
Anti-Inflammatory Effects of Dietary Polyphenols through Inhibitory Activity against Metalloproteinases

 ,
,  , and
, and
Abstract
:1. Introduction
2. Roles of MMPs in Human Inflammatory Diseases
2.1. Rheumatoid Arthritis and Osteoarthritis
2.2. Alzheimer’s Disease
2.3. Other Inflammatory Diseases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference [PMID] | Major Findings | |
|---|---|---|
| OA | Zeng, et al. [19] | A meta-analysis of 10 studies with 458 OA patients and 295 healthy controls indicated that the protein levels of MMP1, MMP2, and MMP9 were higher in patients with OA patients than those in the control group. Asian OA patients showed the higher protein levels of MMP1 and MMP2 compared to the controls, while Caucasians did not show such differences. In both populations, the MMP9 protein levels in OA were higher than in controls. The MMP1 and MMP9 protein levels in the synovial joint fluid were higher in OA patients than in controls. These results suggest an association between the OA pathogenesis and the increased levels of MMP1, MMP2, and MMP9 proteins. |
| Sieghart et al. [29] | Interleukin (IL)1β-stimulation of primary fibroblast-like synoviocytes from OA patients activated several c-Jun N-terminal kinase (JNK) mitogen activated protein kinases (MAPKs), including extracellular signal-regulated kinase (ERK1/2), JNK, heat shock protein 27 (Hsp27), and p38 MAPK (p38). The mRNA levels of MMP2 and MMP14 were upregulated in IL1β-stimulated cells. H2S, which has anti-inflammatory properties [30], attenuated these effects as well as IL1β-induced secretion of IL6, IL8, and RANTES. | |
| RA | Zhou, et al. [31] | Serum MMP3 levels in RA patients with moderate and severe disease activity were higher than in patients with stable RA. Levels in mild RA were not different from those in the stable RA and the healthy control group. Levels decreased in patients treated with certolizumab pegol, a tumor necrosis factor-α (TNFα) inhibitor drug for the treatment of RA. The findings suggest MMP3 as a useful serum biomarker for RA. |
| Crowley et al. [32] Behera et al. [33] | Lyme disease, a form of chronic inflammatory arthritis, is transmitted by ticks (Borrelia burgdorferi) and patients often develop Lyme arthritis (LA). Tick infection in primary human chondrocytes caused upregulation of gene expression of MMP1, MMP3, MMP10, MMP13, MMP19, and TIMP1. Protein levels of MMP10, MMP13, and TIMP1 were increased by 72 h post infection [33]. Autoantigens in antibiotic-refractory LA were found to include MMP10 and its peptide. The levels of serum MMP10 autoantibodies in LA patients, especially antibiotic-refractory LA patients, and those of MMP10 protein in joint fluid were significantly increased. The levels of anti-MMP10 autoantibodies were correlated positively with synovial pathology, suggesting MMP10 as a useful pathological biomarker for LA. | |
| Yu et al. [34] | Seventy RA patients were randomly divided into two groups, and a treatment group received moxibustion, a traditional Oriental therapy with thermal stimulation by burning herbs [35], in addition to the drugs given to the control group. The decreases in the serum levels of IL1β, TNFα, MMP1, MMP3, and vascular endothelial growth factor (VEGF) in the moxibustion-treated group were more pronounced compared to the control group. The serum MMP1 and MMP3 levels are suggested to be useful biomarkers of cartilage and bone erosion in RA patients, and the efficacy of moxibustion on the clinical RA symptoms may be due to its contribution to the downregulation of MMP1 and MMP3. | |
| Wang et al. [36] | The expression levels of MMP3, MMP9, and MMP13 in fibroblast-like synoviocytes derived from RA patients were increased by transfection of mimic RNA of miR-145-5p, a microRNA suggested for involvement in RA development and progression. miR-145-5p inhibitor reduced these levels. The increase in MMP9 levels and the enhancement of nuclear factor (NF)-κB p65 nuclear translocation caused by miR-145-5p overexpression were attenuated by the NF-κB inhibitor, indicating the involvement of NF-κB pathway activation in MMP9 expression, suggesting that modulation of miR-145-5p is useful for the treatment of RA associated with MMPs secretion via activation of the NF-κB pathway. | |
| Inflammatory bowel disease (IBD) | Majster et al. [37] | Analysis of inflammation-related proteins in the serum collected from IBD patients revealed a significant increase in 21 proteins including IL6 and MMP10 in the serum and a significant decrease in 4 proteins compared to the control sample. IL6 and MMP10 were also significantly increased in saliva of these patients and correlated with their expressions in the serum. These findings suggest that the oral cavity reflects ongoing intestinal inflammation, and that saliva can be used as a non-invasive source of IBD biomarkers. |
| Soomro et al. [38] | In the search for a biomarker for IBD, such as CD and ulcerative colitis (UC), MMP9 and MMP12 levels in stools were found to increase significantly in both UC and CD patients. A longitudinal cohort study of 50 patients with UC showed a strong correlation of diagnostic severity of IBD with fibrinogen, MMP8, short peptidoglycan recognition protein, and TIMP2, suggesting that fecal MMP8, MMP9, and MMP12 levels are useful biomarkers for the early detection of IBD. | |
| Buchbender et al. [39] | mRNA expression analysis of inflammatory proteins in gingival pocket biofilms from IBD patients revealed that IL10 mRNA expression levels were higher in both CD and UC patients, and MMP7 mRNA expression levels were significantly higher in CD samples compared to the controls. In contrast, MMP7 mRNA expression levels in UC patients were not different from those in controls. The relationship between the stage of IBD progression and the mRNA expression levels of IL10 and MMP7 was not clear. These findings suggest that IL10 and MMP7 expression levels in oral biofilm may be a useful non-invasive biomarker for IBD. | |
| Lakatos et al. [40] Coufal et al. [41] | When serum antigen levels of MMP2, MMP7, MMP9, TIMP1, and TIMP2 were determined in 23 UC patients and 25 CD patients in comparison with 10 healthy subjects, the levels of MMP9, TIMP1, and TIMP2 were significantly higher in UC and CD, and the levels of MMP7 were higher in CD [40]. MMP9 and TIMP1 levels were positively correlated with disease activity in IBD, while MMP2 and TIMP2 levels were inversely correlated with CD activity. In contrast, lower serum levels of MMP9 and higher serum levels of MMP14 were found in patients with IBD (n = 85), UC (n = 36) and CD (n = 20) compared to healthy subjects (n = 25) [41]. A discrepancy found for MMP9 suggests the need for further studies with larger sample sizes. | |
| AD | Boström et al. [42] | Analysis of 92 neuroinflammatory proteins in the CSF of patients with neurodegenerative diseases such as AD and frontotemporal dementia revealed that MMP10 levels were markedly increased in the CSF of both AD and mild cognitive impairment/AD patients significantly increased. In stratification by patient group and medical facility, a trend toward higher MMP10 levels was observed in all three neurodegenerative disease groups. These results suggest that elevated MMP10 levels are a common feature of AD and frontotemporal dementia, although they have different symptoms due to the different types of inflammatory proteins secreted into the CSF. |
| Sorrentino, et al. [23] | Analysis of changes in the expression of pro- and/or anti-inflammatory cytokines in brain homogenate samples from AD patients and control subjects revealed increased expression of IL4, IL6, IL13, CCL17, MMP7, and CXCL13 in AD patients compared to control subjects. No significant differences were found in MMP1, MMP8, and MMP9 levels between AD patients and the control. Among the three patient clusters divided by hierarchical cluster analysis, in the class 3 group of AD, which was characterized by the low levels of amyloid β (Aβ) peptides in the brain and the longest disease duration, MMPs levels were increased. This group showed the lowest levels of almost all the molecules tested except for MMP8, MMP9, CX3CL1, and LCN2. These findings suggest that neuroinflammatory molecules such as MMP7 and CXCL13 are useful as biomarkers for the AD diagnosis. | |
| Rhinosinusitis | Chen, et al. [43] | It is known that IL17A is markedly elevated in chronic rhinosinusitis with nasal polyps (CRSwNP). The number of IL17A-producing CD8+ T cells was increased in the CRSwNP group compared to the CR group without nasal polyps and the control group. The mRNA and protein expression levels of MMP7 and MMP9 were significantly increased in the CRSwNP group. In addition, exposure of primary human nasal epithelial cells to IL17A increased MMP9 levels. Activation of the NF-κB pathway was found to be involved in the IL17A-induced increase in MMP9 levels. These findings suggest that IL17A-induced MMP9 in the pathogenesis and tissue remodeling of CRSwNP is caused by IL17-stimulated activation of the NF-κB pathway. |
| Wang, et al. [44] | Inflammatory cytokines and MMPs are known to be elevated in the tissue of CRSwNP. Sixty patients were divided into two groups to receive budesonide or placebo for 14 days. The drug treatment reduced the polyp size compared with placebo and improved symptoms. The drug reduced the expression of the pro-inflammatory cytokines IL5 and eotaxin, and increased TGFβ1 and IL10 expressions in the polyp samples. Budesonide also decreased indices of remodeling in these samples including albumin, MMP2, MMP7, MMP8, and MMP9, but increased collagen and TIMP1, TIMP2, and TIMP4 levels. | |
| LA | Crowley et al. [32] | An immunogenic HLA-DR-presented peptide (T-cell epitope) derived from the source protein MMP10 was identified in the synovium of a patient with antibiotic-refractory LA. The level of MMP10 autoantibodies in the serum of LA patients, especially antibiotic-refractory LA patients and the amount of MMP10 protein in the joint fluid were significantly increased regardless of antibiotic-responsive or antibiotic-refractory. A positive correlation was found between anti-MMP10 autoantibodies and synovial pathology. These findings suggest MMP10 as a useful pathologic biomarker for LA. |
3. Polyphenol’s Inhibitory Activity against MMPs in Inflammatory Diseases
3.1. OA and RA
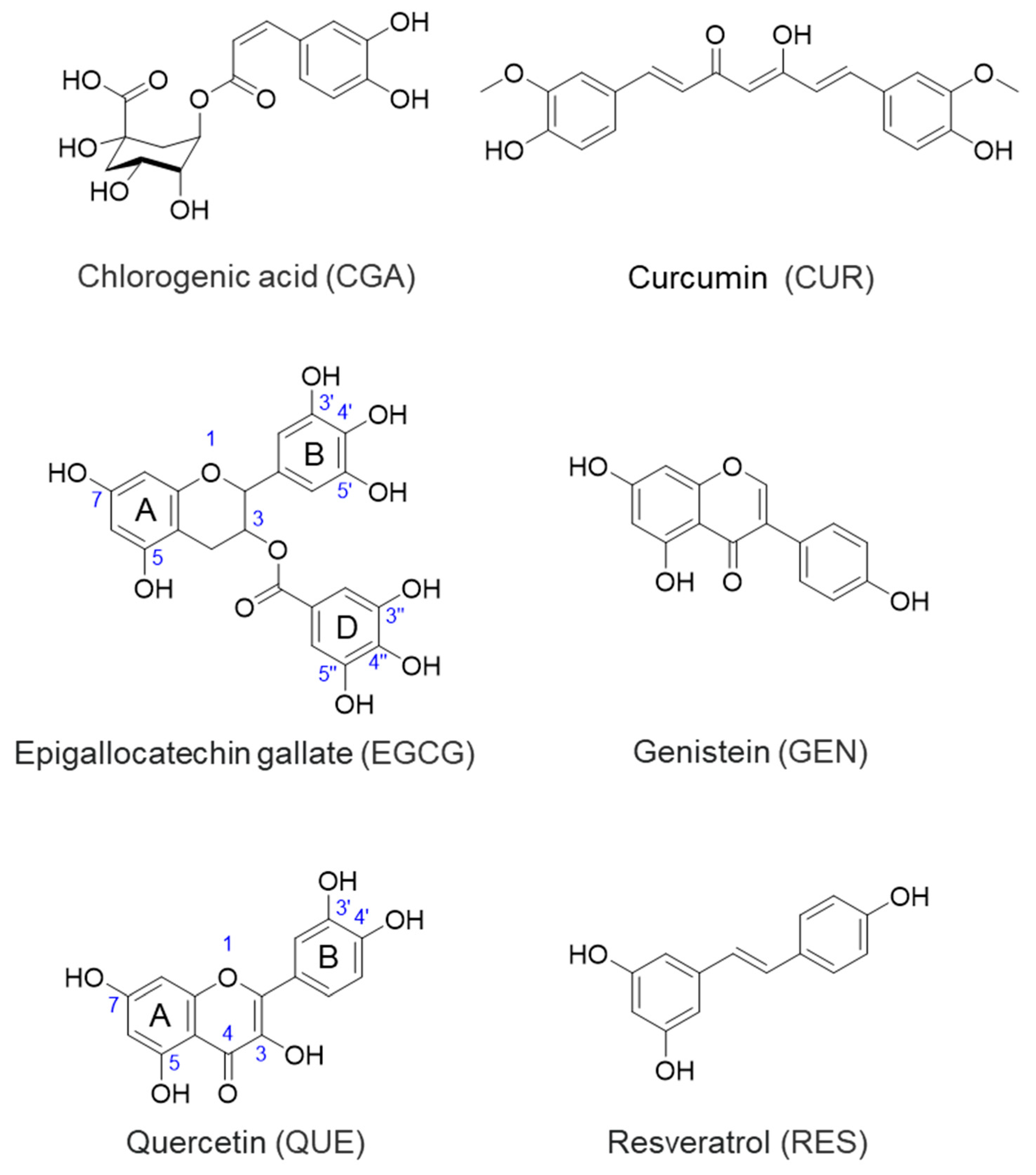
3.1.1. CGA
3.1.2. CUR
3.1.3. EGCG
3.1.4. GEN
3.1.5. QUE
3.1.6. RES
3.2. Asthma
EGCG
3.3. AD
3.3.1. GEN
3.3.2. QUE
3.3.3. RES
3.4. Other Inflammatory Diseases
3.4.1. CUR
3.4.2. EGCG
3.4.3. QUE
3.4.4. RES
4. Mechanistic Considerations on Polyphenol’s Inhibition of MMPs
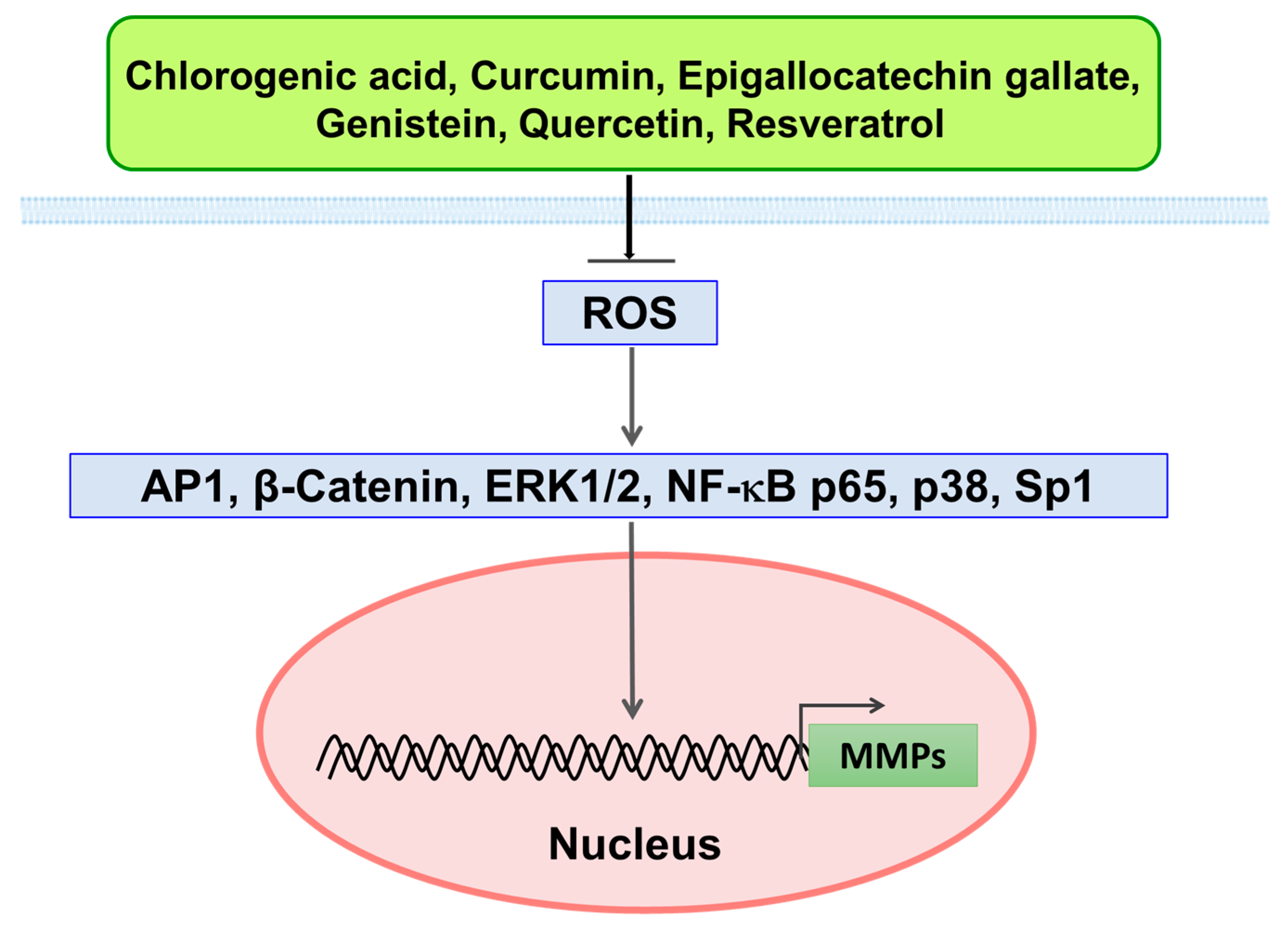
4.1. Inhibitory Effect of Polyphenols on MMPs via ROS-Mediated Signaling Pathways
4.2. MDA of Interactions between Polyphenols and MMPs
4.2.1. Interaction between MMP1 and Polyphenols
CUR, EGCG, QUE, and RES
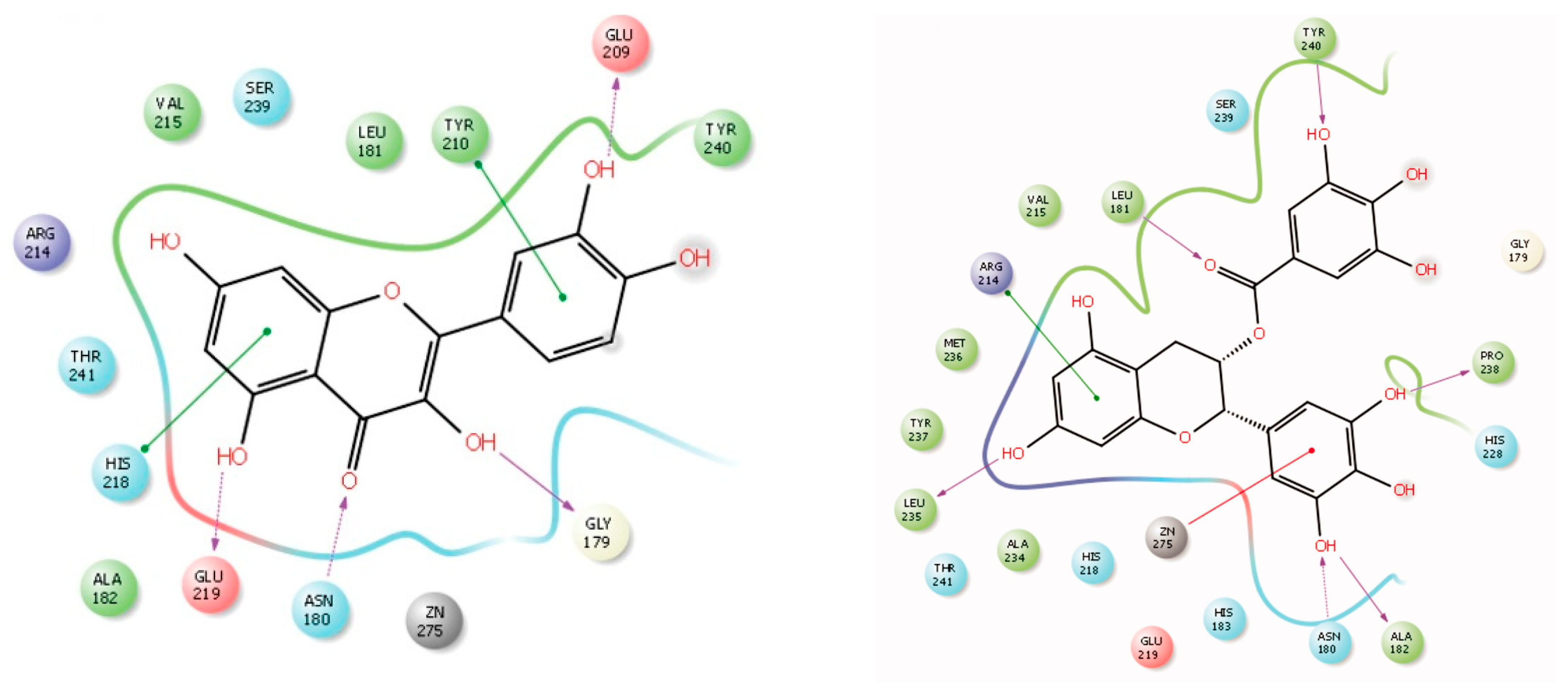
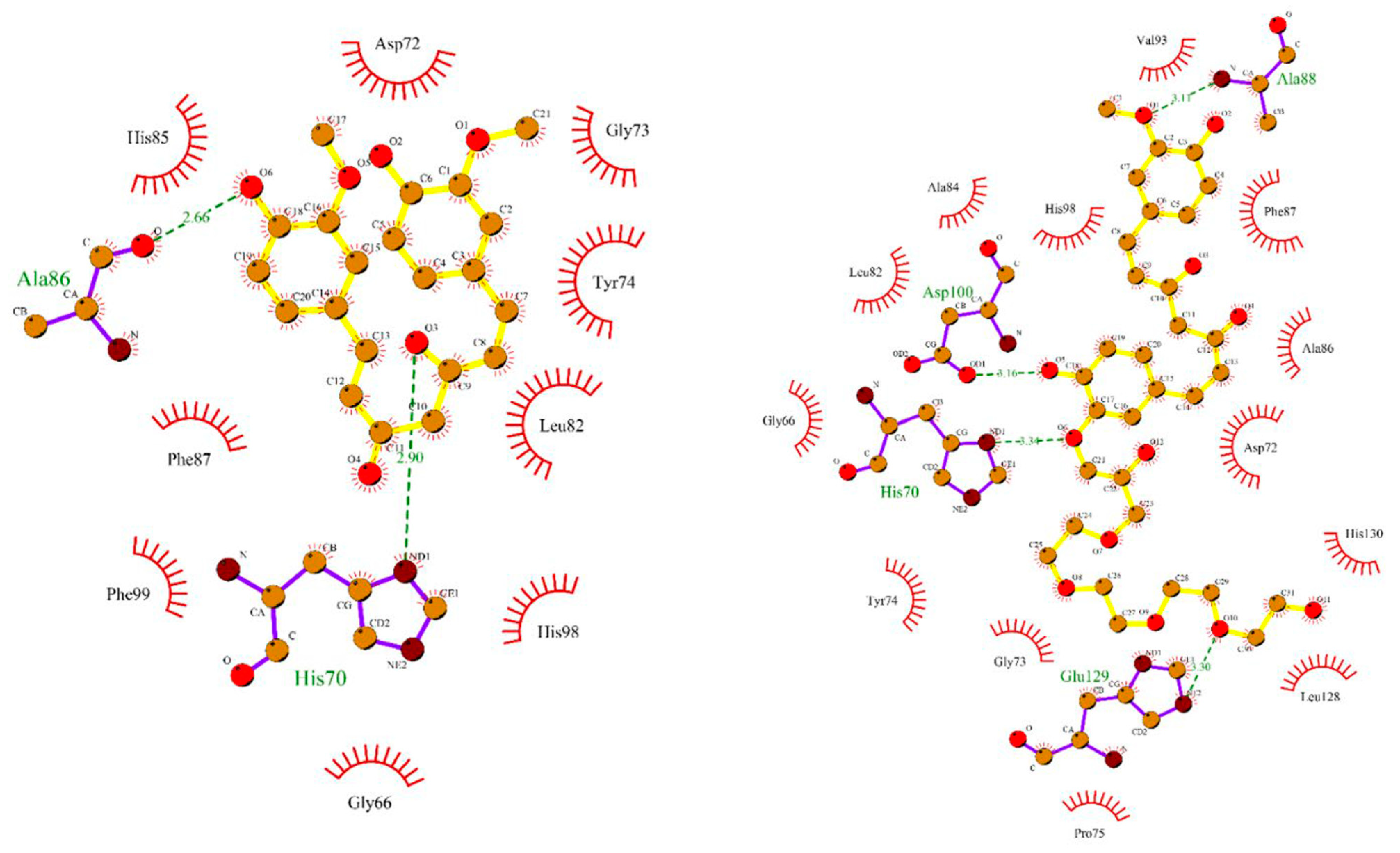
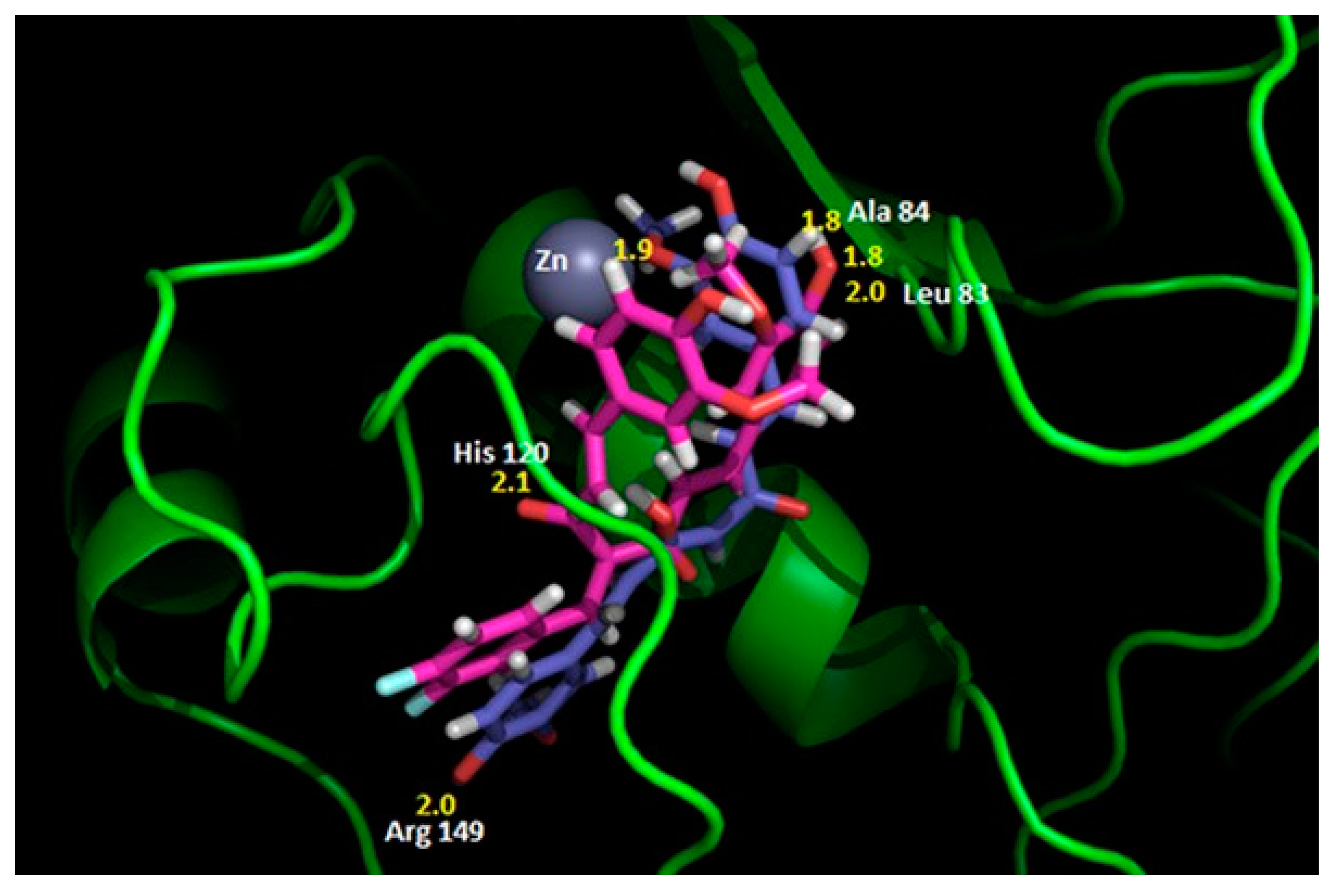
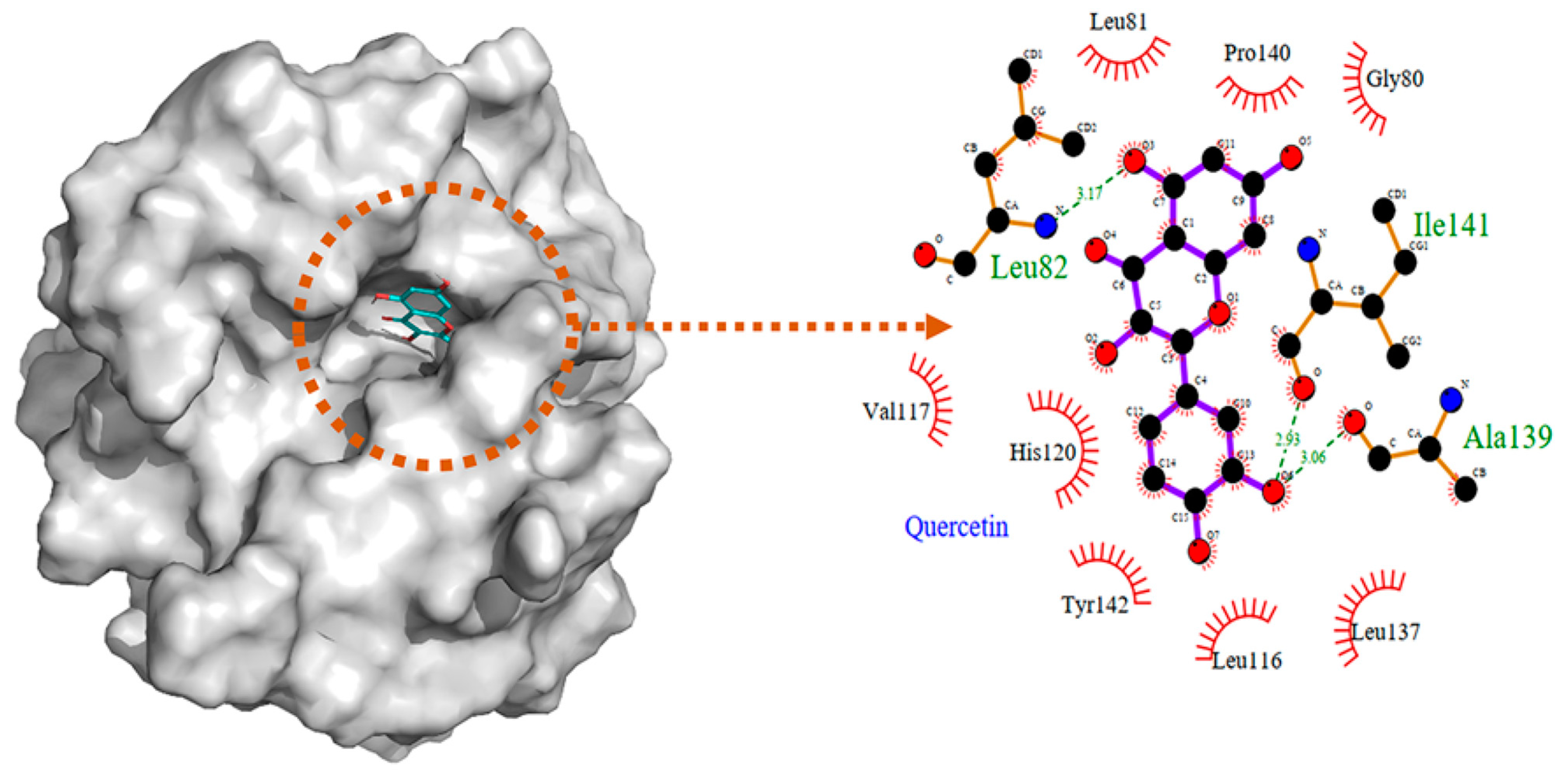
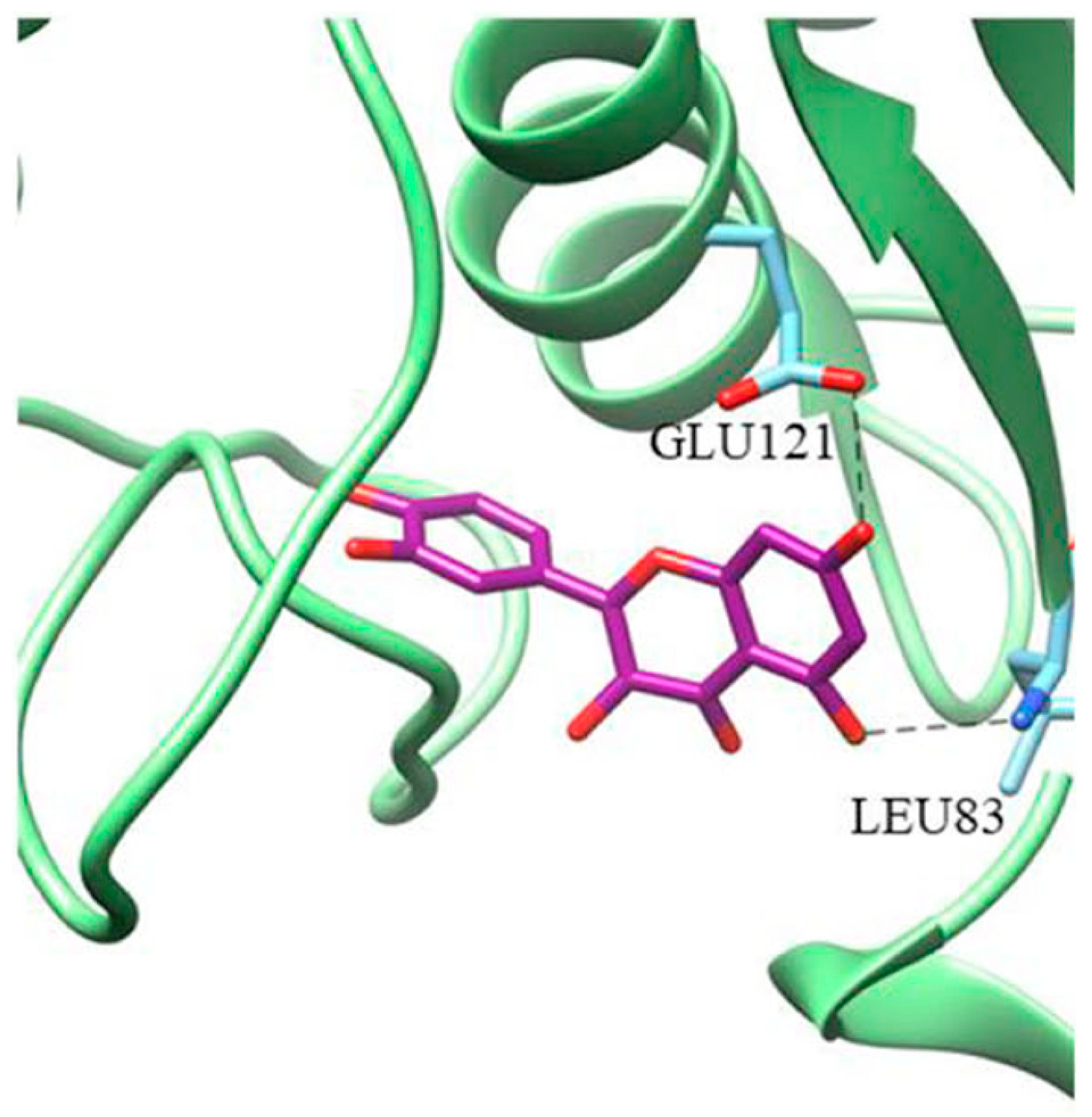
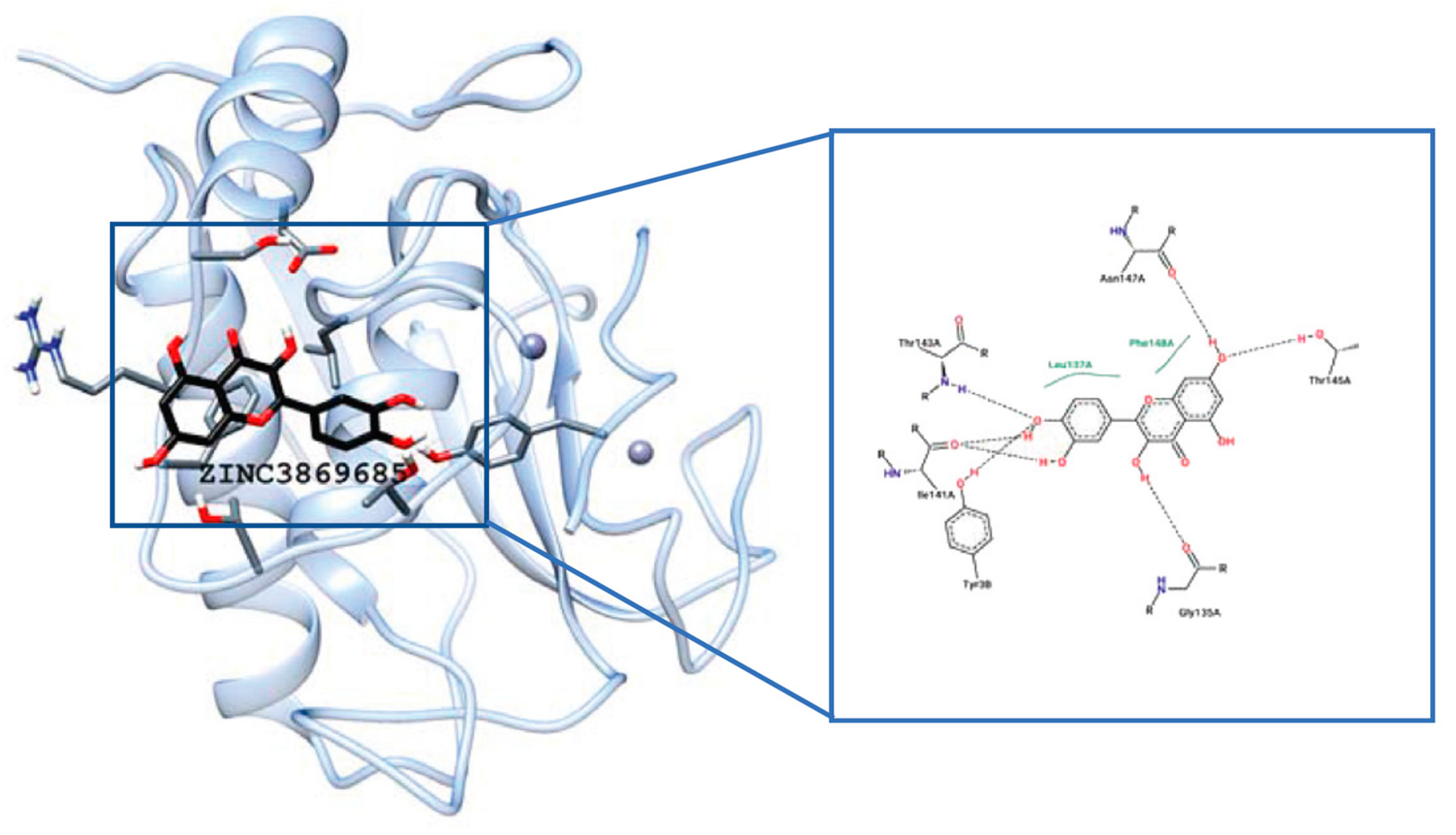
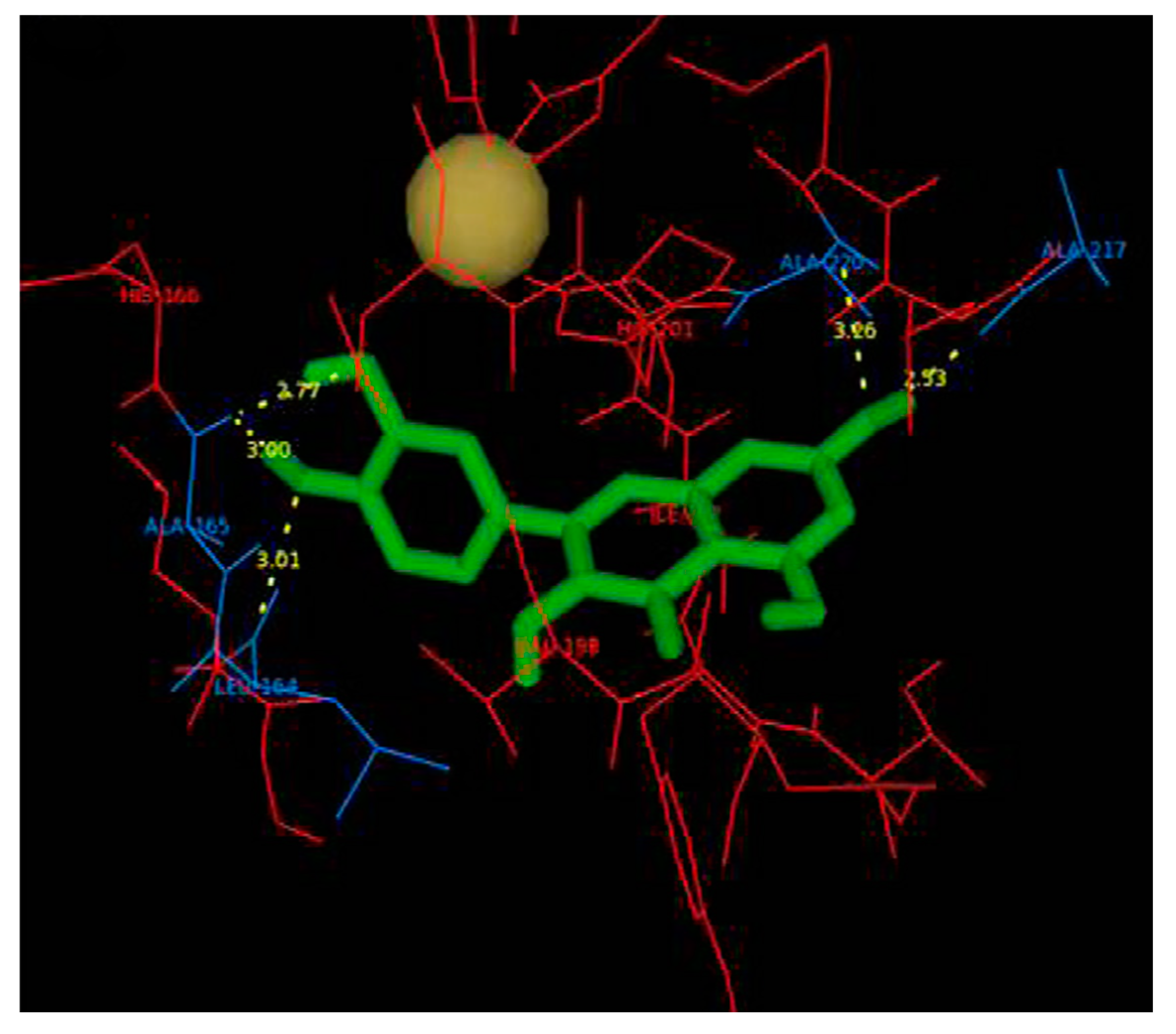
4.2.2. Interaction between MMP2 and Polyphenols
CGA
CUR
EGCG
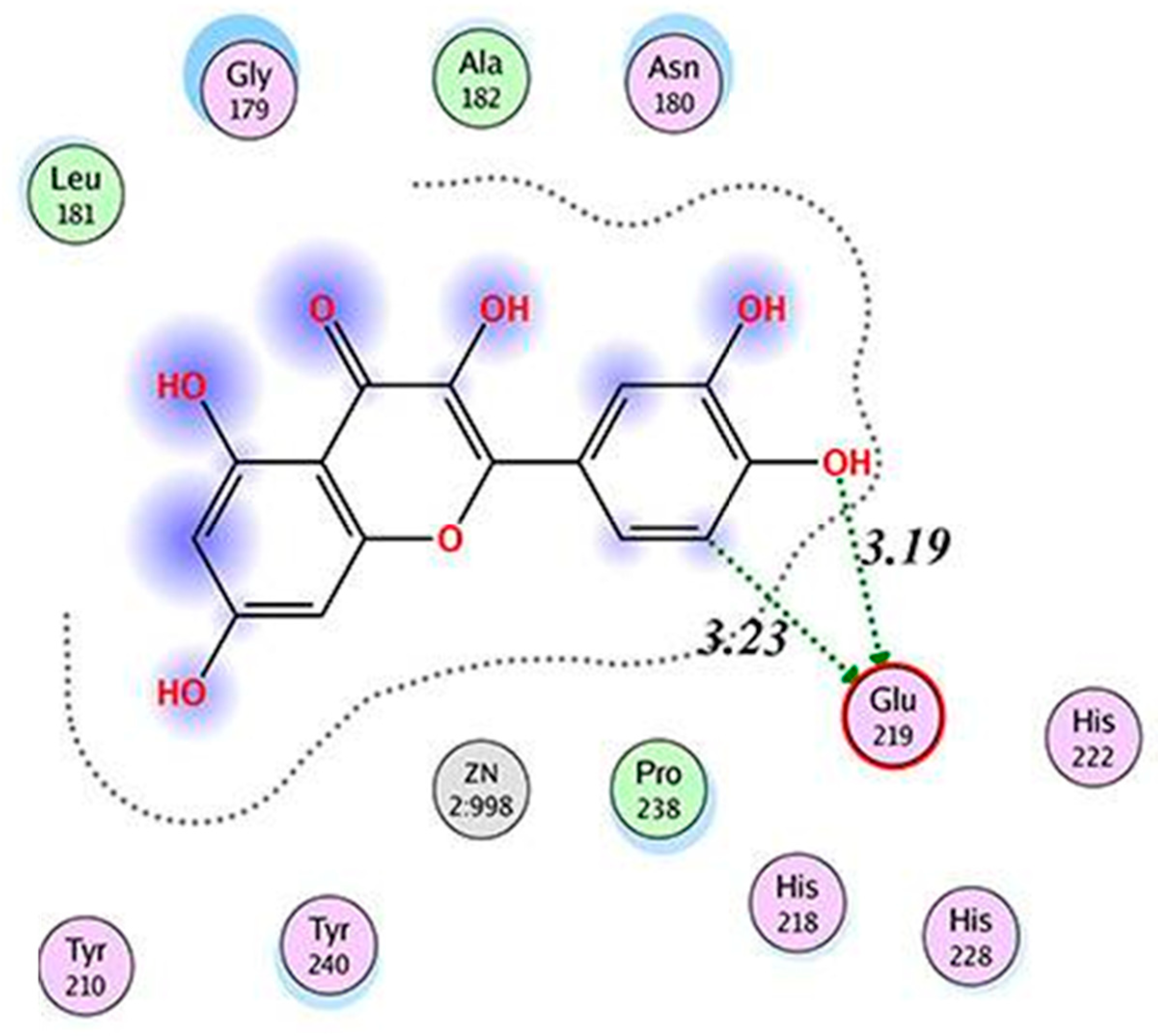
QUE
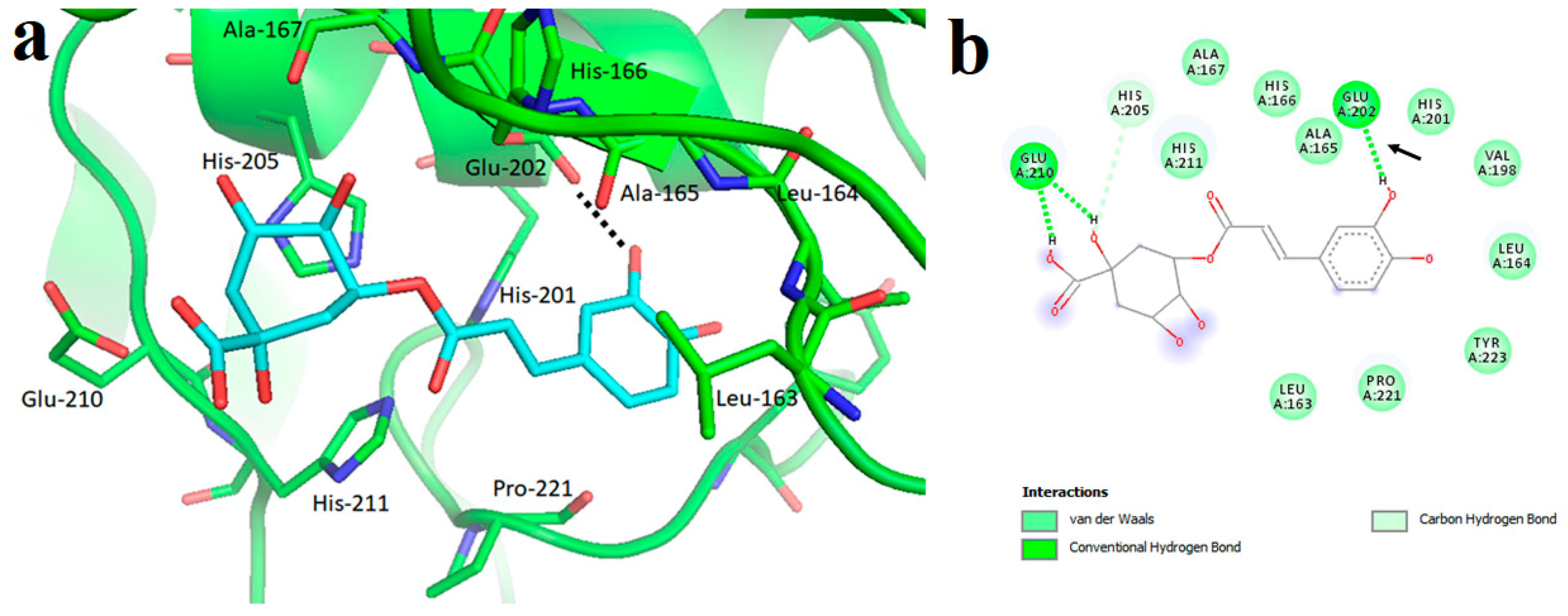
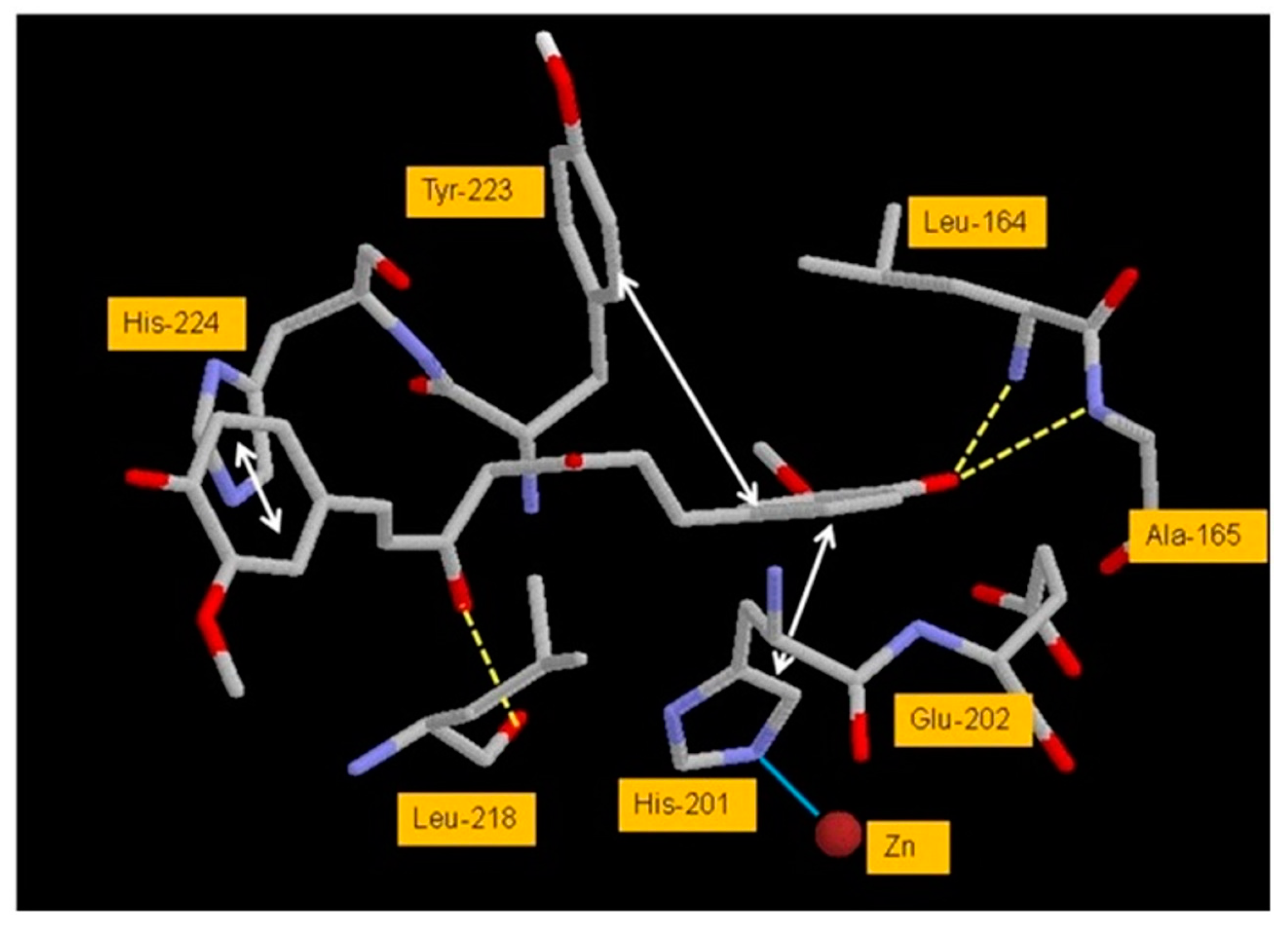
4.2.3. Interaction between MMP3 and Polyphenols
CUR
QUE
4.2.4. Interaction between MMP9 and Polyphenols
CUR
EGCG
QUE
4.2.5. Interaction between MMP14 and Polyphenols
EGCG
5. Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| Adamts 5 | A disintegrin and metalloproteinase with thrombospondin motifs |
| AGE | Advanced glycation end-products |
| AP1 | Activator protein 1 |
| CCL | C-C Motif Ligand |
| CD | Crohn’s disease |
| CGA | Chlorogenic acid |
| CNS | Central nervous system |
| COX-2 | Cyclooxygenase 2 |
| CRSwNP | Chronic rhinosinusitis with nasal polyp |
| CSF | Cerebrospinal fluid |
| CUR | Curcumin |
| CXCL | C-X-C motif chemokine ligand |
| DenCUR | Dendrosome CUR |
| EGC | Epigallocatechin |
| EGCG | Epigallocatechin-3-O-gallate |
| EGF | Epidermal growth factor |
| ERK | Extracellular signal-regulated kinase |
| GEN | Genistein |
| Hsp27 | Heat shock protein 27 |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IL | Interleukin |
| JNK | c-Jun N-terminal kinase |
| LA | Lyme arthritis |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen activated protein kinase |
| MDA | Molecular docking analysis |
| MIF | Migration inhibitory factor |
| MMP | Matrix metalloproteinase |
| MCP | Monocyte chemotactic protein |
| NF | Nuclear factor |
| NOS | Nitric oxide synthase |
| OA | Osteoarthritis |
| p38 | p38 mitogen-activated protein kinases |
| PGE2 | Prostaglandin E2 |
| QUE | Quercetin |
| RA | Rheumatoid arthritis |
| ROS | Reactive oxygen species |
| Sirt1 | Silencing information regulator 2-related enzyme 1 |
| Sp1 | Specificity protein 1 |
| TNFα | Tumor necrosis factor-α |
| UC | Ulcerative colitis |
| VEGF | Vascular endothelial growth factor |
References
- Abenavoli, L.; Larussa, T.; Corea, A.; Procopio, A.C.; Boccuto, L.; Dallio, M.; Federico, A.; Luzza, F. Dietary Polyphenols and Non-Alcoholic Fatty Liver Disease. Nutrients 2021, 13, 494. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Suzuki, T.; Ohishi, T.; Isemura, M.; Nakamura, Y.; Unno, K. Effects of Epigallocatechin-3-Gallate on Matrix Metalloproteinases in Terms of Its Anticancer Activity. Molecules 2023, 28, 525. [Google Scholar] [CrossRef]
- Shamsi, A.; Anwar, S.; Shahbaaz, M.; Mohammad, T.; Alajmi, M.F.; Hussain, A.; Hassan, I.; Ahmad, F.; Islam, A. Evaluation of Binding of Rosmarinic Acid with Human Transferrin and Its Impact on the Protein Structure: Targeting Polyphenolic Acid-Induced Protection of Neurodegenerative Disorders. Oxid. Med. Cell Longev. 2020, 2020, 1245875. [Google Scholar] [CrossRef]
- Shahwan, M.; Alhumaydhi, F.; Ashraf, G.M.; Hasan, P.M.Z.; Shamsi, A. Role of Polyphenols in Combating Type 2 Diabetes and Insulin Resistance. Int. J. Biol. Macromol. 2022, 206, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Majrashi, T.A.; Alshehri, S.A.; Alsayari, A.; Muhsinah, A.B.; Alrouji, M.; Alshahrani, A.M.; Shamsi, A.; Atiya, A. Insight into the Biological Roles and Mechanisms of Phytochemicals in Different Types of Cancer: Targeting Cancer Therapeutics. Nutrients 2023, 15, 1704. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, H.; Yin, C.; Lan, X.; Wu, L.; Du, X.; Griffiths, H.R.; Gao, D. Adipokines, Hepatokines and Myokines: Focus on Their Role and Molecular Mechanisms in Adipose Tissue Inflammation. Front. Endocrinol. 2022, 13, 873699. [Google Scholar] [CrossRef]
- Sarkar, S. Microglial Ion Channels: Key Players in Non-Cell Autonomous Neurodegeneration. Neurobiol. Dis. 2022, 174, 105861. [Google Scholar] [CrossRef] [PubMed]
- Videmark, A.N.; Christensen, I.J.; Feltoft, C.L.; Villadsen, M.; Borg, F.H.; Jørgensen, B.M.; Bojesen, S.E.; Kistorp, C.; Ugleholdt, R.; Johansen, J.S. Combined Plasma C-Reactive Protein, Interleukin 6 and YKL-40 for Detection of Cancer and Prognosis in Patients with Serious Nonspecific Symptoms and Signs of Cancer. Cancer Med. 2023, 12, 6675–6688. [Google Scholar] [CrossRef]
- Shamsi, F.B.; Anwar, A.; Lail, R.A.; Bukhari, M.H.; Naseem, N.; Nagi, A.H. Immunohistochemical Expression of Matrix Metalloproteinase-1 (MMP-1) in Different Types Of Breast Carcinoma And Its Comparison With ER/PER and HER2/Neu. J. Ayub Med. Coll. Abbottabad 2022, 34, 12–16. [Google Scholar] [CrossRef]
- Kumar, L.; Bisen, M.; Khan, A.; Kumar, P.; Patel, S.K.S. Role of Matrix Metalloproteinases in Musculoskeletal Diseases. Biomedicines 2022, 10, 2477. [Google Scholar] [CrossRef]
- Li, R.-L.; Duan, H.-X.; Liang, Q.; Huang, Y.-L.; Wang, L.-Y.; Zhang, Q.; Wu, C.-J.; Liu, S.-Q.; Peng, W. Targeting Matrix Metalloproteases: A Promising Strategy for Herbal Medicines to Treat Rheumatoid Arthritis. Front. Immunol. 2022, 13, 1046810. [Google Scholar] [CrossRef]
- Hayakawa, S.; Ohishi, T.; Oishi, Y.; Isemura, M.; Miyoshi, N. Contribution of Non-Coding RNAs to Anticancer Effects of Dietary Polyphenols: Chlorogenic Acid, Curcumin, Epigallocatechin-3-Gallate, Genistein, Quercetin and Resveratrol. Antioxidants 2022, 11, 2352. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, T.; Miyoshi, N.; Mori, M.; Sagara, M.; Yamori, Y. Health Effects of Soy Isoflavones and Green Tea Catechins on Cancer and Cardiovascular Diseases Based on Urinary Biomarker Levels. Molecules 2022, 27, 8899. [Google Scholar] [CrossRef] [PubMed]
- Keyszer, G.; Lambiri, I.; Nagel, R.; Keysser, C.; Keysser, M.; Gromnica-Ihle, E.; Franz, J.; Burmester, G.R.; Jung, K. Circulating Levels of Matrix Metalloproteinases MMP-3 and MMP-1, Tissue Inhibitor of Metalloproteinases 1 (TIMP-1), and MMP-1/TIMP-1 Complex in Rheumatic Disease. Correlation with Clinical Activity of Rheumatoid Arthritis versus Other Surrogate Markers. J. Rheumatol. 1999, 26, 251–258. [Google Scholar] [PubMed]
- Mahmoud, R.K.; El-Ansary, A.K.; El-Eishi, H.H.; Kamal, H.M.; El-Saeed, N.H. Matrix Metalloproteinases MMP-3 and MMP-1 Levels in Sera and Synovial Fluids in Patients with Rheumatoid Arthritis and Osteoarthritis. Ital. J. Biochem. 2005, 54, 248–257. [Google Scholar]
- Tchetverikov, I.; Lard, L.R.; DeGroot, J.; Verzijl, N.; TeKoppele, J.M.; Breedveld, F.C.; Huizinga, T.W.J.; Hanemaaijer, R. Matrix Metalloproteinases-3, -8, -9 as Markers of Disease Activity and Joint Damage Progression in Early Rheumatoid Arthritis. Ann. Rheum. Dis. 2003, 62, 1094–1099. [Google Scholar] [CrossRef]
- Waszczykowski, M.; Bednarski, I.; Narbutt, J.; Waszczykowska, E.; Lesiak, A.; Fabiś, J. Interleukin-18, Interleukin-20, and Matrix Metalloproteinases (MMP-1, MMP-3) as Markers of Psoriatic Arthritis Disease Severity and Their Correlations with Biomarkers of Inflammation and Turnover of Joint Cartilage. Postep. Dermatol. Alergol. 2020, 37, 1001–1008. [Google Scholar] [CrossRef]
- Ingale, D.; Kulkarni, P.; Electricwala, A.; Moghe, A.; Kamyab, S.; Jagtap, S.; Martson, A.; Koks, S.; Harsulkar, A. Synovium-Synovial Fluid Axis in Osteoarthritis Pathology: A Key Regulator of the Cartilage Degradation Process. Genes 2021, 12, 989. [Google Scholar] [CrossRef]
- Zeng, G.Q.; Chen, A.B.; Li, W.; Song, J.H.; Gao, C.Y. High MMP-1, MMP-2, and MMP-9 Protein Levels in Osteoarthritis. Genet. Mol. Res. 2015, 14, 14811–14822. [Google Scholar] [CrossRef]
- Testa, G.; Gamba, P.; Badilli, U.; Gargiulo, S.; Maina, M.; Guina, T.; Calfapietra, S.; Biasi, F.; Cavalli, R.; Poli, G.; et al. Loading into Nanoparticles Improves Quercetin’s Efficacy in Preventing Neuroinflammation Induced by Oxysterols. PLoS ONE 2014, 9, e96795. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.; Liu, F.; Meng, M.; Zhang, L.; Gordon, M.L.; Wang, Y.; Cai, L.; Zhang, N. Elevated Matrix Metalloproteinase-9 Levels in Neuronal Extracellular Vesicles in Alzheimer’s Disease. Ann. Clin. Transl. Neurol. 2020, 7, 1681–1691. [Google Scholar] [CrossRef]
- Duits, F.H.; Hernandez-Guillamon, M.; Montaner, J.; Goos, J.D.C.; Montañola, A.; Wattjes, M.P.; Barkhof, F.; Scheltens, P.; Teunissen, C.E.; van der Flier, W.M. Matrix Metalloproteinases in Alzheimer’s Disease and Concurrent Cerebral Microbleeds. J. Alzheimers. Dis. 2015, 48, 711–720. [Google Scholar] [CrossRef]
- Sorrentino, S.; Ascari, R.; Maderna, E.; Catania, M.; Ghetti, B.; Tagliavini, F.; Giaccone, G.; Di Fede, G. Microglial Heterogeneity and Its Potential Role in Driving Phenotypic Diversity of Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 2780. [Google Scholar] [CrossRef]
- Martinez-Castillo, M.; Hernandez-Barragan, A.; Flores-Vasconcelos, I.; Galicia-Moreno, M.; Rosique-Oramas, D.; Perez-Hernandez, J.L.; Higuera-De la Tijera, F.; Montalvo-Jave, E.E.; Torre-Delgadillo, A.; Cordero-Perez, P.; et al. Production and Activity of Matrix Metalloproteinases during Liver Fibrosis Progression of Chronic Hepatitis C Patients. World J. Hepatol. 2021, 13, 218–232. [Google Scholar] [CrossRef]
- De Caridi, G.; Bitto, A.; Massara, M.; Pallio, G.; Pizzino, G.; Serra, R.; Altavilla, D.; Squadrito, F.; Spinelli, F. Increased Serum HMGB-1, ICAM-1 and Metalloproteinase-9 Levels in Buerger’s Patients. Curr. Vasc. Pharmacol. 2016, 14, 382–387. [Google Scholar] [CrossRef]
- Wolk, K.; Brembach, T.-C.; Šimaitė, D.; Bartnik, E.; Cucinotta, S.; Pokrywka, A.; Irmer, M.L.; Triebus, J.; Witte-Händel, E.; Salinas, G.; et al. Activity and Components of the Granulocyte Colony-Stimulating Factor Pathway in Hidradenitis Suppurativa. Br. J. Dermatol. 2021, 185, 164–176. [Google Scholar] [CrossRef]
- Baima, J.P.; Imbrizi, M.; Andrade, A.R.; Chebli, L.A.; Argollo, M.C.; Queiroz, N.S.F.; de Azevedo, M.F.C.; Vieira, A.; Costa, M.H.d.M.; de Fróes, R.S.B.; et al. Second Brazilian Consensus on the Management of Ulcerative Colitis in Adults: A Consensus of the Brazilian Organization for Crohn’s Disease and Colitis (GEDIIB). Arq. Gastroenterol. 2023, 59, 51–84. [Google Scholar] [CrossRef]
- Mäkitalo, L.; Sipponen, T.; Kärkkäinen, P.; Kolho, K.-L.; Saarialho-Kere, U. Changes in Matrix Metalloproteinase (MMP) and Tissue Inhibitors of Metalloproteinases (TIMP) Expression Profile in Crohn’s Disease after Immunosuppressive Treatment Correlate with Histological Score and Calprotectin Values. Int. J. Color. Dis. 2009, 24, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, D.; Liszt, M.; Wanivenhaus, A.; Bröll, H.; Kiener, H.; Klösch, B.; Steiner, G. Hydrogen Sulphide Decreases IL-1β-Induced Activation of Fibroblast-like Synoviocytes from Patients with Osteoarthritis. J. Cell Mol. Med. 2015, 19, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.; Lee, W.; Park, J.; Choi, J.; Lee, S.; Li, S.; Jung, H.N.; Lee, J.-S.; Hwang, J.-E.; Park, J.; et al. Development of Spleen Targeting H2S Donor Loaded Liposome for the Effective Systemic Immunomodulation and Treatment of Inflammatory Bowel Disease. ACS Nano 2023, 17, 4327–4345. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, G.; Liu, X.; Song, J.; Chen, L.; Xu, H. Matrix Metalloproteinase-3 and the 7-Joint Ultrasound Score in the Assessment of Disease Activity and Therapeutic Efficacy in Patients with Moderate to Severe Rheumatoid Arthritis. Arthritis Res. Ther. 2017, 19, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowley, J.T.; Strle, K.; Drouin, E.E.; Pianta, A.; Arvikar, S.L.; Wang, Q.; Costello, C.E.; Steere, A.C. Matrix Metalloproteinase-10 Is a Target of T and B Cell Responses That Correlate with Synovial Pathology in Patients with Antibiotic-Refractory Lyme Arthritis. J. Autoimmun. 2016, 69, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, A.K.; Hildebrand, E.; Scagliotti, J.; Steere, A.C.; Hu, L.T. Induction of Host Matrix Metalloproteinases by Borrelia Burgdorferi Differs in Human and Murine Lyme Arthritis. Infect. Immun. 2005, 73, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, Y.; Li, Y.; Liao, C.; Dai, J.; Luo, Y.; Hu, Y.; Tao, S.; Tang, J.; Chen, G.; et al. Effect of Moxibustion on the Serum Levels of MMP-1, MMP-3, and VEGF in Patients with Rheumatoid Arthritis. Evid. Based. Complement. Alternat. Med. 2020, 2020, 7150605. [Google Scholar] [CrossRef]
- Zhou, J.-Y.; Luo, L.; Zhu, L.-L.; Yin, H.-Y.; Wu, Q.; Peng, J.-X.; Zhang, C.-S.; Lv, P.; Tang, Y.; Yu, S.-G. Moxibustion versus Diclofenac Sodium Gel for the Treatment of Knee Osteoarthritis: A Study Protocol for a Double-Blinded, Double-Placebo, Randomised Controlled Trial. BMJ Open 2017, 7, e012879. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tang, K.; Wang, Y.; Chen, Y.; Yang, M.; Gu, C.; Wang, J.; Wang, Y.; Yuan, Y. Elevated MicroRNA-145-5p Increases Matrix Metalloproteinase-9 by Activating the Nuclear Factor-κB Pathway in Rheumatoid Arthritis. Mol. Med. Rep. 2019, 20, 2703–2711. [Google Scholar] [CrossRef] [Green Version]
- Majster, M.; Lira-Junior, R.; Höög, C.M.; Almer, S.; Boström, E.A. Salivary and Serum Inflammatory Profiles Reflect Different Aspects of Inflammatory Bowel Disease Activity. Inflamm. Bowel Dis. 2020, 26, 1588–1596. [Google Scholar] [CrossRef]
- Soomro, S.; Venkateswaran, S.; Vanarsa, K.; Kharboutli, M.; Nidhi, M.; Susarla, R.; Zhang, T.; Sasidharan, P.; Lee, K.H.; Rosh, J.; et al. Predicting Disease Course in Ulcerative Colitis Using Stool Proteins Identified through an Aptamer-Based Screen. Nat. Commun. 2021, 12, 3989. [Google Scholar] [CrossRef]
- Buchbender, M.; Fehlhofer, J.; Proff, P.; Möst, T.; Ries, J.; Hannig, M.; Neurath, M.F.; Gund, M.; Atreya, R.; Kesting, M. Expression of Inflammatory Mediators in Biofilm Samples and Clinical Association in Inflammatory Bowel Disease Patients-a Preliminary Study. Clin. Oral Investig. 2022, 26, 1217–1228. [Google Scholar] [CrossRef]
- Lakatos, G.; Hritz, I.; Varga, M.Z.; Juhász, M.; Miheller, P.; Cierny, G.; Tulassay, Z.; Herszényi, L. The Impact of Matrix Metalloproteinases and Their Tissue Inhibitors in Inflammatory Bowel Diseases. Dig. Dis. 2012, 30, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Coufal, S.; Galanova, N.; Bajer, L.; Gajdarova, Z.; Schierova, D.; Jiraskova Zakostelska, Z.; Kostovcikova, K.; Jackova, Z.; Stehlikova, Z.; Drastich, P.; et al. Inflammatory Bowel Disease Types Differ in Markers of Inflammation, Gut Barrier and in Specific Anti-Bacterial Response. Cells 2019, 8, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boström, G.; Freyhult, E.; Virhammar, J.; Alcolea, D.; Tumani, H.; Otto, M.; Brundin, R.-M.; Kilander, L.; Löwenmark, M.; Giedraitis, V.; et al. Different Inflammatory Signatures in Alzheimer’s Disease and Frontotemporal Dementia Cerebrospinal Fluid. J. Alzheimers. Dis. 2021, 81, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chang, L.; Li, X.; Huang, J.; Yang, L.; Lai, X.; Huang, Z.; Wang, Z.; Wu, X.; Zhao, J.; et al. Tc17/IL-17A Up-Regulated the Expression of MMP-9 via NF-ΚB Pathway in Nasal Epithelial Cells of Patients with Chronic Rhinosinusitis. Front. Immunol. 2018, 9, 2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Lou, H.; Wang, X.; Wang, Y.; Fan, E.; Li, Y.; Wang, H.; Bachert, C.; Zhang, L. Effect of Budesonide Transnasal Nebulization in Patients with Eosinophilic Chronic Rhinosinusitis with Nasal Polyps. J. Allergy Clin. Immunol. 2015, 135, 922–929.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, D.; Sethi, G.; Ahn, K.S.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Aggarwal, A.; Aggarwal, B.B. Natural Products as a Gold Mine for Arthritis Treatment. Curr. Opin. Pharmacol. 2007, 7, 344–351. [Google Scholar] [CrossRef]
- Chen, W.-P.; Tang, J.-L.; Bao, J.-P.; Hu, P.-F.; Shi, Z.-L.; Wu, L.-D. Anti-Arthritic Effects of Chlorogenic Acid in Interleukin-1β-Induced Rabbit Chondrocytes and a Rabbit Osteoarthritis Model. Int. Immunopharmacol. 2011, 11, 23–28. [Google Scholar] [CrossRef]
- Huh, J.-E.; Seo, B.-K.; Baek, Y.-H.; Lee, S.; Lee, J.-D.; Choi, D.-Y.; Park, D.-S. Standardized Butanol Fraction of WIN-34B Suppresses Cartilage Destruction via Inhibited Production of Matrix Metalloproteinase and Inflammatory Mediator in Osteoarthritis Human Cartilage Explants Culture and Chondrocytes. BMC Complement. Altern. Med. 2012, 12, 256. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-C.; Zhang, Y.; Dai, B.-L.; Ma, Y.-J.; Zhang, Q.; Wang, Y.; Yang, H. Chlorogenic Acid Prevents Inflammatory Responses in IL-1β-stimulated Human SW-1353 Chondrocytes, a Model for Osteoarthritis. Mol. Med. Rep. 2017, 16, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.-J.; Wang, H.-D.; Shen, Z.-W.; Yao, X.-D.; Wu, C.-J.; Pan, T. Curcumin Inhibits Proliferation of Synovial Cells by Downregulating Expression of Matrix Metalloproteinase-3 in Osteoarthritis. Orthop. Surg. 2019, 11, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Liacini, A.; Sylvester, J.; Li, W.Q.; Zafarullah, M. Inhibition of Interleukin-1-Stimulated MAP Kinases, Activating Protein-1 (AP-1) and Nuclear Factor Kappa B (NF-Kappa B) Transcription Factors down-Regulates Matrix Metalloproteinase Gene Expression in Articular Chondrocytes. Matrix Biol. 2002, 21, 251–262. [Google Scholar] [CrossRef]
- Onodera, S.; Kaneda, K.; Mizue, Y.; Koyama, Y.; Fujinaga, M.; Nishihira, J. Macrophage Migration Inhibitory Factor Up-Regulates Expression of Matrix Metalloproteinases in Synovial Fibroblasts of Rheumatoid Arthritis. J. Biol. Chem. 2000, 275, 444–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Więcek, K.; Kupczyk, P.; Chodaczek, G.; Woźniak, M. The Impact of Curcumin on the Inflammatory Profile of SW982 Cells in a Rheumatoid Arthritis Model. J. Immunol. Res. 2022, 2022, 1208970. [Google Scholar] [CrossRef]
- Dai, Q.; Zhou, D.; Xu, L.; Song, X. Curcumin Alleviates Rheumatoid Arthritis-Induced Inflammation and Synovial Hyperplasia by Targeting MTOR Pathway in Rats. Drug Des. Devel. Ther. 2018, 12, 4095–4105. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, Z.; Anbazhagan, A.N.; Akhtar, N.; Ramamurthy, S.; Voss, F.R.; Haqqi, T.M. Green Tea Polyphenol Epigallocatechin-3-Gallate Inhibits Advanced Glycation End Product-Induced Expression of Tumor Necrosis Factor-Alpha and Matrix Metalloproteinase-13 in Human Chondrocytes. Arthritis Res. Ther. 2009, 11, R71. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Cao, G.; Ba, X.; Jiang, H. Epigallocatechin-3-O-Gallate Promotes Extracellular Matrix and Inhibits Inflammation in IL-1β Stimulated Chondrocytes by the PTEN/MiRNA-29b Pathway. Pharm. Biol. 2022, 60, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Leong, D.J.; Choudhury, M.; Hanstein, R.; Hirsh, D.M.; Kim, S.J.; Majeska, R.J.; Schaffler, M.B.; Hardin, J.A.; Spray, D.C.; Goldring, M.B.; et al. Green Tea Polyphenol Treatment Is Chondroprotective, Anti-Inflammatory and Palliative in a Mouse Post-Traumatic Osteoarthritis Model. Arthritis Res. Ther. 2014, 16, 508. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Liu, L.; Qu, Z.; Zhang, B.; Gao, X.; Huang, W.; Feng, M.; Gong, Y.; Kong, L.; Wang, Y.; et al. Hypoxia-Inducible Factor 1α Enhances RANKL-Induced Osteoclast Differentiation by Upregulating the MAPK Pathway. Ann. Transl. Med. 2022, 10, 1227. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Iwai, S.; Amano, H.; Irie, Y.; Yatomi, K.; Ryu, K.; Yamada, S.; Inagaki, K.; Oguchi, K. Tea Polyphenols Inhibit Rat Osteoclast Formation and Differentiation. J. Pharmacol. Sci. 2012, 118, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Yun, H.-J.; Yoo, W.-H.; Han, M.-K.; Lee, Y.-R.; Kim, J.-S.; Lee, S.-I. Epigallocatechin-3-Gallate Suppresses TNF-Alpha -Induced Production of MMP-1 and -3 in Rheumatoid Arthritis Synovial Fibroblasts. Rheumatol. Int. 2008, 29, 23–29. [Google Scholar] [CrossRef]
- Eastgate, J.A.; Symons, J.A.; Wood, N.C.; Grinlinton, F.M.; di Giovine, F.S.; Duff, G.W. Correlation of Plasma Interleukin 1 Levels with Disease Activity in Rheumatoid Arthritis. Lancet 1988, 2, 706–709. [Google Scholar] [CrossRef]
- Fechtner, S.; Singh, A.; Chourasia, M.; Ahmed, S. Molecular Insights into the Differences in Anti-Inflammatory Activities of Green Tea Catechins on IL-1β Signaling in Rheumatoid Arthritis Synovial Fibroblasts. Toxicol. Appl. Pharmacol. 2017, 329, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Pakozdi, A.; Koch, A.E. Regulation of Interleukin-1beta-Induced Chemokine Production and Matrix Metalloproteinase 2 Activation by Epigallocatechin-3-Gallate in Rheumatoid Arthritis Synovial Fibroblasts. Arthritis Rheum. 2006, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Marotte, H.; Kwan, K.; Ruth, J.H.; Campbell, P.L.; Rabquer, B.J.; Pakozdi, A.; Koch, A.E. Epigallocatechin-3-Gallate Inhibits IL-6 Synthesis and Suppresses Transsignaling by Enhancing Soluble Gp130 Production. Proc. Natl. Acad. Sci. USA 2008, 105, 14692–14697. [Google Scholar] [CrossRef]
- Poole, A.R.; Ionescu, M.; Fitzcharles, M.A.; Billinghurst, R.C. The Assessment of Cartilage Degradation in Vivo: Development of an Immunoassay for the Measurement in Body Fluids of Type II Collagen Cleaved by Collagenases. J. Immunol. Methods 2004, 294, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-C.; Wang, C.-C.; Lu, J.-W.; Lee, C.-H.; Chen, S.-C.; Ho, Y.-J.; Peng, Y.-J. Chondroprotective Effects of Genistein against Osteoarthritis Induced Joint Inflammation. Nutrients 2019, 11, 1180. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Dong, J.; He, P.; Li, W.; Zhang, Q.; Li, N.; Sun, T. Genistein Inhibit Cytokines or Growth Factor-Induced Proliferation and Transformation Phenotype in Fibroblast-like Synoviocytes of Rheumatoid Arthritis. Inflammation 2012, 35, 377–387. [Google Scholar] [CrossRef]
- Cheng, X.; Xu, P. The Articular Cartilage Preservative Effects of Genistein in an Experimental Model of Knees Osteoarthritis. Appl. Physiol. Nutr. Metab. 2021, 46, 1331–1336. [Google Scholar] [CrossRef]
- Hu, Y.; Gui, Z.; Zhou, Y.; Xia, L.; Lin, K.; Xu, Y. Quercetin Alleviates Rat Osteoarthritis by Inhibiting Inflammation and Apoptosis of Chondrocytes, Modulating Synovial Macrophages Polarization to M2 Macrophages. Free Radic. Biol. Med. 2019, 145, 146–160. [Google Scholar] [CrossRef]
- Wang, H.; Yan, Y.; Pathak, J.L.; Hong, W.; Zeng, J.; Qian, D.; Hao, B.; Li, H.; Gu, J.; Jaspers, R.T.; et al. Quercetin Prevents Osteoarthritis Progression Possibly via Regulation of Local and Systemic Inflammatory Cascades. J. Cell Mol. Med. 2023, 27, 515–528. [Google Scholar] [CrossRef]
- Sung, M.-S.; Lee, E.-G.; Jeon, H.-S.; Chae, H.-J.; Park, S.J.; Lee, Y.C.; Yoo, W.-H. Quercetin Inhibits IL-1β-Induced Proliferation and Production of MMPs, COX-2, and PGE2 by Rheumatoid Synovial Fibroblast. Inflammation 2012, 35, 1585–1594. [Google Scholar] [CrossRef]
- Li, W.; Hu, S.; Chen, X.; Shi, J. The Antioxidant Resveratrol Protects against Chondrocyte Apoptosis by Regulating the COX-2/NF-ΚB Pathway in Created Temporomandibular Osteoarthritis. Biomed Res. Int. 2021, 2021, 9978651. [Google Scholar] [CrossRef]
- Yi, H.; Zhang, W.; Cui, Z.-M.; Cui, S.-Y.; Fan, J.-B.; Zhu, X.-H.; Liu, W. Resveratrol Alleviates the Interleukin-1β-Induced Chondrocytes Injury through the NF-ΚB Signaling Pathway. J. Orthop. Surg. Res. 2020, 15, 424. [Google Scholar] [CrossRef] [PubMed]
- Moon, M.-H.; Jeong, J.-K.; Lee, Y.-J.; Seol, J.-W.; Jackson, C.J.; Park, S.-Y. SIRT1, a Class III Histone Deacetylase, Regulates TNF-α-Induced Inflammation in Human Chondrocytes. Osteoarthr. Cartil. 2013, 21, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elayyan, J.; Lee, E.; Gabay, O.; Smith, C.A.; Qiq, O.; Reich, E.; Mobasheri, A.; Henrotin, Y.; Kimber, S.J.; Dvir-Ginzberg, M. LEF1-mediated MMP13 Gene Expression Is Repressed by SIRT1 in Human Chondrocytes. FASEB J. 2017, 31, 3116–3125. [Google Scholar] [CrossRef] [Green Version]
- Wada, H.; Sato, Y.; Fujimoto, S.; Okamoto, K.; Bando, M.; Kawaguchi, T.; Miyamoto, H.; Muguruma, N.; Horimoto, K.; Matsuzawa, Y.; et al. Resveratrol Inhibits Development of Colorectal Adenoma via Suppression of LEF1; Comprehensive Analysis with Connectivity Map. Cancer Sci. 2022, 113, 4374–4384. [Google Scholar] [CrossRef] [PubMed]
- Dave, M.; Attur, M.; Palmer, G.; Al-Mussawir, H.E.; Kennish, L.; Patel, J.; Abramson, S.B. The Antioxidant Resveratrol Protects against Chondrocyte Apoptosis via Effects on Mitochondrial Polarization and ATP Production. Arthritis Rheum. 2008, 58, 2786–2797. [Google Scholar] [CrossRef]
- Hao, L.; Wan, Y.; Xiao, J.; Tang, Q.; Deng, H.; Chen, L. A Study of Sirt1 Regulation and the Effect of Resveratrol on Synoviocyte Invasion and Associated Joint Destruction in Rheumatoid Arthritis. Mol. Med. Rep. 2017, 16, 5099–5106. [Google Scholar] [CrossRef] [Green Version]
- Csaki, C.; Mobasheri, A.; Shakibaei, M. Synergistic Chondroprotective Effects of Curcumin and Resveratrol in Human Articular Chondrocytes: Inhibition of IL-1beta-Induced NF-KappaB-Mediated Inflammation and Apoptosis. Arthritis Res. Ther. 2009, 11, R165. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-C.; Hung, L.-F.; Wu, W.-L.; Chang, D.-M.; Huang, C.-Y.; Lai, J.-H.; Ho, L.-J. Chondroprotective Effects and Mechanisms of Resveratrol in Advanced Glycation End Products-Stimulated Chondrocytes. Arthritis Res. Ther. 2010, 12, R167. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, P.S.; Dash, D.; Singh, R. Intranasal Curcumin Inhibits Pulmonary Fibrosis by Modulating Matrix Metalloproteinase-9 (MMP-9) in Ovalbumin-Induced Chronic Asthma. Inflammation 2017, 40, 248–258. [Google Scholar] [CrossRef]
- Islam, R.; Dash, D.; Singh, R. Intranasal Curcumin and Sodium Butyrate Modulates Airway Inflammation and Fibrosis via HDAC Inhibition in Allergic Asthma. Cytokine 2022, 149, 155720. [Google Scholar] [CrossRef]
- Kumari, A.; Singh, D.K.; Dash, D.; Singh, R. Intranasal Curcumin Protects against LPS-Induced Airway Remodeling by Modulating Toll-like Receptor-4 (TLR-4) and Matrixmetalloproteinase-9 (MMP-9) Expression via Affecting MAP Kinases in Mouse Model. Inflammopharmacology 2019, 27, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Bae, C.H.; Song, S.-Y.; Kim, Y.-D. The Effect of Epigallocatechin-3-Gallate in Allergic Airway Inflammation. Rhinology 2014, 52, 406–412. [Google Scholar] [CrossRef]
- Kim, S.-H.; Park, H.-J.; Lee, C.-M.; Choi, I.-W.; Moon, D.-O.; Roh, H.-J.; Lee, H.-K.; Park, Y.-M. Epigallocatechin-3-Gallate Protects Toluene Diisocyanate-Induced Airway Inflammation in a Murine Model of Asthma. FEBS Lett. 2006, 580, 1883–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Palanivelu, K. The Effect of Curcumin (Turmeric) on Alzheimer’s Disease: An Overview. Ann. Indian Acad. Neurol. 2008, 11, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Heitman, E.; Ingram, D.K. Cognitive and Neuroprotective Effects of Chlorogenic Acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Syarifah-Noratiqah, S.-B.; Naina-Mohamed, I.; Zulfarina, M.S.; Qodriyah, H.M.S. Natural Polyphenols in the Treatment of Alzheimer’s Disease. Curr. Drug Targets 2018, 19, 927–937. [Google Scholar] [CrossRef]
- Fukutomi, R.; Ohishi, T.; Koyama, Y.; Pervin, M.; Nakamura, Y.; Isemura, M. Beneficial Effects of Epigallocatechin-3-O-Gallate, Chlorogenic Acid, Resveratrol, and Curcumin on Neurodegenerative Diseases. Molecules 2021, 26, 415. [Google Scholar] [CrossRef]
- Chong, Y.H.; Sung, J.H.; Shin, S.A.; Chung, J.H.; Suh, Y.H. Effects of the Beta-Amyloid and Carboxyl-Terminal Fragment of Alzheimer’s Amyloid Precursor Protein on the Production of the Tumor Necrosis Factor-Alpha and Matrix Metalloproteinase-9 by Human Monocytic THP-1. J. Biol. Chem. 2001, 276, 23511–23517. [Google Scholar] [CrossRef] [Green Version]
- Gamba, P.; Testa, G.; Sottero, B.; Gargiulo, S.; Poli, G.; Leonarduzzi, G. The Link between Altered Cholesterol Metabolism and Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 2012, 1259, 54–64. [Google Scholar] [CrossRef]
- Moussa, C.; Hebron, M.; Huang, X.; Ahn, J.; Rissman, R.A.; Aisen, P.S.; Turner, R.S. Resveratrol Regulates Neuro-Inflammation and Induces Adaptive Immunity in Alzheimer’s Disease. J. Neuroinflammation 2017, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.F.; Li, N.; Wang, Q.; Cheng, X.J.; Li, X.M.; Liu, T.T. Resveratrol Decreases the Insoluble Aβ1-42 Level in Hippocampus and Protects the Integrity of the Blood-Brain Barrier in AD Rats. Neuroscience 2015, 310, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.J.; Maut, C.; Gogoi, H.K.; Ahmed, S.I.; Kashyap, A. A Review on Herbal Drugs Used in the Treatment of Peptic Ulcer. Curr. Drug Discov. Technol. 2022, 20, e121222211869. [Google Scholar] [CrossRef]
- Swarnakar, S.; Ganguly, K.; Kundu, P.; Banerjee, A.; Maity, P.; Sharma, A. V Curcumin Regulates Expression and Activity of Matrix Metalloproteinases 9 and 2 during Prevention and Healing of Indomethacin-Induced Gastric Ulcer. J. Biol. Chem. 2005, 280, 9409–9415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motawi, T.K.; Rizk, S.M.; Shehata, A.H. Effects of Curcumin and Ginkgo Biloba on Matrix Metalloproteinases Gene Expression and Other Biomarkers of Inflammatory Bowel Disease. J. Physiol. Biochem. 2012, 68, 529–539. [Google Scholar] [CrossRef]
- Epstein, J.; Docena, G.; MacDonald, T.T.; Sanderson, I.R. Curcumin Suppresses P38 Mitogen-Activated Protein Kinase Activation, Reduces IL-1beta and Matrix Metalloproteinase-3 and Enhances IL-10 in the Mucosa of Children and Adults with Inflammatory Bowel Disease. Br. J. Nutr. 2010, 103, 824–832. [Google Scholar] [CrossRef] [Green Version]
- Fontani, F.; Marcucci, T.; Picariello, L.; Tonelli, F.; Vincenzini, M.T.; Iantomasi, T. Redox Regulation of MMP-3/TIMP-1 Ratio in Intestinal Myofibroblasts: Effect of N-Acetylcysteine and Curcumin. Exp. Cell Res. 2014, 323, 77–86. [Google Scholar] [CrossRef]
- Liu, Q.; Qian, Y.; Chen, F.; Chen, X.; Chen, Z.; Zheng, M. EGCG Attenuates Pro-Inflammatory Cytokines and Chemokines Production in LPS-Stimulated L02 Hepatocyte. Acta Biochim. Biophys. Sin. 2014, 46, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.-C.; Wu, Y.-H.; Huang, W.-C.; Pang, J.-H.S.; Huang, T.-H.; Cheng, C.-Y. Anti-Inflammatory Property of Quercetin through Downregulation of ICAM-1 and MMP-9 in TNF-α-Activated Retinal Pigment Epithelial Cells. Cytokine 2019, 116, 48–60. [Google Scholar] [CrossRef]
- Im, H.-J.; Li, X.; Chen, D.; Yan, D.; Kim, J.; Ellman, M.B.; Stein, G.S.; Cole, B.; Kc, R.; Cs-Szabo, G.; et al. Biological Effects of the Plant-Derived Polyphenol Resveratrol in Human Articular Cartilage and Chondrosarcoma Cells. J. Cell Physiol. 2012, 227, 3488–3497. [Google Scholar] [CrossRef] [Green Version]
- Cichocki, M.; Dałek, M.; Szamałek, M.; Baer-Dubowska, W. Naturally Occurring Phenolic Acids Modulate TPA-Induced Activation of EGFR, AP-1, and STATs in Mouse Epidermis. Nutr. Cancer 2014, 66, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Sarawi, W.S.; Alhusaini, A.M.; Fadda, L.M.; Alomar, H.A.; Albaker, A.B.; Aljrboa, A.S.; Alotaibi, A.M.; Hasan, I.H.; Mahmoud, A.M. Nano-Curcumin Prevents Cardiac Injury, Oxidative Stress and Inflammation, and Modulates TLR4/NF-ΚB and MAPK Signaling in Copper Sulfate-Intoxicated Rats. Antioxidants 2021, 10, 1414. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.-Y.; Wu, C.-C.; Hsu, H.-Y.; Chuang, H.-Y.; Huang, S.-Y.; Tsai, C.-H.; Chang, Y.; Tsao, G.S.-W.; Chen, C.-L.; Chen, J.-Y. EGCG Inhibits Proliferation, Invasiveness and Tumor Growth by up-Regulation of Adhesion Molecules, Suppression of Gelatinases Activity, and Induction of Apoptosis in Nasopharyngeal Carcinoma Cells. Int. J. Mol. Sci. 2015, 16, 2530–2558. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-D.; Chen, B.-C.; Kao, S.-T.; Liu, C.-J.; Yeh, C.-C. Genistein Inhibits Tumor Invasion by Suppressing Multiple Signal Transduction Pathways in Human Hepatocellular Carcinoma Cells. BMC Complement. Altern. Med. 2014, 14, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, M.K.; Song, N.R.; Kang, N.J.; Lee, K.W.; Lee, H.J. Activation of Phosphatidylinositol 3-Kinase Is Required for Tumor Necrosis Factor-Alpha-Induced Upregulation of Matrix Metalloproteinase-9: Its Direct Inhibition by Quercetin. Int. J. Biochem. Cell Biol. 2009, 41, 1592–1600. [Google Scholar] [CrossRef]
- Kim, Y.S.; Sull, J.W.; Sung, H.J. Suppressing Effect of Resveratrol on the Migration and Invasion of Human Metastatic Lung and Cervical Cancer Cells. Mol. Biol. Rep. 2012, 39, 8709–8716. [Google Scholar] [CrossRef]
- Hu, X.; Wang, L.; He, Y.; Wei, M.; Yan, H.; Zhu, H. Chlorogenic Acid Promotes Osteogenic Differentiation of Human Dental Pulp Stem Cells Through Wnt Signaling. Stem Cells Dev. 2021, 30, 641–650. [Google Scholar] [CrossRef]
- Mohamadian, M.; Bahrami, A.; Moradi Binabaj, M.; Asgharzadeh, F.; Ferns, G.A. Molecular Targets of Curcumin and Its Therapeutic Potential for Ovarian Cancer. Nutr. Cancer 2022, 74, 2713–2730. [Google Scholar] [CrossRef]
- Yeh, C.-C.; Fan, Y.; Jiang, L.; Yang, Y.-L.; He, B.; You, L.; Mann, M. Genistein Suppresses Growth of Human Uterine Sarcoma Cell Lines via Multiple Mechanisms. Anticancer Res. 2015, 35, 3167–3173. [Google Scholar]
- Murata, M.; Komatsu, S.; Miyamoto, E.; Oka, C.; Lin, I.; Kumazoe, M.; Yamashita, S.; Fujimura, Y.; Tachibana, H. Quercetin Up-Regulates the Expression of Tumor-Suppressive MicroRNAs in Human Cervical Cancer. Biosci. Microbiota Food Health 2023, 42, 87–93. [Google Scholar] [CrossRef]
- Xie, C.; Liang, C.; Wang, R.; Yi, K.; Zhou, X.; Li, X.; Chen, Y.; Miao, D.; Zhong, C.; Zhu, J. Resveratrol Suppresses Lung Cancer by Targeting Cancer Stem-like Cells and Regulating Tumor Microenvironment. J. Nutr. Biochem. 2023, 112, 109211. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Wang, C.; Yu, L.; Sheng, T.; Wu, Z.; Wang, X.; Zhang, D.; Lin, Y.; Gong, Y. Chlorogenic Acid Attenuates Dextran Sodium Sulfate-Induced Ulcerative Colitis in Mice through MAPK/ERK/JNK Pathway. Biomed Res. Int. 2019, 2019, 6769789. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Ip, M.S.M.; Mak, J.C.W. (-)-Epigallocatechin-3-Gallate Suppresses Cigarette Smoke-Induced Inflammation in Human Cardiomyocytes via ROS-Mediated MAPK and NF-ΚB Pathways. Phytomedicine 2019, 58, 152768. [Google Scholar] [CrossRef]
- Ye, G.; Lin, C.; Zhang, Y.; Ma, Z.; Chen, Y.; Kong, L.; Yuan, L.; Ma, T. Quercetin Alleviates Neuropathic Pain in the Rat CCI Model by Mediating AMPK/MAPK Pathway. J. Pain Res. 2021, 14, 1289–1301. [Google Scholar] [CrossRef]
- Chang, W.-S.; Tsai, C.-W.; Yang, J.-S.; Hsu, Y.-M.; Shih, L.-C.; Chiu, H.-Y.; Bau, D.-T.; Tsai, F.-J. Resveratrol Inhibited the Metastatic Behaviors of Cisplatin-Resistant Human Oral Cancer Cells via Phosphorylation of ERK/p-38 and Suppression of MMP-2/9. J. Food Biochem. 2021, 45, e13666. [Google Scholar] [CrossRef] [PubMed]
- Tikoo, K.; Meena, R.L.; Kabra, D.G.; Gaikwad, A.B. Change in Post-Translational Modifications of Histone H3, Heat-Shock Protein-27 and MAP Kinase P38 Expression by Curcumin in Streptozotocin-Induced Type I Diabetic Nephropathy. Br. J. Pharmacol. 2008, 153, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, J.; Tian, C.; Zeng, Y.; Zheng, Y.; Fang, Q.; Li, H. Epigallocatechin Gallate Inhibits Angiotensin II-Induced Endothelial Barrier Dysfunction via Inhibition of the P38 MAPK/HSP27 Pathway. Acta Pharmacol. Sin. 2010, 31, 1401–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Bergan, R.C. Genistein Inhibits Matrix Metalloproteinase Type 2 Activation and Prostate Cancer Cell Invasion by Blocking the Transforming Growth Factor Beta-Mediated Activation of Mitogen-Activated Protein Kinase-Activated Protein Kinase 2-27-KDa Heat Shock Protein Pa. Mol. Pharmacol. 2006, 70, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Sang, D.-P.; Li, R.-J.; Lan, Q. Quercetin Sensitizes Human Glioblastoma Cells to Temozolomide in Vitro via Inhibition of Hsp27. Acta Pharmacol. Sin. 2014, 35, 832–838. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Chávez, J.; Fonseca-Sánchez, M.A.; Arechaga-Ocampo, E.; Flores-Pérez, A.; Palacios-Rodríguez, Y.; Domínguez-Gómez, G.; Marchat, L.A.; Fuentes-Mera, L.; Mendoza-Hernández, G.; Gariglio, P.; et al. Proteomic Profiling Reveals That Resveratrol Inhibits HSP27 Expression and Sensitizes Breast Cancer Cells to Doxorubicin Therapy. PLoS ONE 2013, 8, e64378. [Google Scholar] [CrossRef] [Green Version]
- Moslehi, A.; Komeili-Movahhed, T.; Ahmadian, M.; Ghoddoosi, M.; Heidari, F. Chlorogenic Acid Attenuates Liver Apoptosis and Inflammation in Endoplasmic Reticulum Stress-Induced Mice. Iran. J. Basic Med. Sci. 2023, 26, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Daglia, M.; Tundis, R.; Loizzo, M.R.; Sobarzo-Sanchez, E.; Orhan, I.E.; Nabavi, S.M. Genistein: A Boon for Mitigating Ischemic Stroke. Curr. Top. Med. Chem. 2015, 15, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-Y.; Park, S.-J.; Kwon, M.-J.; Jeong, T.-S.; Bok, S.-H.; Choi, W.-Y.; Jeong, W.-I.; Ryu, S.-Y.; Do, S.-H.; Lee, C.-S.; et al. Quercetin Suppresses Proinflammatory Cytokines Production through MAP Kinases AndNF-KappaB Pathway in Lipopolysaccharide-Stimulated Macrophage. Mol. Cell Biochem. 2003, 243, 153–160. [Google Scholar] [CrossRef]
- Liu, T.; Long, T.; Li, H. Curcumin Suppresses the Proliferation of Oral Squamous Cell Carcinoma through a Specificity Protein 1/Nuclear Factor-ΚB-Dependent Pathway. Exp. Ther. Med. 2021, 21, 202. [Google Scholar] [CrossRef]
- Miyamoto, N.; Higuchi, Y.; Tsurudome, M.; Ito, M.; Nishio, M.; Kawano, M.; Sudo, A.; Kato, K.; Uchida, A.; Ito, Y. Induction of C-Src in Human Blood Monocytes by Anti-CD98/FRP-1 MAb in an Sp1-Dependent Fashion. Cell Immunol. 2000, 204, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Cho, J.H.; Jeon, Y.-J.; Bang, W.; Cho, J.-J.; Choi, N.-J.; Seo, K.S.; Shim, J.-H.; Chae, J.-I. Quercetin Induces Antiproliferative Activity Against Human Hepatocellular Carcinoma (HepG2) Cells by Suppressing Specificity Protein 1 (Sp1). Drug Dev. Res. 2015, 76, 9–16. [Google Scholar] [CrossRef]
- Zeng, K.; Wang, Y.; Yang, N.; Wang, D.; Li, S.; Ming, J.; Wang, J.; Yu, X.; Song, Y.; Zhou, X.; et al. Resveratrol Inhibits Diabetic-Induced Müller Cells Apoptosis through MicroRNA-29b/Specificity Protein 1 Pathway. Mol. Neurobiol. 2017, 54, 4000–4014. [Google Scholar] [CrossRef]
- Tan, S.; Yan, F.; Li, Q.; Liang, Y.; Yu, J.; Li, Z.; He, F.; Li, R.; Li, M. Chlorogenic Acid Promotes Autophagy and Alleviates Salmonella Typhimurium Infection Through the LncRNAGAS5/MiR-23a/PTEN Axis and the P38 MAPK Pathway. Front. Cell Dev. Biol. 2020, 8, 552020. [Google Scholar] [CrossRef]
- Sazuka, M.; Imazawa, H.; Shoji, Y.; Mita, T.; Hara, Y.; Isemura, M. Inhibition of Collagenases from Mouse Lung Carcinoma Cells by Green Tea Catechins and Black Tea Theaflavins. Biosci. Biotechnol. Biochem. 1997, 61, 1504–1506. [Google Scholar] [CrossRef] [Green Version]
- Djerir, D.; Iddir, M.; Bourgault, S.; Lamy, S.; Annabi, B. Biophysical Evidence for Differential Gallated Green Tea Catechins Binding to Membrane Type-1 Matrix Metalloproteinase and Its Interactors. Biophys. Chem. 2018, 234, 34–41. [Google Scholar] [CrossRef]
- Tsuchiya, A.; Kobayashi, M.; Kamatari, Y.O.; Mitsunaga, T.; Yamauchi, K. Development of Flavonoid Probes and the Binding Mode of the Target Protein and Quercetin Derivatives. Bioorg. Med. Chem. 2022, 68, 116854. [Google Scholar] [CrossRef]
- Shunmuga Priya, V.; Pradiba, D.; Aarthy, M.; Singh, S.K.; Achary, A.; Vasanthi, M. In-Silico Strategies for Identification of Potent Inhibitor for MMP-1 to Prevent Metastasis of Breast Cancer. J. Biomol. Struct. Dyn. 2021, 39, 7274–7293. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, R.; Cai, X.; Huang, X.; Ye, Z. Mechanism of Action of Ermiao San on Rheumatoid Arthritis Based on Bioinformatics and Molecular Dynamics. Comb. Chem. High Throughput Screen. 2022, 25, 2153–2164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, B.; Huang, J.; Huang, S.; He, D.; Peng, W.; Zhang, S. A Network Pharmacology Analysis of the Active Components of the Traditional Chinese Medicine Zuojinwan in Patients with Gastric Cancer. Med. Sci. Monit. 2020, 26, e923327. [Google Scholar] [CrossRef] [PubMed]
- D’Abadia, P.L.; BailÃo, E.F.L.C.; Lino JÚnior, R.S.; Oliveira, M.G.; Silva, V.B.; Oliveira, L.A.R.; ConceiÇÃo, E.C.; Melo-Reis, P.R.; Borges, L.L.; GonÇalves, P.J.; et al. Hancornia Speciosa Serum Fraction Latex Stimulates the Angiogenesis and Extracellular Matrix Remodeling Processes. An. Acad. Bras. Cienc. 2020, 92, e20190107. [Google Scholar] [CrossRef]
- Seyed Hosseini, E.; Alizadeh Zarei, M.; Tarrahimofrad, H.; Zamani, J.; Haddad Kashani, H.; Ahmad, E.; Nikzad, H. Synergistic Effects of Dendrosomal Nanocurcumin and Oxaliplatin on Oncogenic Properties of Ovarian Cancer Cell Lines by Down-Expression of MMPs. Biol. Res. 2023, 56, 3. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi Mirgani, M.; Isacchi, B.; Sadeghizadeh, M.; Marra, F.; Bilia, A.R.; Mowla, S.J.; Najafi, F.; Babaei, E. Dendrosomal Curcumin Nanoformulation Downregulates Pluripotency Genes via MiR-145 Activation in U87MG Glioblastoma Cells. Int. J. Nanomedicine 2014, 9, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Sayed, A.; Ginnebaugh, K.R.; Sharma, V.; Suri, A.; Saraph, A.; Padhye, S.; Sarkar, F.H. Molecular Docking and Inhibition of Matrix Metalloproteinase-2 by Novel Difluorinatedbenzylidene Curcumin Analog. Am. J. Transl. Res. 2015, 7, 298–308. [Google Scholar]
- Chowdhury, A.; Nandy, S.K.; Sarkar, J.; Chakraborti, T.; Chakraborti, S. Inhibition of Pro-/Active MMP-2 by Green Tea Catechins and Prediction of Their Interaction by Molecular Docking Studies. Mol. Cell Biochem. 2017, 427, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Shi, Y.; Zhang, Y.; Yang, X.; Wang, Y.; Jiang, K.; Hua, C.; Wu, C.; Sun, C.; Qin, Y.; et al. Investigating the Mechanism of Xian-Ling-Lian-Xia-Fang for Inhibiting Vasculogenic Mimicry in Triple Negative Breast Cancer via Blocking VEGF/MMPs Pathway. Chin. Med. 2022, 17, 44. [Google Scholar] [CrossRef]
- Erusappan, T.; Kondapuram, S.K.; Ekambaram, S.P.; Coumar, M.S. Investigation of Alpinia Calcarata Constituent Interactions with Molecular Targets of Rheumatoid Arthritis: Docking, Molecular Dynamics, and Network Approach. J. Mol. Model. 2021, 27, 14. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kang, F.; Wang, W.; Liu, S.; Xie, J.; Yang, X. Comparison between Heat-Clearing Medicine and Antirheumatic Medicine in Treatment of Gastric Cancer Based on Network Pharmacology, Molecular Docking, and Tumor Immune Infiltration Analysis. Evid. Based. Complement. Alternat. Med. 2022, 2022, 7490279. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Verma, S.; Bhattacharya, P.; Paul, S.; Mishra, A.; Patnaik, R. An In-Silico Strategy to Explore Neuroprotection by Quercetin in Cerebral Ischemia: A Novel Hypothesis Based on Inhibition of Matrix Metalloproteinase (MMPs) and Acid Sensing Ion Channel 1a (ASIC1a). Med. Hypotheses 2012, 79, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Jerah, A.; Hobani, Y.; Kumar, B.V.; Bidwai, A. Curcumin Binds in Silico to Anti-Cancer Drug Target Enzyme MMP-3 (Human Stromelysin-1) with Affinity Comparable to Two Known Inhibitors of the Enzyme. Bioinformation 2015, 11, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Wees, M.A.; Faure, T.D.; Carrillo, C.; Arbiser, J.; Bowen, J.P. The Impact of Ionization States of Matrix Metalloproteinase Inhibitors on Docking-Based Inhibitor Design. ACS Med. Chem. Lett. 2011, 2, 455–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, J.; Nandy, S.K.; Chowdhury, A.; Chakraborti, T.; Chakraborti, S. Inhibition of MMP-9 by Green Tea Catechins and Prediction of Their Interaction by Molecular Docking Analysis. Biomed. Pharmacother. 2016, 84, 340–347. [Google Scholar] [CrossRef]
- Yu, J.; Fu, Y.; Zeng, L.; Zheng, Y. Investigating the Active Components of Huatan Tongjing Decoction for the Treatment of Polycystic Ovary Syndrome via Network Pharmacology. Chem. Biol. Drug Des. 2022, 100, 515–524. [Google Scholar] [CrossRef]
- Saragusti, A.C.; Ortega, M.G.; Cabrera, J.L.; Estrin, D.A.; Marti, M.A.; Chiabrando, G.A. Inhibitory Effect of Quercetin on Matrix Metalloproteinase 9 Activity Molecular Mechanism and Structure-Activity Relationship of the Flavonoid-Enzyme Interaction. Eur. J. Pharmacol. 2010, 644, 138–145. [Google Scholar] [CrossRef]
- Huynh, C.B.; Nagaarudkumaran, N.; Kalyaanamoorthy, S.; Ngo, W. In Silico and In Vitro Approach for Validating the Inhibition of Matrix Metalloproteinase-9 by Quercetin. Eye Contact Lens 2023, 49, 193–198. [Google Scholar] [CrossRef]
- Vicentini, F.T.M.C.; He, T.; Shao, Y.; Fonseca, M.J.V.; Verri, W.A.; Fisher, G.J.; Xu, Y. Quercetin Inhibits UV Irradiation-Induced Inflammatory Cytokine Production in Primary Human Keratinocytes by Suppressing NF-ΚB Pathway. J. Dermatol. Sci. 2011, 61, 162–168. [Google Scholar] [CrossRef]
- Ohishi, T.; Hayakawa, S.; Miyoshi, N. Involvement of MicroRNA Modifications in Anticancer Effects of Major Polyphenols from Green Tea, Coffee, Wine, and Curry. Crit. Rev. Food Sci. Nutr. 2022, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, E.; Rajasekaran, R. Quantum Chemical and Molecular Mechanics Studies on the Assessment of Interactions between Resveratrol and Mutant SOD1 (G93A) Protein. J. Comput. Aided. Mol. Des. 2018, 32, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Baruah, I.; Kashyap, C.; Guha, A.K.; Borgohain, G. Insights into the Interaction between Polyphenols and β-Lactoglobulin through Molecular Docking, MD Simulation, and QM/MM Approaches. ACS Omega 2022, 7, 23083–23095. [Google Scholar] [CrossRef] [PubMed]

















| CGA | CUR | EGCG | GEN | QUR | RES | |
|---|---|---|---|---|---|---|
| AP1 | Cichocki et al. [101] | Sarawi et al. [102] | Fang et al. [103] | Wang et al. [104] | Hwang et al. [105] | Kim et al. [106] |
| β-catenin | Hu et al. [107] | Mohamadian et al. [108] | Fang et al. [103] | Yeh et al. [109] | Murata et al. [110] | Xie et al. [111] |
| ERK1/2 | Gao et al. [112] | Mohamadian et al. [108] | Liang et al. [113] | Yeh et al. [109] | Ye et al. [114] | Chang et al. [115] |
| Hsp27 | NF * | Tikoo et al. [116] | Yang et al. [117] | Xu et al. [118] | Sang et al. [119] | Díaz-Chávez et al. [120] |
| NF-κB | Moslehi et al. [121] | Sarawi et al. [102] | Fang et al. [103] | Nabavi et al. [122] | Cho et al. [123] | Kim et al. [106] |
| Specificity protein 1 (Sp1) | NF * | Liu et al. [124] | Fang et al. [103] | Miyamoto et al. [125] | Lee et al. [126] | Zeng et al. [127] |
| p38 | Tan et al. [128] | Tikoo et al. [116] | Fechtner et al. [61] | Xu et al. [118] | Ye et al. [114] | Chang et al. [115] |
| QUE | CUR | EGCG | |
|---|---|---|---|
| MMP1 | −7.15 kcal/mol (1) (Glu219) | ||
| MMP2 | −8.17 kcal/mol (2) (Leu82, Ala139, Ile141) −10.1 kcal/mol (3) (Leu83, Val117, Glu121, Ala136) −7.90 kcal/mol (4) (Tyr3, Ile141, Thr143, Thr145, Asn147, Phe148) −9.11 kcal/mol (5) (Leu164, Ala165, Ala217, Ala220) | −7.35 kcal/mol (8) (Arg149) | −32.72 kcal/mol (10) (Leu399, His403, Glu404, Ala192) |
| MMP3 | −7.25 kcal/mol (1) (Tyr155, Leu164) | −10.2 kcal/mol (9) (Leu164, Ala165, Leu218) | |
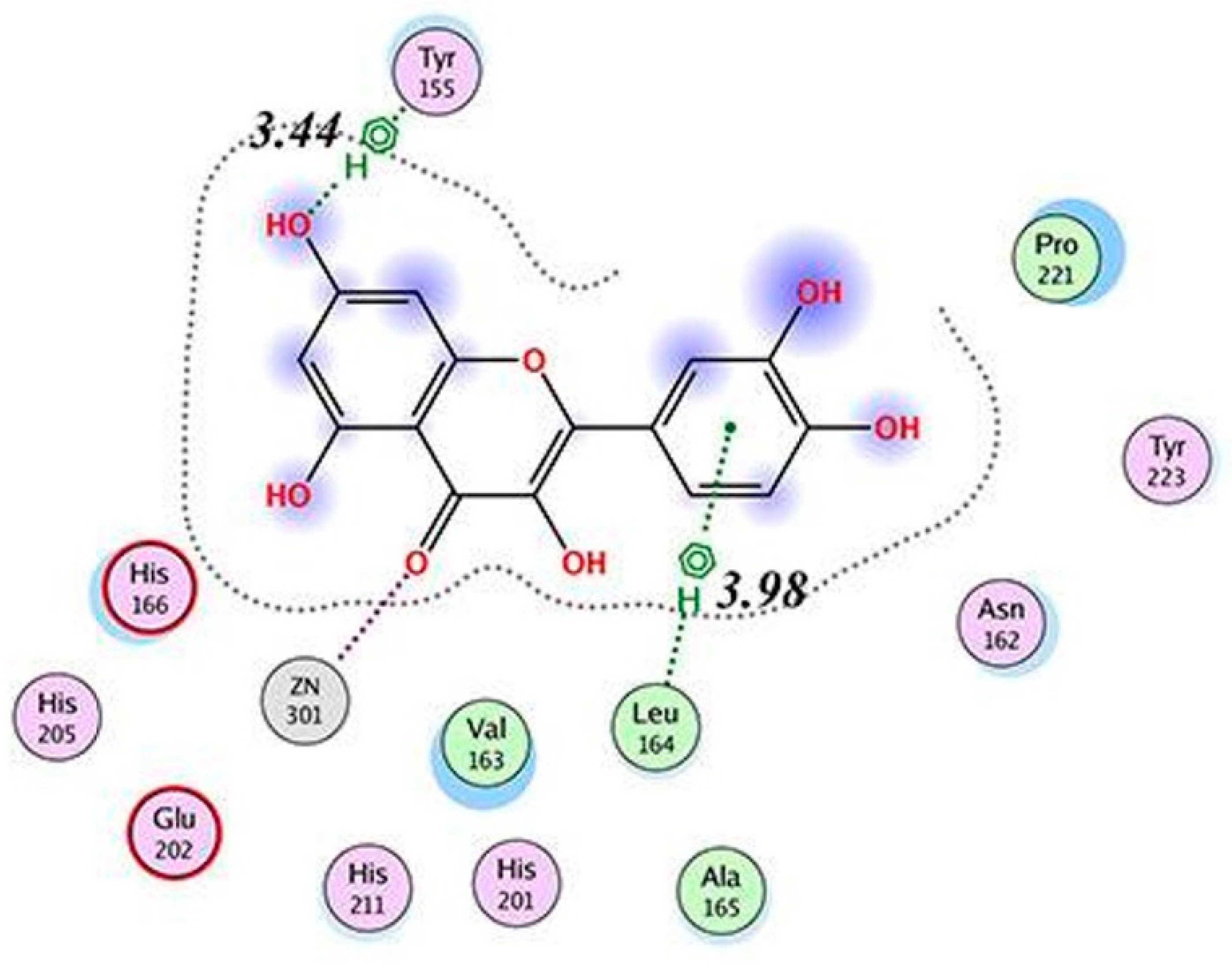
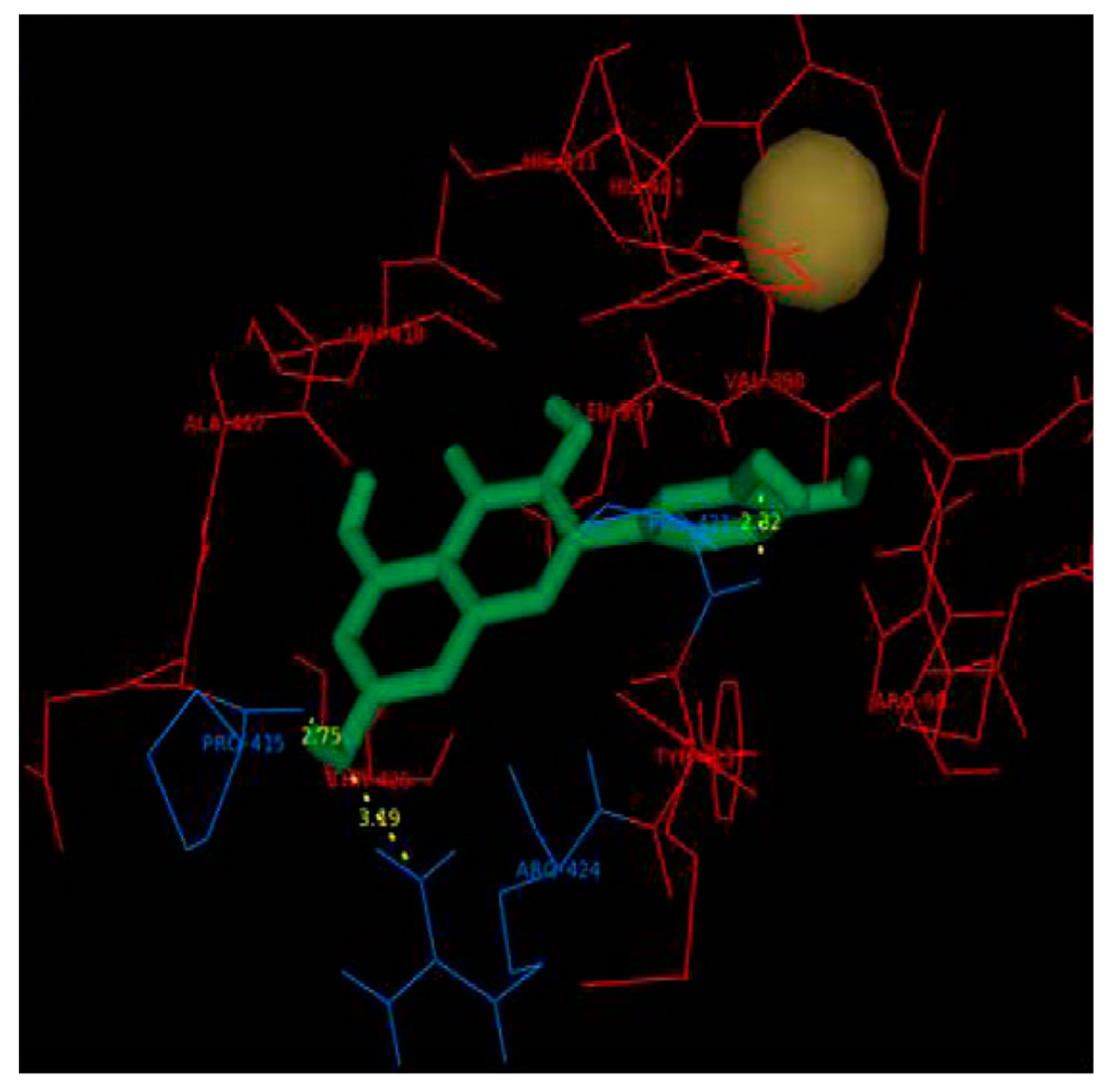
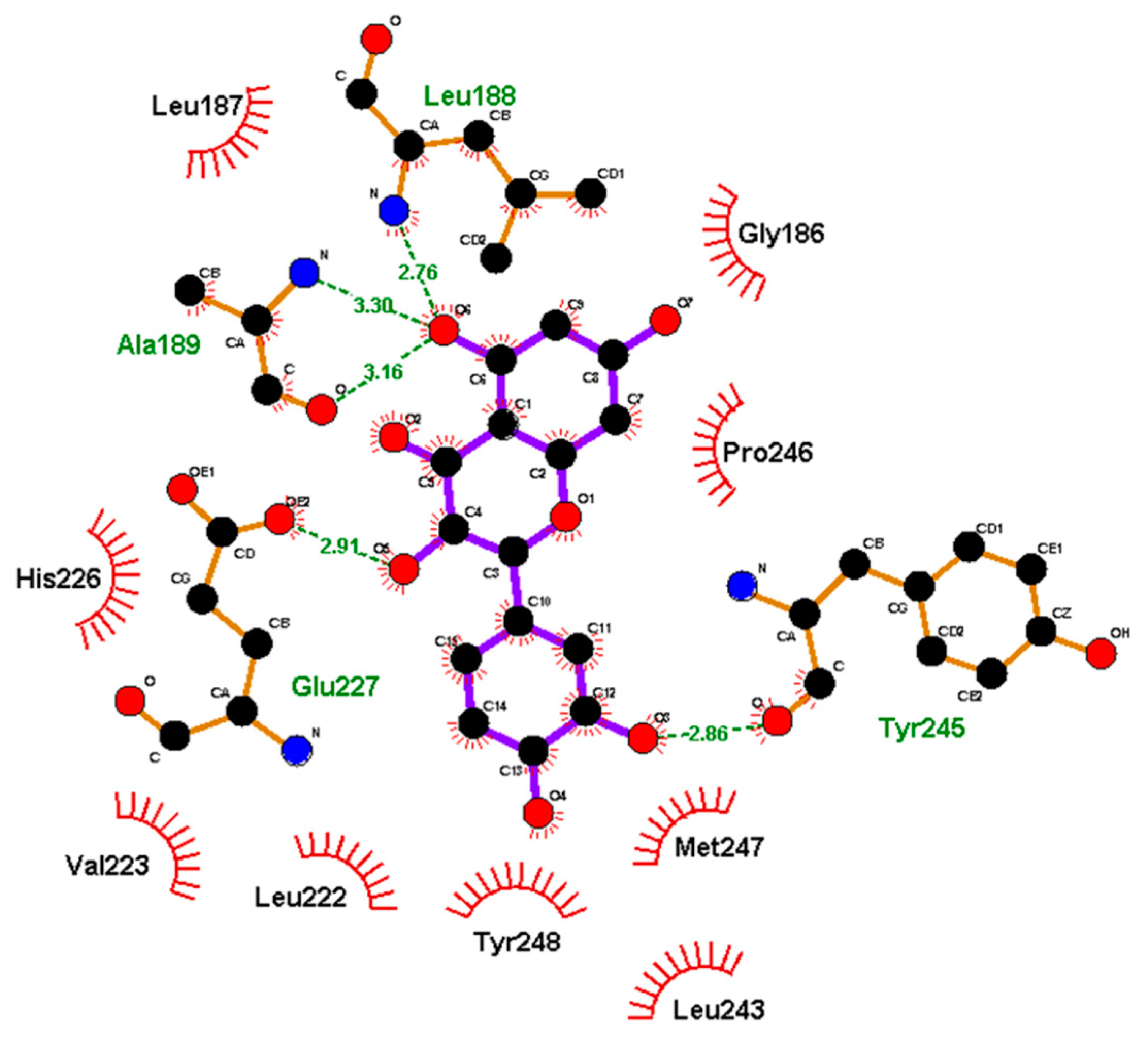
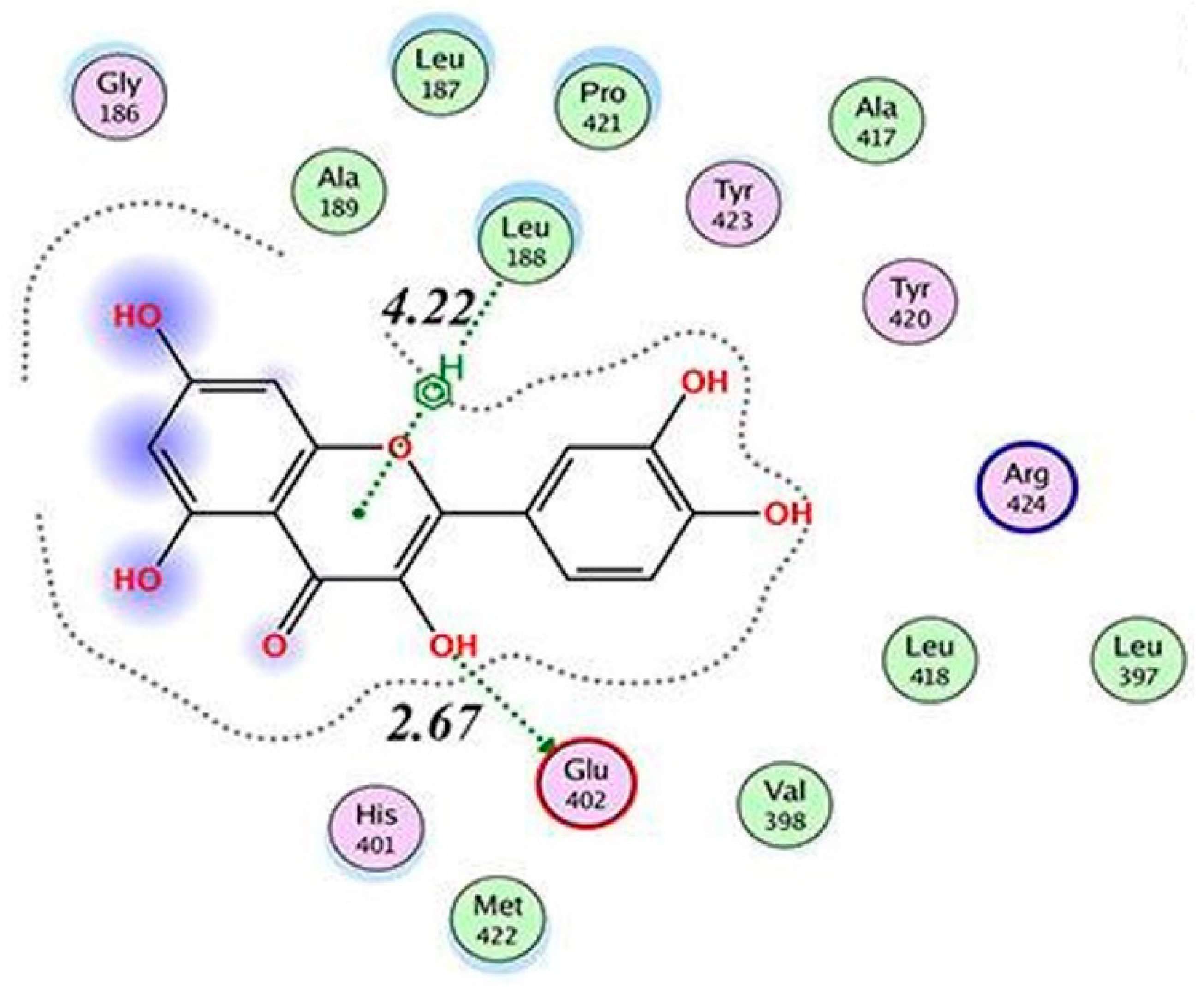
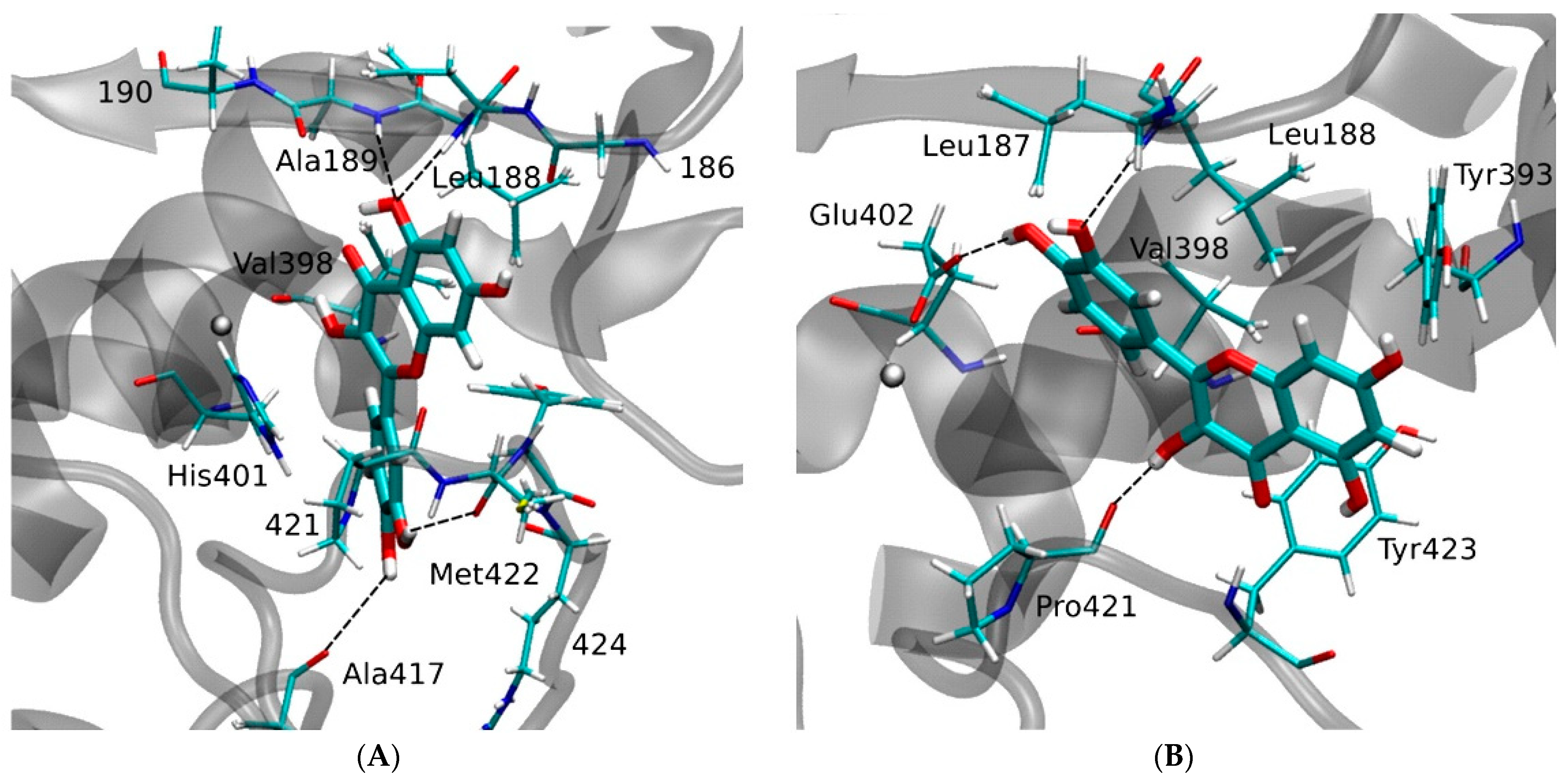
| MMP9 | −8.82 kcal/mol (5) (Pro415)−10.8 kcal/mol (6) (Leu188, Ala189, Glu227, Tyr245) −6.16 kcal/mol (1) (Glu402)−9.9 kcal/mol (7) (Leu188, Ala189, Glu227, Met247) | ||
| MMP14 | −57.61 kcal/mol (10) (Leu199, Phe234, His239, Glu240, Met257, Gln262) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Ohishi, T.; Tanabe, H.; Miyoshi, N.; Nakamura, Y. Anti-Inflammatory Effects of Dietary Polyphenols through Inhibitory Activity against Metalloproteinases. Molecules 2023, 28, 5426. https://doi.org/10.3390/molecules28145426
Suzuki T, Ohishi T, Tanabe H, Miyoshi N, Nakamura Y. Anti-Inflammatory Effects of Dietary Polyphenols through Inhibitory Activity against Metalloproteinases. Molecules. 2023; 28(14):5426. https://doi.org/10.3390/molecules28145426
Chicago/Turabian StyleSuzuki, Takuji, Tomokazu Ohishi, Hiroki Tanabe, Noriyuki Miyoshi, and Yoriyuki Nakamura. 2023. "Anti-Inflammatory Effects of Dietary Polyphenols through Inhibitory Activity against Metalloproteinases" Molecules 28, no. 14: 5426. https://doi.org/10.3390/molecules28145426
APA StyleSuzuki, T., Ohishi, T., Tanabe, H., Miyoshi, N., & Nakamura, Y. (2023). Anti-Inflammatory Effects of Dietary Polyphenols through Inhibitory Activity against Metalloproteinases. Molecules, 28(14), 5426. https://doi.org/10.3390/molecules28145426