
Effects of Different Freezing Rate and Frozen Storage Temperature on the Quality of Large-Mouth Bass (Micropterus salmoides)

Abstract
:1. Introduction
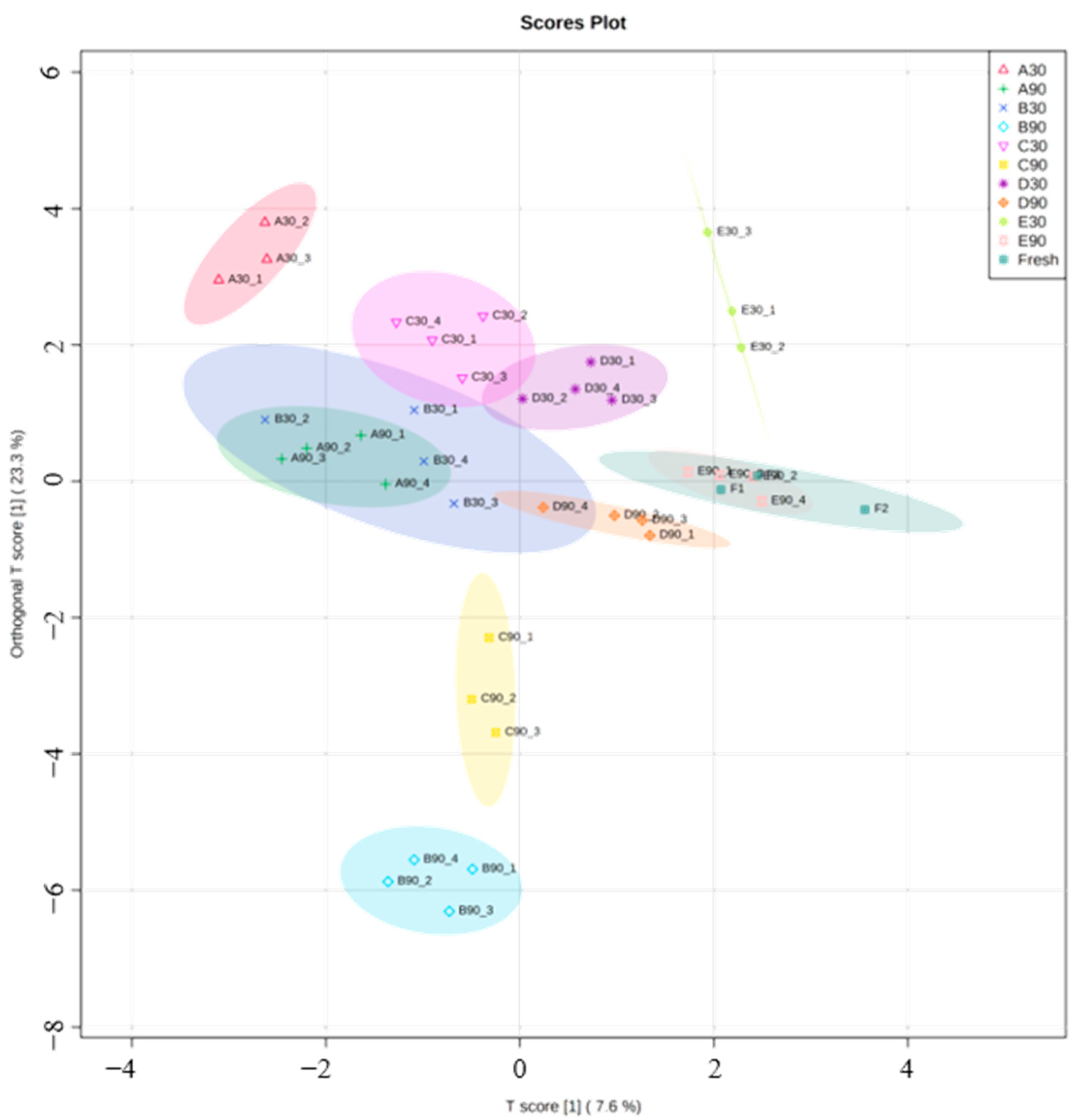
2. Results and Discussion
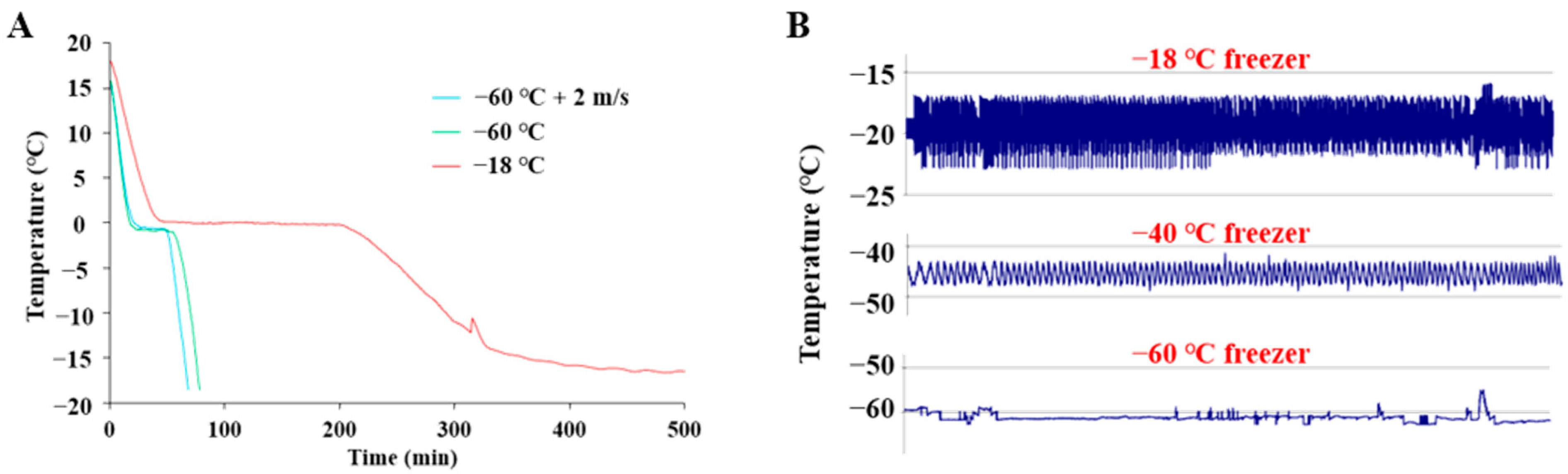
2.1. The Freezing Process and Frozen Storage
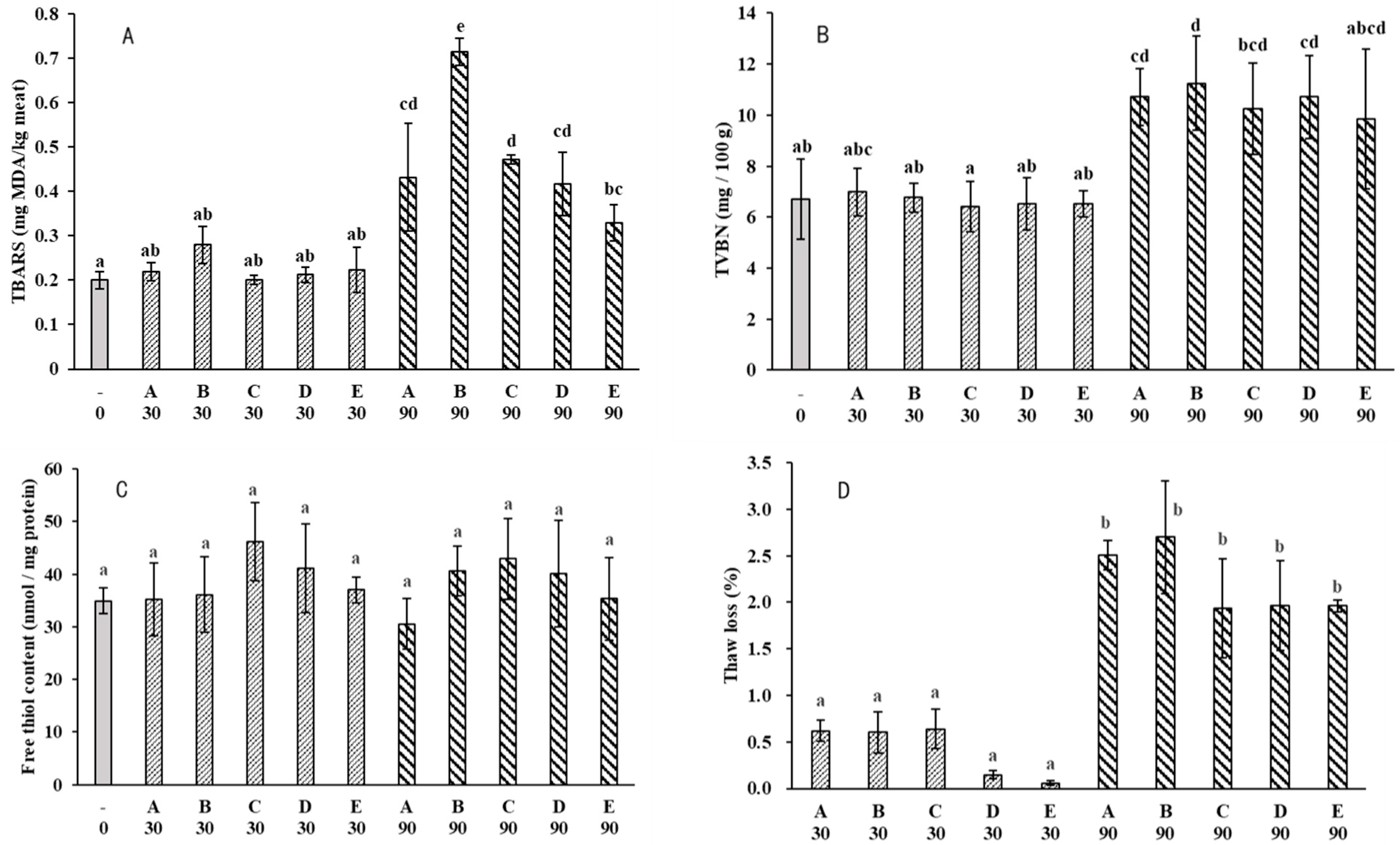
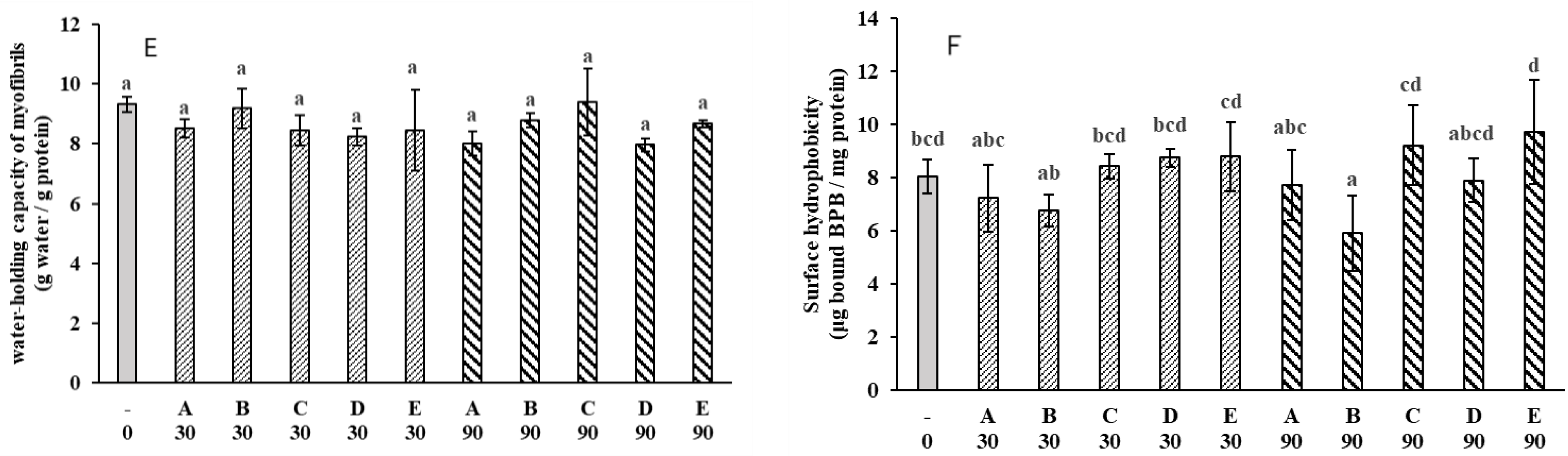
2.2. Water-Holding
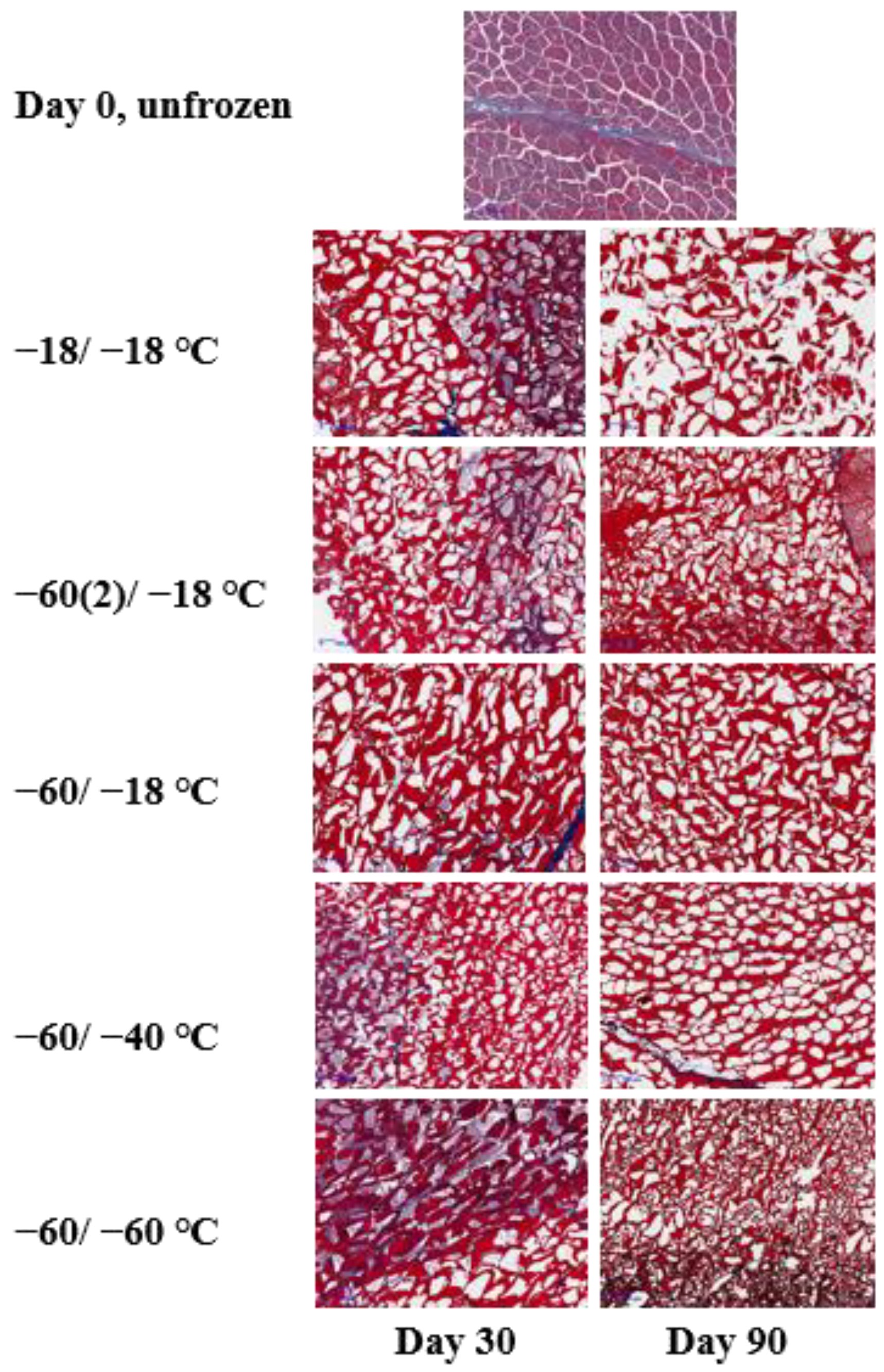
2.3. Microstructure Changes
2.4. Lipid Oxidation
2.5. Protein Changes
3. Materials and Methods
3.1. Sample Preparation
3.2. Water-Holding Capacity
3.3. Lipid Oxidation
3.4. Total Volatile Basic Nitrogen (TVB-N)
3.5. Protein Oxidation and Denaturation
3.6. Freeze Substitution Histological Observation
3.7. Comprehensive Two-Dimensional Gas Chromatography Time-of-Flight Mass Spectrometry (GC × GC-TOFMS)
3.8. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fishery Bureau of Ministry of Agriculture of the People’s Republic of China. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2021.
- Bao, Y.; Ertbjerg, P.; Estévez, M.; Yuan, L.; Gao, R. Freezing of meat and aquatic food: Underlying mechanisms and implications on protein oxidation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5548–5569. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qian, S.; Song, Y.; Guo, Y.; Huang, F.; Han, D.; Zhang, C.; Blecker, C. New insights into the mechanism of freeze-induced damage based on ice crystal morphology and exudate proteomics. Food Res. Int. 2022, 161, 111757. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ertbjerg, P. On the origin of thaw loss: Relationship between freezing rate and protein denaturation. Food Chem. 2019, 299, 125104. [Google Scholar] [CrossRef]
- Nakazawa, N.; Okazaki, E. Recent research on factors influencing the quality of frozen seafood. Fish. Sci. 2020, 86, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Tolstorebrov, I.; Eikevik, T.M.; Bantle, M. Effect of low and ultra-low temperature applications during freezing and frozen storage on quality parameters for fish. Int. J. Refrig. 2016, 63, 37–47. [Google Scholar] [CrossRef]
- Hsieh, R.J.; Kinsella, J.E. Lipoxygenase generation of specific volatile flavor carbonyl compounds in fish tissues. J. Agric. Food Chem. 1989, 37, 279–286. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [Green Version]
- Dunkel, A.; Steinhaus, M.; Kotthoff, M.; Nowak, B.; Krautwurst, D.; Schieberle, P.; Hofmann, T. Nature’s chemical signatures in human olfaction: A foodborne perspective for future biotechnology. Angew. Chem. Int. Ed. 2014, 53, 7124–7143. [Google Scholar] [CrossRef]
- Ratel, J.; Engel, E. Determination of benzenic and halogenated volatile organic compounds in animal-derived food products by one-dimensional and comprehensive two-dimensional gas chromatography-mass spectrometry. J. Chromatogr. A 2009, 1216, 7889–7898. [Google Scholar] [CrossRef]
- Wang, W.; Feng, X.; Zhang, D.; Li, B.; Sun, B.; Tian, H.; Liu, Y. Analysis of volatile compounds in Chinese dry-cured hams by comprehensive two-dimensional gas chromatography with high-resolution time-of-flight mass spectrometry. Meat Sci. 2018, 140, 14–25. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, Y.; Zhang, J.; Wang, X.; Shi, W. Characteristic volatile compounds in different parts of grass carp by comprehensive two-dimensional gas chromatography/time-of-flight mass spectrometry. Int. J. Food Prop. 2011, 23, 777–796. [Google Scholar] [CrossRef]
- Giri, A.; Khummueng, W.; Mercier, F.; Kondjoyan, N.; Tournayre, P.; Meurillon, M.; Ratel, J.; Engel, E. Relevance of two-dimensional gas chromatography and high resolution olfactometry for the parallel determination of heat-induced toxicants and odorants in cooked food. J. Chromatogr. A 2015, 1388, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Bao, Y.; Wang, K.; Yin, L.; Zhou, P. Protein changes in shrimp (Metapenaeus ensis) frozen stored at different temperatures and the relation to water-holding capacity. Int. J. Food Sci. Techol. 2021, 56, 3924–3937. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, B.S. Freezing tilapia by airblast and liquid nitrogen—Freezing point and freezing rate. Int. J. Food Sci. Technol. 1995, 30, 167–173. [Google Scholar] [CrossRef]
- Qu, J.; Zhang, L.; Yin, L.; Liu, J.; Sun, Z.; Zhou, P. Changes in bioactive proteins and serum proteome of human milk under different frozen storage. Food Chem. 2021, 352, 129436. [Google Scholar] [CrossRef] [PubMed]
- Offer, G.; Knight, P. Part 1: General principles and water uptake in meat processing. In The Structural Basis of Water-Holding in Meat; Lawrie, R., Ed.; Developments in Meat Science; Elsevier Applied Science: London, UK, 1988; Volume 4, pp. 63–171. [Google Scholar]
- Hughes, J.; Oiseth, S.; Purslow, P.; Warner, R. A structural approach to understanding the interactions between color, water-holding capacity and tenderness. Meat Sci. 2014, 98, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Zuanazzi, J.S.G.; Goes, E.S.d.R.; Almeida, F.L.A.d.; Goes, M.D.; Lara, J.A.F.d.; Ribeiro, R.P. Effects of freezing and thawing cycles on the quality of Nile tilapia fillets. Food Sci. Technol. 2020, 40 (Suppl. 1), 300–304. [Google Scholar] [CrossRef] [Green Version]
- Du, N.; Sun, Y.; Chen, Z.; Huang, X.; Li, C.; Gao, L.; Bai, S.; Wang, P.; Hao, Q. Effects of Multiple Freeze-Thaw Cycles on Protein and Lipid Oxidation, Microstructure and Quality Characteristics of Rainbow Trout (Oncorhynchus mykiss). Fishes 2023, 8, 108. [Google Scholar] [CrossRef]
- Ngapo, T.M.; Babare, I.H.; Reynolds, J.; Mawson, R.F. Freezing rate and frozen storage effects on the ultrastructure of samples of pork. Meat Sci. 1999, 53, 159–168. [Google Scholar] [CrossRef]
- Bevilacqua, A.E.; Zaritzky, N.E. Ice Recrystallization in Frozen Beef. J. Food Sci. 1982, 47, 1410–1414. [Google Scholar] [CrossRef]
- Baron, C.P.; Kjaersgard, I.V.H.; Jessen, F.; Jacobsen, C. Protein and lipid oxidation during frozen storage of rainbow trout (Oncorhynchus mykiss). J. Agric. Food Chem. 2007, 55, 8118–8125. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Cheng, Y.; Kang, D.; Zhang, W.; Zhou, G. Quality changes of pork during frozen storage: Comparison of immersion solution freezing and air blast freezing. Int. J. Food Sci. Techol. 2020, 55, 109–118. [Google Scholar] [CrossRef]
- Huang, L.; Xiong, Y.L.; Kong, B.; Huang, X.; Li, J. Influence of storage temperature and duration on lipid and protein oxidation and flavour changes in frozen pork dumpling filler. Meat Sci. 2013, 95, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Ozen, B.O.; Soyer, A. Effect of plant extracts on lipid and protein oxidation of mackerel (Scomber scombrus) mince during frozen storage. J. Food Sci. Techol. 2018, 55, 120–127. [Google Scholar] [CrossRef]
- Richards, M.P. Lipid chemistry and biochemistry. In Handbook of Food Science, Technology, and Engineering; Hui, Y., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Boca Raton, FL, USA, 2006; Volume 8, pp. 1–21. [Google Scholar]
- Calvelo, A. Recent studies on meat freezing. In Developments in Meat Science; Lawrie, R., Ed.; Elsevier Applied Science: London, UK, 1981; Volume 2, pp. 125–158. [Google Scholar]
- Love, R.M. Studies on protein denaturation in frozen fish. III.—The mechanism and site of denaturation at low temperatures. J. Sci. Food Agric. 1958, 9, 609–617. [Google Scholar] [CrossRef]
- Karlsdottir, M.G.; Sveinsdottir, K.; Kristinsson, H.G.; Villot, D.; Craft, B.D.; Arason, S. Effects of temperature during frozen storage on lipid deterioration of saithe (Pollachius virens) and hoki (Macruronus novaezelandiae) muscles. Food Chem. 2014, 156, 234–242. [Google Scholar] [CrossRef]
- Schaich, K.M.; Shahidi, F.; Zhong, Y.; Eskin, N.A.M. Lipid Oxidation. In Biochemistry of Foods, 3rd ed.; Eskin, N.A.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 419–478. [Google Scholar]
- Leduc, F.; Krzewinski, F.; Le Fur, B.; N’Guessan, A.; Malle, P.; Kol, O.; Duflos, G. Differentiation of fresh and frozen/thawed fish, European sea bass (Dicentrarchus labrax), gilthead seabream (Sparus aurata), cod (Gadus morhua) and salmon (Salmo salar), using volatile compounds by SPME/GC/MS. J. Sci. Food Agric. 2012, 92, 2560–2568. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, G.; Luo, L.; Chen, G. Study on seafood volatile profile characteristics during storage and its potential use for freshness evaluation by headspace solid phase microextraction coupled with gas chromatography–mass spectrometry. Anal. Chim. Acta 2010, 659, 151–158. [Google Scholar] [CrossRef]
- Barriuso, B.; Astiasarán, I.; Ansorena, D. A review of analytical methods measuring lipid oxidation status in foods: A challenging task. Eur. Food Res. Technol. 2012, 236, 1–15. [Google Scholar] [CrossRef]
- Pan, N.; Dong, C.; Du, X.; Kong, B.; Sun, J.; Xia, X. Effect of freeze-thaw cycles on the quality of quick-frozen pork patty with different fat content by consumer assessment and instrument-based detection. Meat Sci. 2021, 172, 108313. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Ganhao, R.; Estevez, M. Role of Phenolics Extracting from Rosa canina L. on Meat Protein Oxidation During Frozen Storage and Beef Patties Processing. Food Bioprocess Technol. 2015, 8, 854–864. [Google Scholar] [CrossRef]
- Zheng, Y.; Qiu, Z.; Wang, X. Protein oxidation and tandem mass tag-based proteomic analysis in the dorsal muscle of farmed obscure pufferfish subjected to multiple freeze–thaw cycles. J. Food Process. Preserv. 2020, 44, e14721. [Google Scholar] [CrossRef]
- Qian, S.; Li, X.; Wang, H.; Mehmood, W.; Zhang, C.; Blecker, C. Effects of frozen storage temperature and duration on changes in physicochemical properties of beef myofibrillar protein. J. Food Qual. 2021, 2021, 8836749. [Google Scholar] [CrossRef]
- Bao, Y.; Ertbjerg, P. Relationship between oxygen concentration, shear force and protein oxidation in modified atmosphere packaged pork. Meat Sci. 2015, 110, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Chelh, I.; Gatellier, P.; Santé-Lhoutellier, V. A simplified procedure for myofibril hydrophobicity determination. Meat Sci. 2006, 74, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Wang, K.; Yang, H.; Regenstein, J.M.; Ertbjerg, P.; Zhou, P. Protein degradation of black carp (Mylopharyngodon piceus) muscle during cold storage. Food Chem. 2020, 308, 125576. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, M.; Liu, Z.; Chen, S.; Xu, Y. Three extraction methods in combination with GC× GC-TOFMS for the detailed investigation of volatiles in Chinese herbaceous aroma-type baijiu. Molecules 2020, 25, 4429. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Time t | Freezing Rate FR | Temperature ST | t ∗ FR | t ∗ ST |
|---|---|---|---|---|---|
| TVBN | *** | NS | NS | NS | NS |
| TBARS | *** | *** | *** | ** | *** |
| Water-holding of myofibrils | NS | NS | * | * | NS |
| Thaw loss | *** | NS | NS | NS | NS |
| Thiols | NS | ** | NS | NS | NS |
| Surface hydrophobicity | NS | *** | *** | NS | NS |
| Compounds | Freezing Rate | Storage Temperature | Storage Time |
|---|---|---|---|
| Hexanoic acid | NS | NS | ** |
| Heptanoic acid | NS | NS | * |
| Pentadecanal | * | NS | NS |
| Octanoic acid | NS | NS | NS |
| 2-Propenal, 3-phenyl- | NS | NS | NS |
| Nonanoic acid | NS | NS | ** |
| n-Decanoic acid | NS | NS | NS |
| Heptanal | *** | *** | NS |
| 2-Hexenal | *** | *** | NS |
| 2-Pentanone | *** | *** | *** |
| 1-Pentanol | *** | *** | *** |
| 1-Heptadecanol | NS | *** | *** |
| 2-Butanone, 3-methyl- | NS | *** | ** |
| 2-Butanone, 3-hydroxy- | NS | ** | * |
| Octanal | NS | NS | NS |
| 2,3-Octanedione | * | * | NS |
| 5-Hepten-2-one, 6-methyl- | NS | NS | NS |
| 1-Hexanol | *** | *** | ** |
| 3-Heptanol, 6-methyl- | NS | NS | NS |
| 2-Nonanone | NS | NS | * |
| Nonanal | NS | NS | NS |
| 2-Octenal | NS | NS | NS |
| 1-Octen-3-ol | * | * | NS |
| 1-Heptanol | *** | *** | ** |
| Decanal | NS | ** | * |
| 2,4-Heptadienal | *** | *** | NS |
| (5Z)-Octa-1,5-dien-3-ol | *** | *** | ** |
| 4-Hepten-1-ol | *** | *** | NS |
| 1-Hexanol, 2-ethyl- | NS | NS | * |
| 2-Nonenal | NS | NS | NS |
| 1-Octanol | NS | NS | * |
| 2,7-Octadien-1-ol | *** | *** | NS |
| Undecanal | *** | *** | NS |
| 3-Undecanone | *** | NS | ** |
| 2-Undecanone | NS | * | NS |
| 2-Octen-1-ol | ** | NS | NS |
| 2-Undecanone | NS | * | NS |
| 1-Nonanol | NS | * | * |
| 11-Dodecenol | *** | *** | NS |
| 2(5H)-furanone | NS | NS | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Y.; Zhang, Y.; Xu, W. Effects of Different Freezing Rate and Frozen Storage Temperature on the Quality of Large-Mouth Bass (Micropterus salmoides). Molecules 2023, 28, 5432. https://doi.org/10.3390/molecules28145432
Bao Y, Zhang Y, Xu W. Effects of Different Freezing Rate and Frozen Storage Temperature on the Quality of Large-Mouth Bass (Micropterus salmoides). Molecules. 2023; 28(14):5432. https://doi.org/10.3390/molecules28145432
Chicago/Turabian StyleBao, Yulong, Yaqi Zhang, and Wanjun Xu. 2023. "Effects of Different Freezing Rate and Frozen Storage Temperature on the Quality of Large-Mouth Bass (Micropterus salmoides)" Molecules 28, no. 14: 5432. https://doi.org/10.3390/molecules28145432
APA StyleBao, Y., Zhang, Y., & Xu, W. (2023). Effects of Different Freezing Rate and Frozen Storage Temperature on the Quality of Large-Mouth Bass (Micropterus salmoides). Molecules, 28(14), 5432. https://doi.org/10.3390/molecules28145432