
Theoretical and Experimental Studies on the Evidence of 1,3-β-Glucan in Marennine of Haslea ostrearia
 , , , , ,
, , , , ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Computational Study
2.2. Enzymatic Reactions
2.3. Spectroscopic Analysis
2.4. Colorimetric Assay
3. Discussion
4. Materials and Methods
4.1. Computational Study
4.1.1. Model Construction
4.1.2. Molecular Docking
4.1.3. Molecular Dynamics (MD) Simulation and Binding Energy Calculation
4.2. Materials
4.3. Purification of Marennine
4.4. The Carbohydrate Hydrolytic Enzymes Assay
4.4.1. Detection of Carbohydrate Contents (DNS Assays)
4.4.2. Thin-Layer Chromatography
4.5. Nuclear Magnetic Resonance Spectroscopy
4.6. Mass Spectroscopy
4.7. Colorimetric Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bandaranayake, W.M. The nature and role of pigments of marine invertebrates. Nat. Prod. Rep. 2006, 23, 223–255. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, C.; Liu, B.; Lin, L.; Sarker, S.D.; Nahar, L.; Xiao, J. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Trends Food Sci. Technol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Microalgae for aquaculture: Opportunities and constraints. J. Appl. Phycol. 1997, 9, 393–401. [Google Scholar] [CrossRef]
- Wrede, F.; Strack, E. Über das Pyocyanin, den blauen Farbstoff des Bacillus Pyocyaneus. IV. Die Konstitution und Synthese des Pyocyanins Hoppe-Seyler’s Z. Physiol. Chem. 1929, 181, 58–76. [Google Scholar] [CrossRef]
- Frank, L.H.; DeMoss, R.D. On the biosynthesis of pyocyanine. J. Bacteriol. 1959, 77, 776. [Google Scholar] [CrossRef]
- Hassan, H.M.; Fridovich, I. Mechanism of the antibiotic action pyocyanine. J. Bacteriol. 1980, 141, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, H.; Akimoto, R. New blue pigment produced by Pantoea agglomerans and its production characteristics at various temperatures. Appl. Environ. Microbiol. 2011, 77, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Fenical, W. Chemical studies of marine bacteria: Developing a new resource. Chem. Rev. 1993, 93, 1673–1683. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Thorwest, M.; Plitzko, I.; Brinkhoff, T.; Simon, M.; Zeeck, A. Production of a blue pigment (glaukothalin) by marine Rheinheimera spp. Int. J. Microbiol. 2009, 2009, 701735. [Google Scholar] [CrossRef] [Green Version]
- Brettar, I.; Christen, R.; Höfle, M.G. Rheinheimera baltica gen. nov., sp. nov., a blue-coloured bacterium isolated from the central Baltic Sea. Int. J. Syst. Evol. Microbiol. 2002, 52, 1851–1857. [Google Scholar]
- Kai, A.; Yoshii, Y.; Nakayama, T.; Inouye, I. Aurearenophyceae classis nova, a new class of Heterokontophyta based on a new marine unicellular alga Aurearena cruciata gen. et sp. nov. inhabiting sandy beaches. Protist 2008, 159, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.-M.; Morançais, M.; Pradier, E.; Mouget, J.L.; Tremblin, G. Extraction and quantitative analysis of the blue-green pigment “marennine” synthesized by the diatom Haslea ostrearia. J. Appl. Phycol. 2002, 14, 299–305. [Google Scholar] [CrossRef]
- Gastineau, R.; Turcotte, F.; Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Windarto, E.; Mouget, J.-L. Marennine, promising blue pigments from a widespread Haslea diatom species complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [Green Version]
- Prasetiya, F.S. Greening Phenomenon in Bivalve by Marennine Produced from Haslea ostrearia and Its Consequences on Bivalve’s Integrated Response. Universite du Maine. Available online: https://theses.hal.science/tel-01279527 (accessed on 19 April 2020).
- Gastineau, R.; Davidovich N a Bardeau, J.F.; Caruso a Leignel, V.; Hardivillier, Y.; Mouget, J.L. Haslea karadagensis (Bacillariophyta): A second blue diatom, recorded from the Black Sea and producing a novel blue pigment. Eur. J. Phycol. 2012, 47, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Prasetiya, F.S.; Gastineau, R.; Poulin, M.; Lemieux, C.; Turmel, M.; Syakti, A.D.; Leignel, V. Haslea nusantara (Bacillariophyceae), a new blue diatom from the Java Sea, Indonesia: Morphology, biometry and molecular characterization. Plant Ecol. Evol. 2019, 151, 188–202. [Google Scholar] [CrossRef]
- Gastineau, R.; Pouvreau, J.B.; Hellio, C.; Morançais, M.; Fleurence, J.; Gaudin, P.; Mouget, J.L. Biological activities of purified marennine, the blue pigment responsible for the greening of oysters. J. Agric. Food Chem. 2012, 60, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Gastineau, R.; Hardivillier, Y.; Leignel, V.; Tekaya, N.; Morançais, M.; Fleurence, J.; Mouget, J.L. Greening effect on oysters and biological activities of the blue pigments produced by the diatom Haslea karadagensis (Naviculaceae). Aquaculture 2012, 368–369, 61–67. [Google Scholar] [CrossRef]
- Prasetiya, F.S.; Sunarto, S.; Bachtiar, E.; Agung MU, K.; Nathanael, B.; Pambudi, A.C.; Mouget, J. Effect of the blue pigment produced by the tropical diatom Haslea nusantara on marine organisms from different trophic levels and its bioactivity. Aquac. Rep. 2020, 17, 100389. [Google Scholar] [CrossRef]
- Lankester, E.R. On green oysters. Q. J. Microsc. Sci. 1886, 26, 71–94. [Google Scholar] [CrossRef] [Green Version]
- Valenciennes, A. Sur les causes de la coloration en vert de certaines huîtres. Comptes Rendus Hebd. Seances L’académie Sci. 1841, 12, 345–346. [Google Scholar]
- Boyce, R.W.; Herdman, W.A. The possible infectivity of the oyster, and upon the green disease in oysters. Comm. Br. Assoc. Adv. Sci. 1896, 66, 663–669. [Google Scholar]
- Mitchell, P.H.; Barney, R.L. The occurrence in Virginia of green-gilled oysters similar to those of Marennes. Bull. US Bur. Fish. 1918, 35, 135–149. [Google Scholar]
- Ranson, G. Le verdissement des huitres. Science 1937, 8, 13–24. [Google Scholar]
- Robert, J.M. Fertilite des Claires Ostre!Icoles et Verdissement: Utilisation de L’azote par les Diatomees Dominantes. Ph.D. Thesis, University of Nantes, Nantes, France, 1983; p. 282. [Google Scholar]
- Bachrach, E. Le bleuissement des diatomées et le verdissement des huîtres-revue critique. Rev. Trav. L’institut Pêches Marit. 1935, 8, 112–123. [Google Scholar]
- Ranson, G. L’absorption de matières organiques dissoutes par la surface expérieure du corps chez les animaux aquatiques. Annales l’Institut Océanographique 1927, IV, 49–174. [Google Scholar]
- Bachrach, E.; Jouvent, E. Sur la Pigmentation Bleue de Certaines Diatomées. Bull. Mens. Soc. Linnéenne Lyon 1933, 2, 113–116. [Google Scholar]
- Bachrach, E.; Simonet, M. Le phénomène du bleuissement chez les diatomées. Rev. Trav. L’institut Pêches Marit. 1936, 9, 113–116. [Google Scholar]
- Moreau, J. Recherches préliminaires sur le verdissement en claires: L’évolution de leurs divers pigments liée au complexe pigmentaire de Navicula ostrearia Bory. Revue des travaux de l’Institut des pêches maritimes. Rev. Trav. L’institut Pêches Marit. 1967, 31, 372–382. [Google Scholar]
- Ryder, J.A. On the cause of the greening of oysters. With a supplementary note on the coloration of the blood corpuscles of the oysters. In Report of Commission of Fish and Fisheries for 1882; Government Printing Office Washington: Washington, DC, USA, 1884; Volume 10, pp. 793–800. [Google Scholar]
- Robert, M.J.; Hallet, J.N. Absorptio Spectrum in vivo of the Blue Pigment ‘Marennine’of the Pennate Diatom Navicula ostrearia Bory. J. Exp. Bot. 1981, 32, 341–345. [Google Scholar] [CrossRef]
- Bocat, L. Sur la Marennine de la Diatomée Bleue; Comparaison Avec la Phycocyanine. Comptes Rendus Soc. Biol. 1907, 62, 1073–1075. [Google Scholar]
- Neuville, D.; Daste, P. Production de pigment bleu par la Diatomée Navicula ostrearia (Gaillon) Bory, maintenue en culture unialgale sur un milieu synthétique carencé en azote nitrique. Comptes Rendus L’académie Sci. Paris Série D 1972, 274, 2030–2033. [Google Scholar]
- Hardouin, V.; Vandanjon, L.; Jaouen, P.; Robert, J.M. Procédé combiné extraction-membranes pour l’isolement et la purification de pigments naturels. In Actes Colloque Interfiltra Intermembranes, Paris; European Federation of Chemical Engineering: Paris, France, 1994; pp. 175–181. [Google Scholar]
- Pouvreau, J.B.; Morançais, M.; Massé, G.; Rosa, P.; Robert, J.M.; Fleurence, J.; Pondaven, P. Preliminary characterisation of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 757–767. [Google Scholar] [CrossRef]
- Zebiri, I.; Jacquette, B.; Francezon, N.; Herbaut, M.; Latigui, A.; Bricaud, S.; Dittmer, J. The Polysaccharidic Nature of the Skeleton of Marennine as Determined by NMR Spectroscopy. Mar. Drugs 2023, 21, 42. [Google Scholar] [CrossRef] [PubMed]
- Gügi, B.; Le Costaouec, T.; Burel, C.; Lerouge, P.; Helbert, W.; Bardor, M. Diatom-Specific Oligosaccharide and Polysaccharide Structures Help to Unravel Biosynthetic Capabilities in Diatoms. Mar. Drugs 2015, 13, 5993–6018. [Google Scholar] [CrossRef] [Green Version]
- Pouvreau, J.B.; Morançais, M.; Massé, G.; Rosa, P.; Robert, J.M.; Fleurence, J.; Pondaven, P. Purification of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 769–781. [Google Scholar] [CrossRef]
- Gastineau, R.; Prasetiya, F.S.; Falaise, C.; Cognie, B.; Decottignies, P.; Morançais, M.; Mouget, J.-L. Marennine-like Pigments: Blue Diatom or Green Oyster Cult? In Blue Biotechnology: Production and Use of Marine Molecules, 1st ed.; La Barre, S., Bates, S.S., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018; Volume 1, pp. 529–551. [Google Scholar] [CrossRef]
- Young, S.H.; Dong, W.J.; Jacobs, R.R. Observation of a partially opened triple-helix conformation in 1→3-β- glucan by fluorescence resonance energy transfer spectroscopy. J. Biol. Chem. 2000, 275, 11874–11879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semedo, M.C.; Karmali, A.; Fonseca, L. A high throughput colorimetric assay of β-1,3-d-glucans by Congo red dye. J. Microbiol. Methods 2015, 109, 140–148. [Google Scholar] [CrossRef]
- Abraham, R.J.; Mobli, M. Modelling 1H NMR Spectra of Organic Compounds: Theory, Applications and NMR Prediction Software; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Berman ES, F.; Kulp, K.S.; Knize, M.G.; Wu, L.; Nelson, E.J.; Nelson, D.O.; Wu, K.J. Distinguishing monosaccharide stereo- and structural isomers with TOF-SIMS and multivariate statistical analysis. Anal. Chem. 2006, 78, 6497–6503. [Google Scholar] [CrossRef] [Green Version]
- Lehtovaara, B.C.; Gu, F.X. Pharmacological, structural, and drug delivery properties and applications of 1,3-β-glucans. J. Agric. Food Chem. 2011, 59, 6813–6828. [Google Scholar] [CrossRef]
- Legentil, L.; Paris, F.; Ballet, C.; Trouvelot, S.; Daire, X.; Vetvicka, V.; Ferrières, V. Molecular interactions of β-(1→3)-glucans with their receptors. Molecules 2015, 20, 9745–9766. [Google Scholar] [CrossRef]
- Qin, Z.; Yang, D.; You, X.; Liu, Y.; Hu, S.; Yan, Q.; Jiang, Z. The recognition mechanism of triple-helical β-1,3-glucan by a β-1,3-glucanase. Chem. Commun. 2017, 53, 9368–9371. [Google Scholar] [CrossRef]
- Pouvreau, J.B.; Morançais, M.; Taran, F.; Rosa, P.; Dufossé, L.; Guérard, F.; Pondaven, P. Antioxidant and free radical scavenging properties of marennine, a blue-green polyphenols pigment from the diatom Haslea ostrearia (Gaillon/Bory) Simonsen responsible for the natural greening of cultured oysters. J. Agric. Food Chem. 2008, 56, 6278–6286. [Google Scholar] [CrossRef] [PubMed]
- Chuah, C.T.; Sarko, A.; Deslandes, Y.; Marchessault, R.H. Triple-Helical Crystalline Structure of Curdlan and Paramylon Hydrates. Macromolecules 1983, 16, 1375–1382. [Google Scholar] [CrossRef]
- Kirschner, K.N.; Yongye, A.B.; Tschampel, S.M.; Gonzalez-Outeirino, J.; Daniels, C.R.; Foley, L.; Woods, R.J. GLYCAM06: A Generalizable Biomolecular Force Field. Carbohydrates. J. Comput. Chem. 2008, 29, 622–655. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.R.; Mcgee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA py: An Efficient Program for End-State Free Energy Calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Falaise, C.; Cormier, P.; Tremblay, R.; Audet, C.; Deschênes, J.S.; Turcotte, F.; Mouget, J.L. Harmful or harmless: Biological effects of marennine on marine organisms. Aquat. Toxicol. 2019, 209, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Mouget, J.L.; Gastineau, R.; Davidovich, O.; Gaudin, P.; Davidovich, N.A. Light is a key factor in triggering sexual reproduction in the pennate diatom Haslea ostrearia. FEMS Microbiol. Ecol. 2009, 69, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, P.J. Specificity in the interaction of direct dyes with polysaccharides. Carbohydr. Res. 1980, 85, 271–287. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
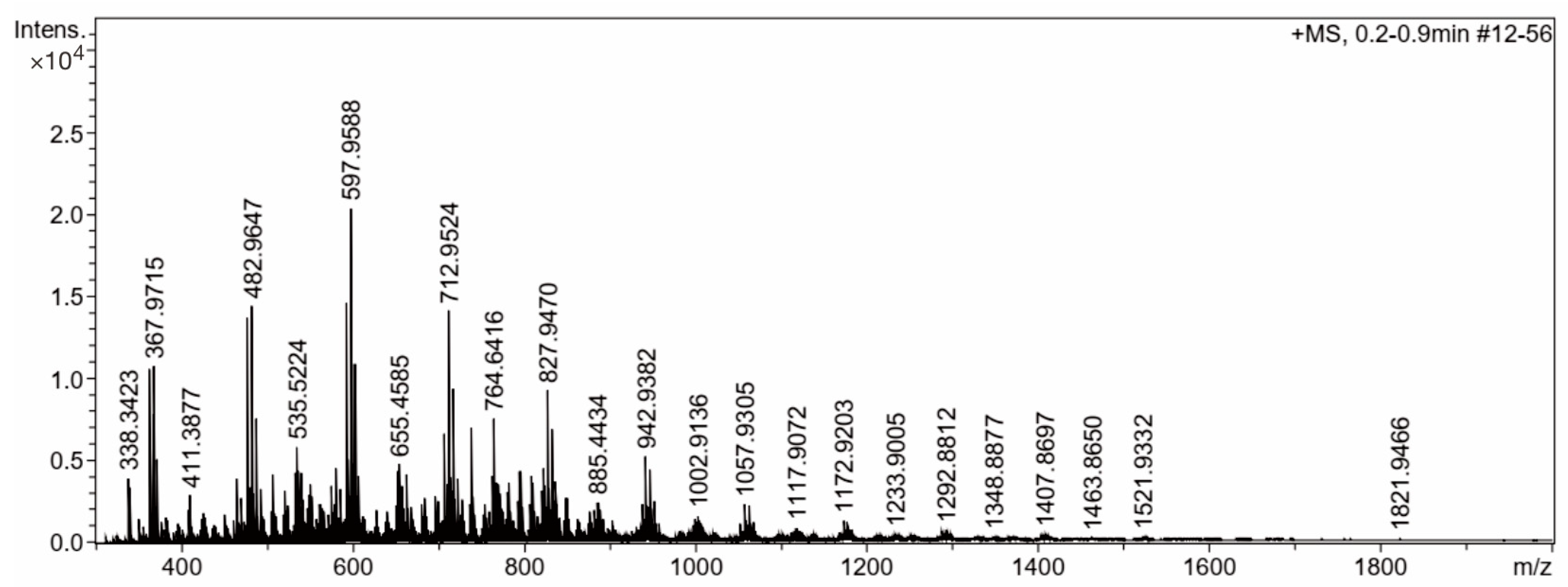{kind=link}
{kind=link}
| m/z | Res. | S/N | I | Area | m/z | Res. | S/N | I | Area |
|---|---|---|---|---|---|---|---|---|---|
| 338.3423 | 7858 | 61.7 | 3964 | 198 | 663.4514 | 8787 | 53.3 | 4257 | 388 |
| 340.2616 | 7269 | 53.4 | 3432 | 184 | 707.9951 | 9313 | 76.5 | 6636 | 625 |
| 363.0163 | 8122 | 165.8 | 10,582 | 541 | 710.4839 | 7626 | 41.7 | 3621 | 386 |
| 367.9715 | 8102 | 169.6 | 10,784 | 557 | 712.9524 | 9273 | 162.5 | 14,136 | 1269 |
| 368.4239 | 7415 | 85.9 | 5460 | 318 | 713.9527 | 8976 | 40.0 | 3480 | 353 |
| 369.3569 | 6621 | 113.1 | 7183 | 536 | 714.9494 | 9167 | 46.0 | 4008 | 377 |
| 372.9262 | 8127 | 80.1 | 5098 | 279 | 715.4331 | 9081 | 36.6 | 3200 | 299 |
| 465.9335 | 8246 | 62.1 | 3940 | 256 | 717.9085 | 9600 | 107.4 | 9419 | 819 |
| 478.0092 | 8300 | 212.8 | 13,685 | 886 | 722.8635 | 9379 | 44.4 | 3936 | 357 |
| 479.4769 | 7682 | 51.1 | 3287 | 264 | 738.6270 | 8533 | 76.0 | 7017 | 702 |
| 480.4846 | 8295 | 52.0 | 3351 | 251 | 739.6300 | 8094 | 37.2 | 3438 | 370 |
| 482.9647 | 8276 | 222.6 | 14,382 | 949 | 762.6233 | 8849 | 42.7 | 4081 | 441 |
| 484.9620 | 8455 | 46.4 | 3004 | 203 | 764.6416 | 8659 | 78.8 | 7577 | 782 |
| 487.9197 | 8431 | 116.8 | 7554 | 495 | 765.6435 | 8608 | 41.0 | 3958 | 417 |
| 493.4906 | 7827 | 50.9 | 3276 | 263 | 766.6724 | 6902 | 37.4 | 3629 | 455 |
| 507.5080 | 7881 | 64.7 | 4187 | 344 | 767.9697 | 9063 | 31.7 | 3081 | 344 |
| 521.5163 | 6961 | 48.3 | 3186 | 289 | 768.6959 | 8135 | 37.2 | 3633 | 415 |
| 533.5245 | 8423 | 65.0 | 4339 | 347 | 770.4509 | 9338 | 36.8 | 3606 | 365 |
| 535.5224 | 6200 | 87.2 | 5832 | 579 | 772.9288 | 9481 | 30.7 | 3026 | 304 |
| 537.9850 | 8686 | 65.2 | 4353 | 331 | 780.6974 | 8191 | 31.5 | 3118 | 400 |
| 540.4665 | 7830 | 64.5 | 4319 | 346 | 782.7118 | 8268 | 36.9 | 3650 | 414 |
| 550.5512 | 7702 | 53.6 | 3606 | 334 | 794.7155 | 8860 | 44.4 | 4443 | 545 |
| 575.9760 | 8884 | 48.7 | 3447 | 262 | 796.7300 | 8753 | 43.8 | 4394 | 536 |
| 580.9268 | 8622 | 63.8 | 4586 | 359 | 808.7304 | 8727 | 40.1 | 4087 | 470 |
| 585.8814 | 8629 | 45.4 | 3286 | 267 | 810.7429 | 8640 | 35.6 | 3635 | 423 |
| 593.0026 | 8843 | 202.2 | 14,650 | 1158 | 820.7454 | 5771 | 30.1 | 3093 | 531 |
| 595.0003 | 8691 | 51.3 | 3718 | 287 | 822.7517 | 8179 | 44.7 | 4594 | 579 |
| 595.4838 | 8419 | 70.5 | 5117 | 440 | 822.9850 | 9277 | 39.4 | 4053 | 452 |
| 597.9588 | 9052 | 279.4 | 20,362 | 1514 | 824.7585 | 8208 | 33.7 | 3467 | 456 |
| 598.9578 | 8835 | 52.5 | 3834 | 313 | 827.9470 | 9632 | 89.7 | 9298 | 945 |
| 599.9560 | 8854 | 66.2 | 4836 | 391 | 828.9453 | 9355 | 30.2 | 3134 | 347 |
| 600.4503 | 6397 | 47.8 | 3498 | 337 | 829.9440 | 9542 | 31.4 | 3268 | 352 |
| 602.9137 | 8932 | 148.4 | 10,889 | 848 | 832.9015 | 9696 | 66.5 | 6947 | 713 |
| 607.8698 | 8968 | 55.7 | 4109 | 325 | 837.8419 | 8148 | 35.6 | 3740 | 429 |
| 652.9772 | 9018 | 55.5 | 4357 | 397 | 942.9382 | 9748 | 49.8 | 5284 | 676 |
| 655.4585 | 8970 | 61.2 | 4828 | 502 | 947.8961 | 9786 | 42.8 | 4527 | 551 |
| 657.9346 | 9103 | 44.5 | 3516 | 313 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yusuf, M.; Baroroh, U.; Nuwarda, R.F.; Prasetiya, F.S.; Ishmayana, S.; Novianti, M.T.; Tohari, T.R.; Hardianto, A.; Subroto, T.; Mouget, J.-L.; et al. Theoretical and Experimental Studies on the Evidence of 1,3-β-Glucan in Marennine of Haslea ostrearia. Molecules 2023, 28, 5625. https://doi.org/10.3390/molecules28155625
Yusuf M, Baroroh U, Nuwarda RF, Prasetiya FS, Ishmayana S, Novianti MT, Tohari TR, Hardianto A, Subroto T, Mouget J-L, et al. Theoretical and Experimental Studies on the Evidence of 1,3-β-Glucan in Marennine of Haslea ostrearia. Molecules. 2023; 28(15):5625. https://doi.org/10.3390/molecules28155625
Chicago/Turabian StyleYusuf, Muhammad, Umi Baroroh, Rina Fajri Nuwarda, Fiddy Semba Prasetiya, Safri Ishmayana, Mia Tria Novianti, Taufik Ramdani Tohari, Ari Hardianto, Toto Subroto, Jean-Luc Mouget, and et al. 2023. "Theoretical and Experimental Studies on the Evidence of 1,3-β-Glucan in Marennine of Haslea ostrearia" Molecules 28, no. 15: 5625. https://doi.org/10.3390/molecules28155625
APA StyleYusuf, M., Baroroh, U., Nuwarda, R. F., Prasetiya, F. S., Ishmayana, S., Novianti, M. T., Tohari, T. R., Hardianto, A., Subroto, T., Mouget, J. -L., & Pasetto, P. (2023). Theoretical and Experimental Studies on the Evidence of 1,3-β-Glucan in Marennine of Haslea ostrearia. Molecules, 28(15), 5625. https://doi.org/10.3390/molecules28155625