGAE/gextract | |||
| SLE | Methanol (70% v/v) | 10 | 4 h | rt | 0.5 mgGAE/gd.b | [58] |
| MAE (2.45 GHz) | 15 min | 110 °C | 1.4 mgGAE/gd.b | |||
| UAE (80 W/cm2) | Acetone (70% v/v) | 30 | 4 min | <35 °C | 31.8 mgPE/gd.b | [59] |
| SLE | Water | 20 | 150 min | 70 °C | 0.17 mgPE/gd.b | [60] |
| HCl (0.1 M) | 0.11 mgPE/gd.b | |||||
| UAE (35.6 W/cm2) | Water | 15 min | <35 °C | 0.16 mgPE/gd.b | ||
| HCl (0.1 M) | 0.13 mgPE/gd.b | |||||
| UAE (75.8 W/cm2) + SLE | HCl (0.03 M) | 10 | 10 min + 22 h | <35 °C | 135.7 mgGAE/gd.b | [61] |
| UAE (75.8 W/cm2) | HCl (0.03 M) | 10 | 25 min | <35 °C | 143.1 mgGAE/gd.b | [62] |
| SLE | Water | 10 | 60 min | 20 °C | 178.0 mgGAE/mL | [63] |
| HCl (5 mM) | 210.0 mgGAE/mL | |||||
| SLE | Water | 20 | 24 h | rt | 70.5 mgPE/gextract | [43] |
| Ethanol (80% v/v) | 10 | 66.3 mgPE/gextract | ||||
| Acetone (80% v/v) | 155.9 mgPE/gextract | |||||
| PLE | Water | 2 | NS | 120 °C (1500 psi) | 93.4 mgPE/gextract | |
| Ethanol (80% v/v) | 100 °C (1000 psi) | 101.3 mgPE/gextract | ||||
| Acetone (80% v/v) | 60 °C (1000 psi) | 127.4 mgPE/gextract | ||||
| SLE | Methanol (60% v/v) | 15 | 3 h | 40 °C | 4.5 mgGAE/gd.b | [64] |
| SLE | Ethanol (50% v/v) | 12.5 | 90 min | 80 °C | 38.8 mgPE/gd.b | [65] |
| SLE | Water | 20 | 24 h | rt | 138.0 mgPE/gextract | [66] |
| Acetone (70% v/v) | 159.0 mgPE/gextract |
| Phlorotannin | Seaweed | Reference | |
|---|---|---|---|
| 2-Phloroeckol |  | Eisenia bicyclis | [67] |
| Euphorbia stolonifera | [68] | ||
| 6,6′-Bieckol |  | Ishige okamurae | [69] |
| 7-Phloroeckol |  | Euphorbia stolonifera | [68] |
| Ascophyllum nodosum | [70] | ||
| 8,8′-Bieckol |  | Eisenia bicyclis, Ecklonia cava and Ecklonia kurome | [71] |
| Dieckol |  | Eisenia bicyclis | [72] |
| Dioxinodehydroeckol |  | Euphorbia stolonifera | [68] |
| Diphlorethohydroxycarmalol |  | Ishige okamurae | [73] |
| Eckol |  | Ecklonia kurome | [74] |
| Fucodiphloroethol G |  | Ecklonia cava | [75] |
| Fucophlorethols |  | Cystophora retroflexa | [76] |
| Cupressus torulosa | [77] | ||
| Sargassum spinuligerum | |||
| Phlorofucofuroeckol A |  | Ecklonia kurome | [78] |
| Euphorbia stolonifera | [27] | ||
| Phlorofucofuroeckol B |  | Erica arborea | [79] |
| Bifuhalol |  | Bifurcaria bifurcata | [80] |
| Tetraphlorethols E |  | Cystophora retroflexa | [76] |
| Triphlorethol |  | Ecklonia cava | [81] |
| Extraction Conditions (Method, Replicates, LS, t, Sol, T) | Inhibitor | Substrate | Enzyme | Reaction Conditions | IC50 | Reference |
|---|---|---|---|---|---|---|
| SLE ×3 10 gsol/galgae 4 h Methanol (70% v/v) rt | Raw extract | Potato starch (1% w/w) | α-amylase (1 U/mL) | t = 10 min T = 37 °C | 0.052 mg/mL | [58] |
| p-NPG (6 mM) | α-glucosidase (0.2 mg/mL) | t = 10 min T = 37 °C | 0.52 mg/mL | |||
| SLE 5 gsol/galgae Overnight Chloroform rt | Raw extract | p-NPG (10 mM) | α- glucosidase (2 U/mL) | t = 10 min T = 28 °C | 194.2 µg/mL | [141] |
| SLE 5 gsol/galgae Overnight EtOH rt | 36.3 µg/mL | |||||
| SLE 5 gsol/galgae Overnight Acetone (70% v/v) rt | 13.0 µg/mL | |||||
| SLE (x3) 40 gsol/galgae 3 h Water 25 °C | Raw extract (10 mg/mL) | Wheat starch (1% w/v) | α-amylase (NS) | t = 10 min T = 25 °C | * 10% | [139] |
| p-NPG (5 mM) | α-glucosidase (0.1 U/mL) | t = 5 min T = 37 °C | * 8% | |||
| SLE (x3) 10 gsol/galgae Overnight MetOH NS | Raw extract | Potato starch (1% w/v) | α-amylase (0.4 mg/mL) | t = 2 h T = 37 °C | 0.1 µg/mL | [70] |
| p-NPG (2 mM) | α-glucosidase (10 mg/mL) | t = 2 h T = 37 °C | 19.0 µg/mL | |||
| SLE 20 gsol/galgae 30 min Water 80 °C | Raw extract | Wheat starch (1% w/w) | α-amylase (13 U/mL) | t = 10 min T = 25 °C | 1.34 µg | [22] |
| p-NPG (5 mM) | α-glucosidase (NS) | t = 20 min T = 37 °C | 0.24 µg | |||
| NS NS NS NS rt | Commercial raw extract | Corn starch (1% w/w) | α-amylase (0.2 U/mL) | t = 5 min T = 20 °C | 2.8 µg/mL | [142] |
| p-NPG (NS) | α-glucosidase (NS) | t = 20 min T = 37 °C | 5.0 µg/mL | |||
| SLE 12.5 gsol/galgae 90 min EtOH (50% v/v) 80 °C | Raw extract | p-NPG (0.7 mM) | α-glucosidase (0.1 mg/mL) | t = 15 min T = 37 °C | 38.0 µg/mL | [65] |
| Seaweed spp. | Extraction (Method, Replicates, LS, t, Solvent and T) | Inhibitor | Substrate | Enzyme | Reaction Conditions (t and T) | IC50 % Inhibition | Reference |
|---|---|---|---|---|---|---|---|
| AM FD SG SL PF UL | SLE ×2 10gsol/galgae 3 h MetOH (80% v/v) NS | Raw extracts (4 mg/mL) | Wheat starch 1% w/v | α-amylase NS | 10 min 37 °C | AM 6.4% FD 3.4% SG 76.1% SL 75.1% PF 6.6% UL 88.0% | [143] |
| p-NPG 1 mM | α-glucosidase 75 U/mg | 5 min 30 °C | AM 7.9% FD 18.4% SG 65.5% SL 56.3% PF 62.2% UL 82.4% | ||||
| Ecklonia stlonifera | SLE x0 4gsol/galgae 30 min Water NS | Raw extract | Wheat starch 20 mg/mL | α-glucosidase 10 U/mL | 5 min 37 °C | 0.026 mg/mL | [27] |
| Ecklonia cava | SLE x3 0.5gsol/galgae 10 days MetOH rt | Fucodiphloroethol G Dieckol 6,6′-Bieckol 7-Phloroeckol Phlorofucofuroeckol A | p-NPG 3 mM | α-amylase 1 U/mL | 5 min 37.5 °C | >0.5 mM | [144] |
| 0.1 mM | |||||||
| >0.5 mM | |||||||
| 0.3 mM | |||||||
| >0.5 mM | |||||||
| >0.5 mM | |||||||
| α-glucosidase 2 U/mL | 20 min 37.5 °C | 19.5 µM | |||||
| 10.8 µM | |||||||
| 22.2 µM | |||||||
| 49.5 µM | |||||||
| 19.7 µM | |||||||
| Ecklonia cava | SLE x3 NS 10 days MetOH 80% v/v rt | Dieckol | PNPG7 3 mM | α-amylase 0.7 U | 5 min 37.5 °C | 0.66 mM | [145] |
| α-glucosidase 1 U/mL | 0.24 mM | ||||||
| Ecklonia cava | SLE x3 50gsol/galgae 3 h EtOH 70% v/v rt | Phlorofucofuroeckol A | PNPG7 5 mM | α-amylase 100 U | t5 min rt | 6.3 µM | [146] |
| p-NPG 5 mM | α-glucosidase 0.7 U | 19.5 µM | |||||
| Ecklonia stolonifera and Eisenia bicyclis | SLE x3 20 gsol/galgae 1 h MetOH 40 °C | Phloroglucinol | p-NPG 2.5 mM | α-glucosidase 0.2 U/mL | 15 min 37 °C | 141.2 µM | [147] |
| Dioxinodehydroeckol | 34.6 µM | ||||||
| Eckol | 11.8 µM | ||||||
| Phlorofucofuroeckol A | 1.4 µM | ||||||
| Dieckol | 1.6 µM | ||||||
| Eisenia bicyclis | SLE x3 2 gsol/galgae 3 h MetOH rt | Phloroglucinol tetramer (1 mM) | Wheat starch 1% w/v | α-amylase NS | 15 min 37 °C | 96.2% | [67] |
| Eckol (1 mM) | 86.7% | ||||||
| Dieckol (1 mM) | 76.0% | ||||||
| Eisenia bicclis | SLE ×3 40 gsol/galgae 3 h EtOH 25 °C | Methanolic extract n-Hexane fraction Dichloromethane fraction Ethyl acetate fraction n-Butanol fraction Water fraction Fucofuroeckol A Dioxinodehydroeckol | Potato starch 1% w/v | α-amylase 0.4 mg/mL | 2 h 37.5 °C | 0.5 µg/mL | [148] |
| 3.5 µg/mL | |||||||
| 0.3 µg/mL | |||||||
| 48.1 ng/mL | |||||||
| 0.2 µg/mL | |||||||
| 1.9 µM | |||||||
| 51.6 nM | |||||||
| 93.3 nM | |||||||
| p-NPG 3 mM | α- glucosidase 0.4 mg/mL | 20 min 37.5 °C | >500 µg/mL | ||||
| >500 µg/mL | |||||||
| 39.98 g/mL | |||||||
| 2.9 µg/mL | |||||||
| 4.6 µg/mL | |||||||
| >500 µg/mL | |||||||
| 42.9 µM | |||||||
| 0.47 mM | |||||||
| Ishige foliacea | SLE x3 40 gsol/galgae 3 h MetOH 80% v/v 25 °C | Octaphlorethol A | p-NPG 5 mM | α- glucosidase 0.7 U/mL | 5 min rt | 0.11 mM | [149] |
| Ishige okamurae | SLE NS NS MetOH 80% v/v rt | Diphlorethohydroxycarmalol | p-NPG 5 mM | α-amylase 0.7 U | 5 min rt | 0.53 mM | [73] |
| α-glucosidase 0.7 U/mL | 0.16 mM | ||||||
| Sargassum ringgoldianum | SLE x3 NS NS MetOH 80% v/v NS | Raw extract | PNPG7 5 mM | α-amylase 100 U | 5 min rt | 0.18 mg/dL | [150] |
| p-NPG 5 mM | α-glucosidase 0.7 U | 0.12 mg/dL | |||||
| p-NPG 5 mM | α-glucosidase NS | 20 min 37 °C | 0.24 µg | ||||
| SF SA SS SP SD | SLE x3 40 gsol/galgae 30 min EtAc 90 °C | Raw extracts | Wheat starch 1% w/v) | α-amylase 13 U/mL | 10 min 25 °C | SF 27.0% SA 55.0% SS 30.0% SP 42.0% SD 36.0% | [151] |
| p-NPG 5 mM | α-glucosidase 0.7 U | 5 min 25 °C | SF 37.0% SA 65.0% SS 40.0% SP 52.0% SD 46.0% | ||||
| Triticum aestivum | SLE x2 40 gsol/galgae 1 h EtOH 25 °C | (2-(4-(3,5-dihydroxyphenoxy)-3,5-dihydroxyphenoxy) benzene-1,3,5-triol) | Amylopectin 1% w/v | α-amylase 1 µM | 5 min 37 °C | 3.2 µg/mL | [152] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gisbert, M.; Franco, D.; Sineiro, J.; Moreira, R. Antioxidant and Antidiabetic Properties of Phlorotannins from Ascophyllum nodosum Seaweed Extracts. Molecules 2023, 28, 4937. https://doi.org/10.3390/molecules28134937
Gisbert M, Franco D, Sineiro J, Moreira R. Antioxidant and Antidiabetic Properties of Phlorotannins from Ascophyllum nodosum Seaweed Extracts. Molecules. 2023; 28(13):4937. https://doi.org/10.3390/molecules28134937
Chicago/Turabian StyleGisbert, Mauro, Daniel Franco, Jorge Sineiro, and Ramón Moreira. 2023. "Antioxidant and Antidiabetic Properties of Phlorotannins from Ascophyllum nodosum Seaweed Extracts" Molecules 28, no. 13: 4937. https://doi.org/10.3390/molecules28134937
APA StyleGisbert, M., Franco, D., Sineiro, J., & Moreira, R. (2023). Antioxidant and Antidiabetic Properties of Phlorotannins from Ascophyllum nodosum Seaweed Extracts. Molecules, 28(13), 4937. https://doi.org/10.3390/molecules28134937