


Cytoprotective Effect of Bambusae caulis in Liquamen by Blocking Oxidative Stress in Hepatocytes

Abstract
:1. Introduction
2. Results
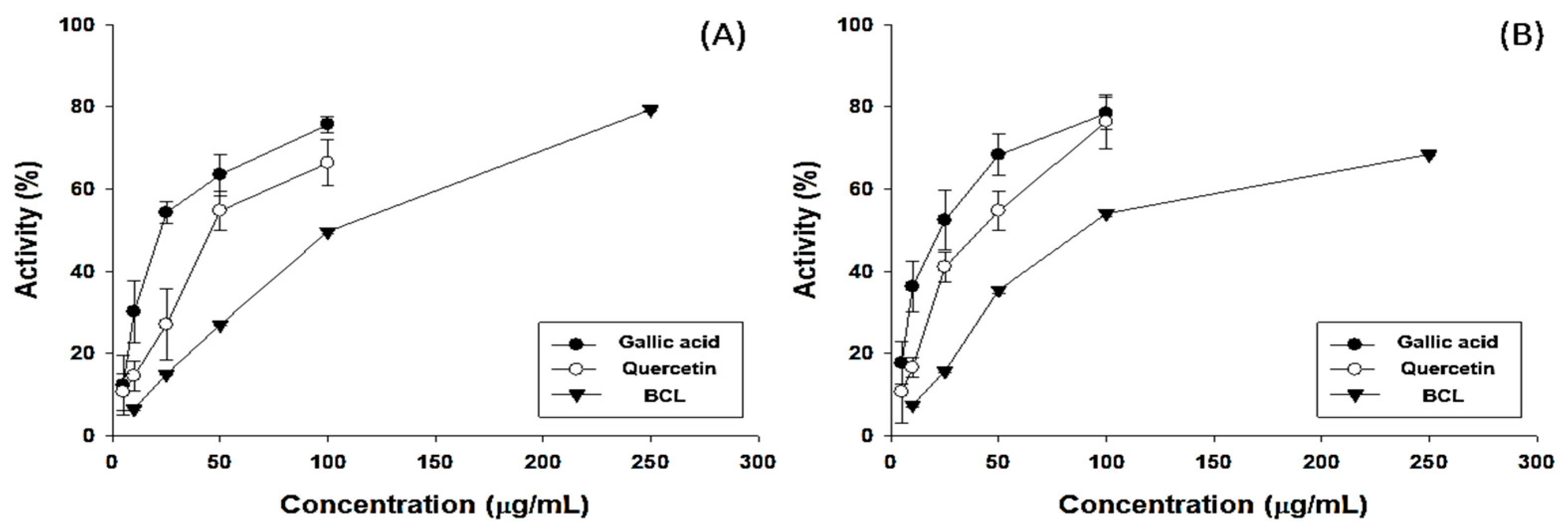
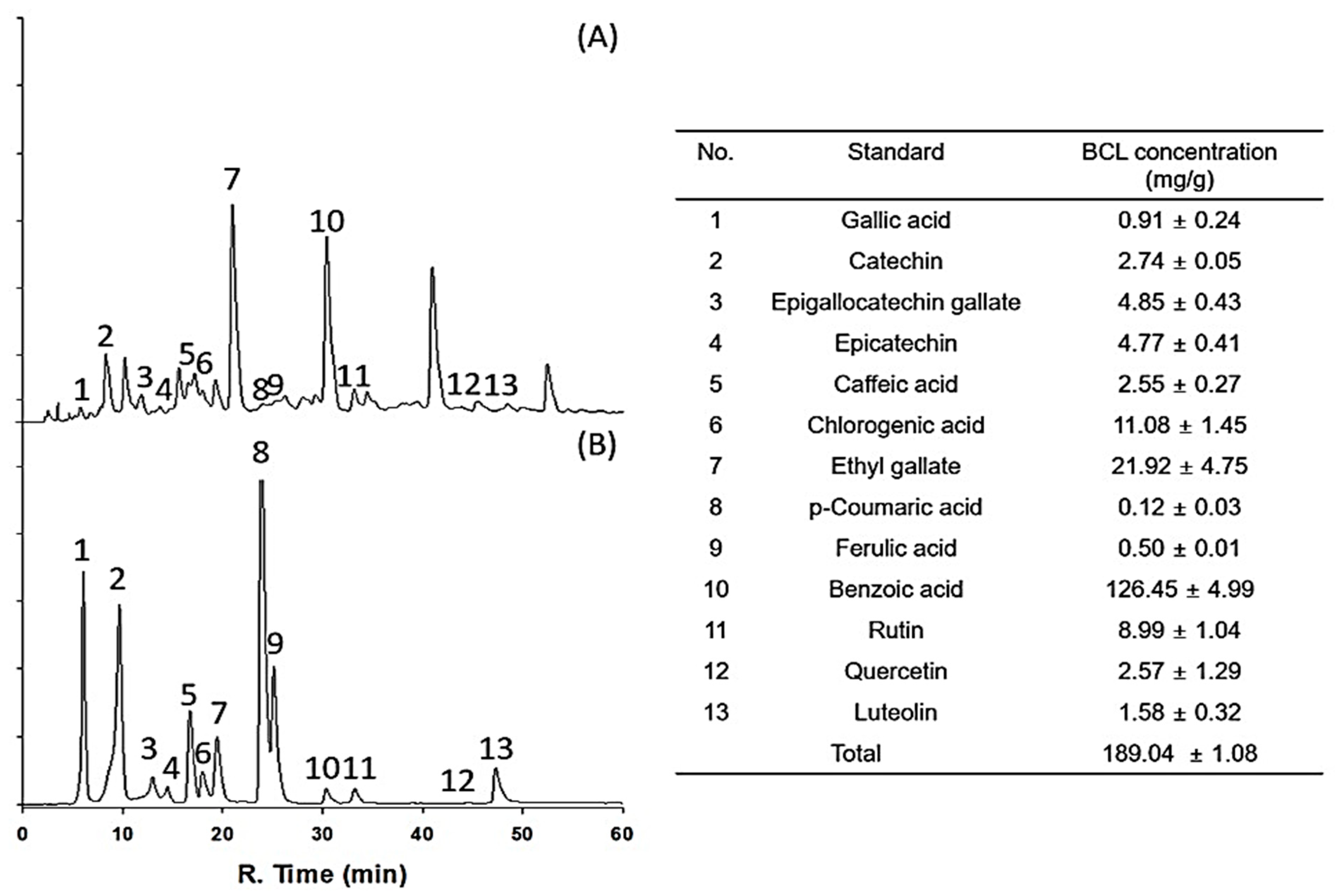
2.1. Antioxidative Potential and Component Analysis of BCL
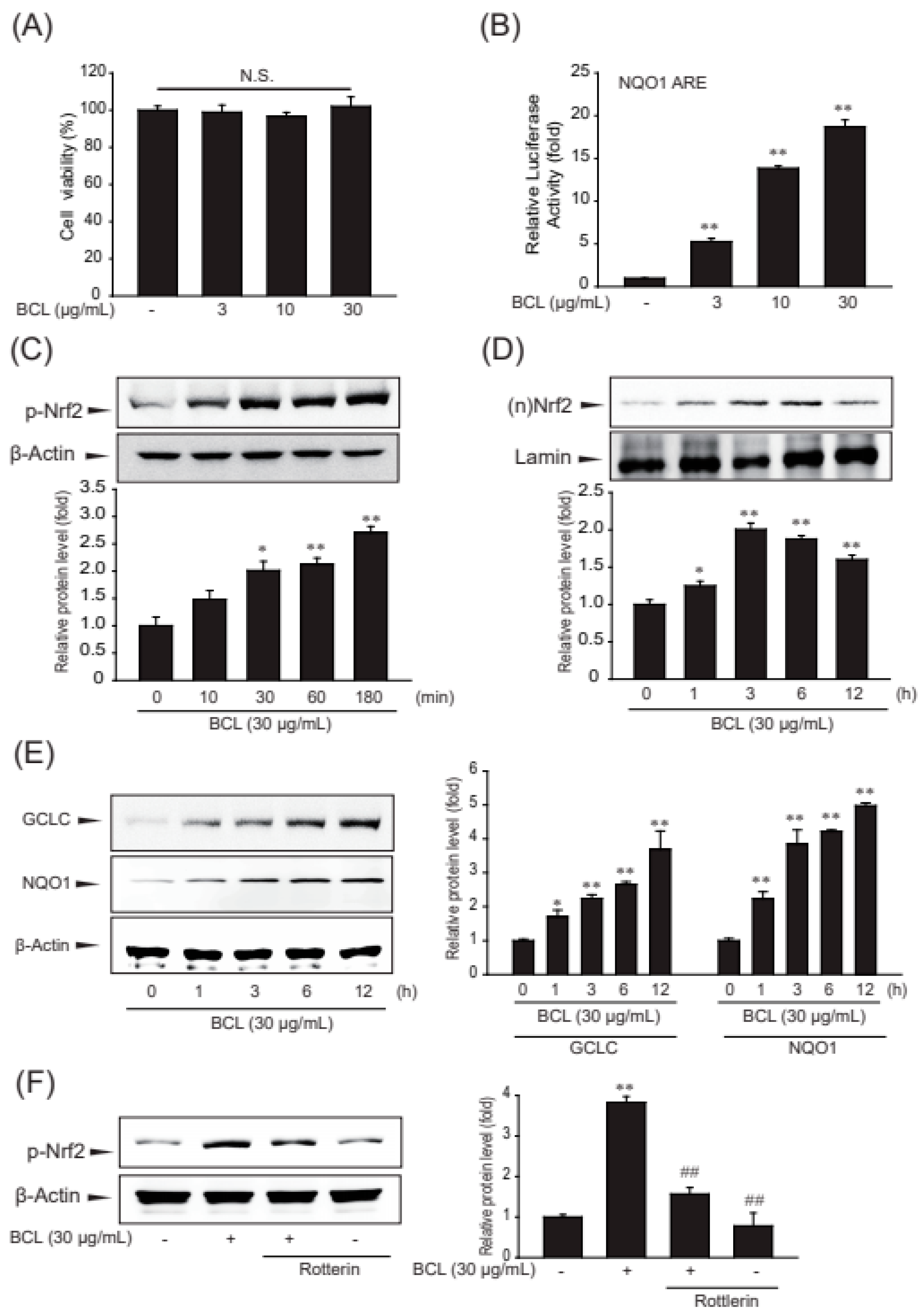
2.2. BCL-Induced Antioxidant Enzymes and Nrf2 Activation in Hepatocytes
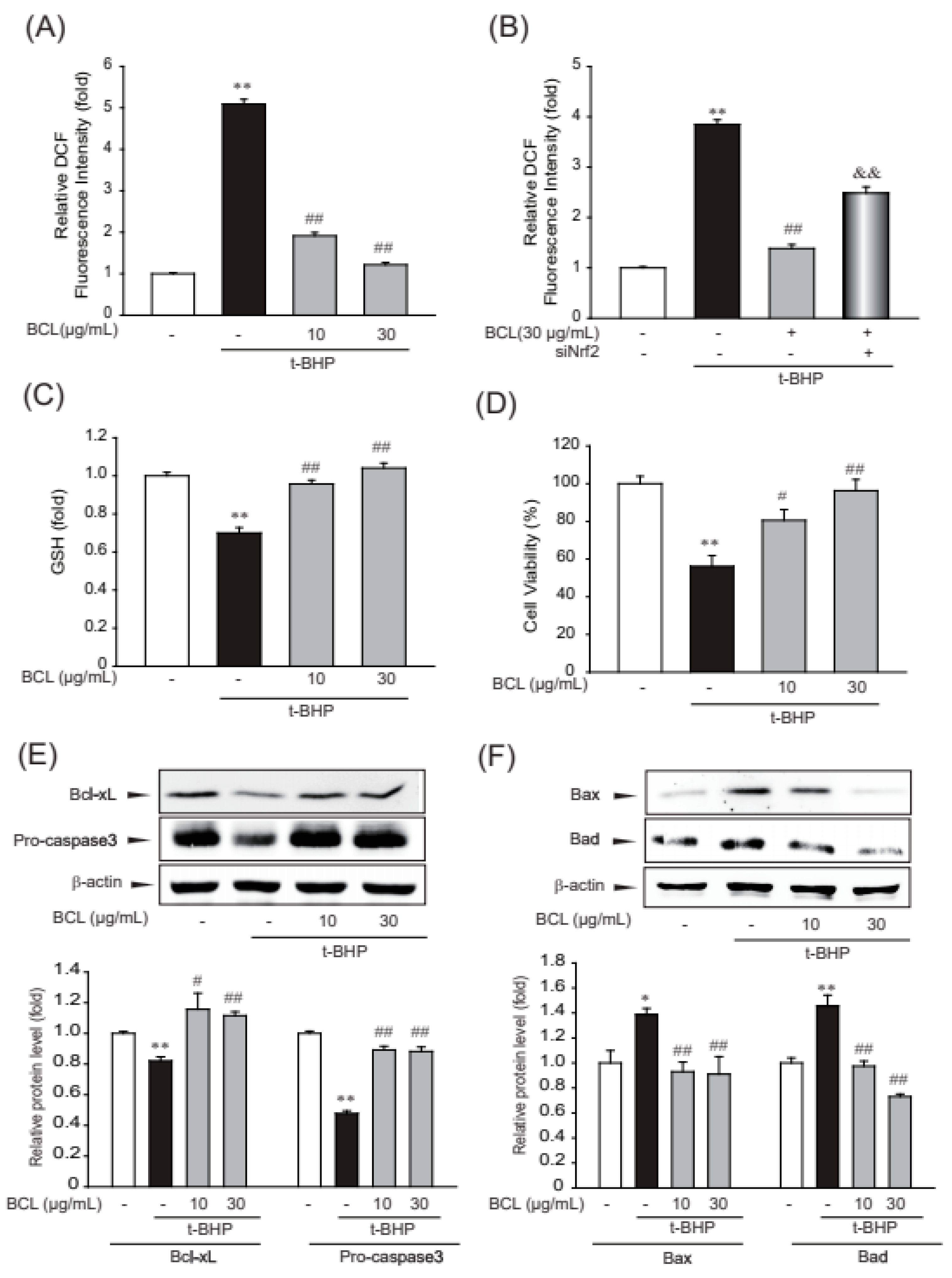
2.3. BCL Suppressed Oxidative Stress and Cell Death Induced by t-BHP
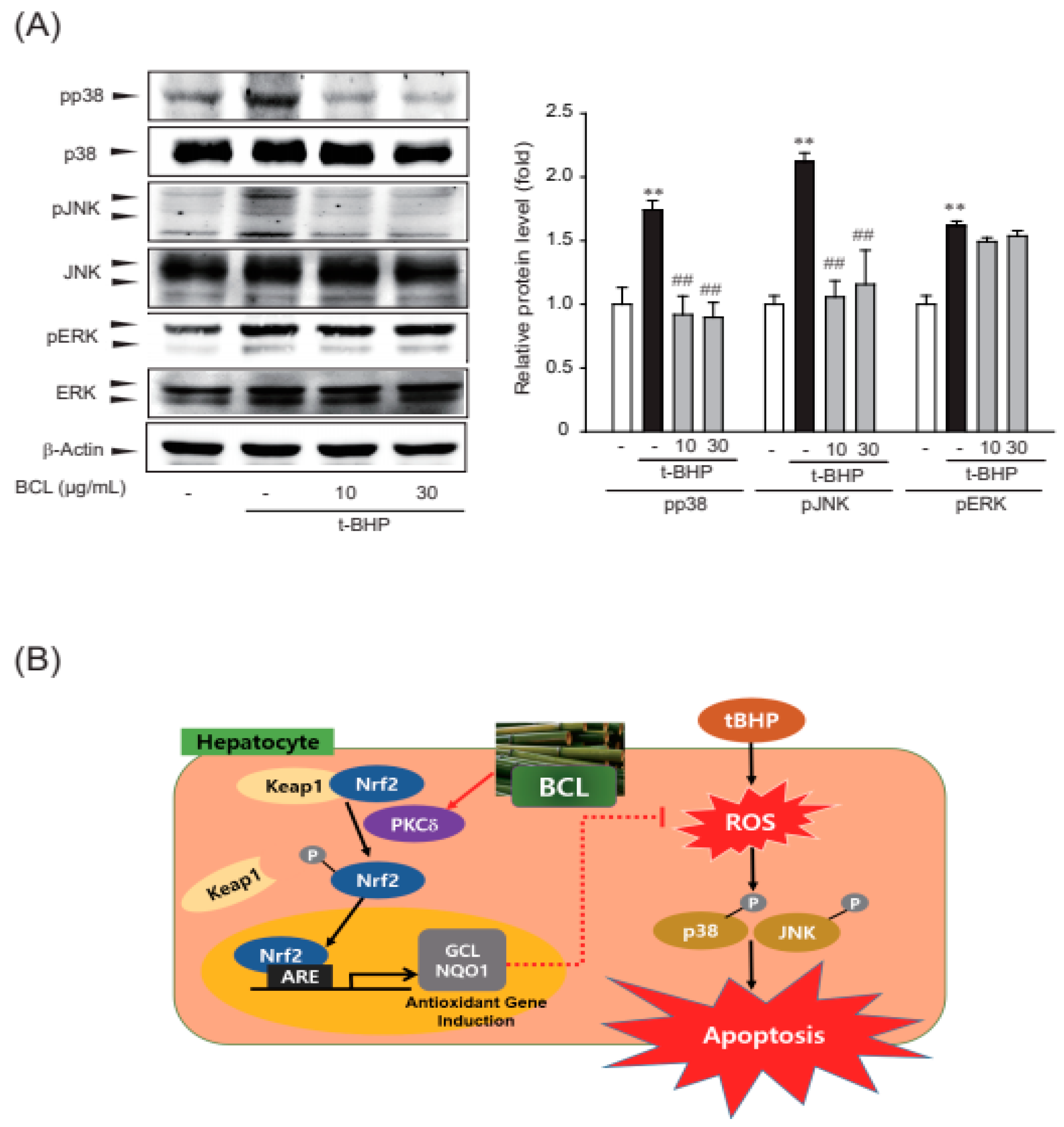
2.4. Effect of BCL on p38 and c-Jun N-terminal Kinase (JNK) Phosphorylation
3. Discussion
4. Materials and Methods
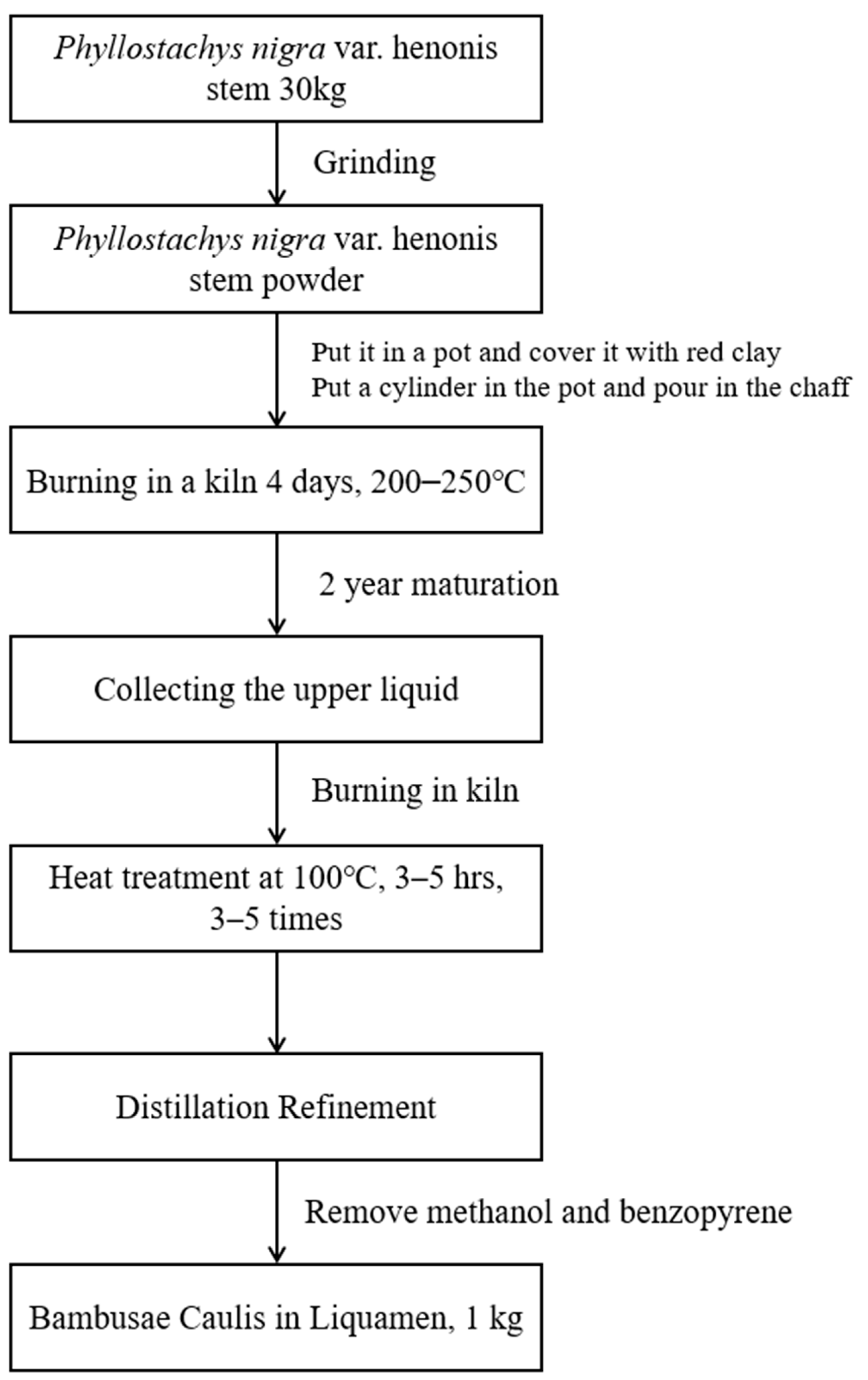
4.1. Preparation of BCL
4.2. Materials
4.3. Radical Scavenging Activity Assay Using DPPH
4.4. Radical Scavenging Activity Assay Using ABTS
4.5. Total Phenolic Content
4.6. Total Flavonoid Content
4.7. Phenolic Compounds Identified in BCL Quantified by HPLC-MS/MS
4.8. HPLC with Diode-Array Detection (HPLC–DAD) Analysis
4.9. Cell Culture
4.10. Cytotoxicity Assay
4.11. Measurement of ROS Generation
4.12. Immunoblot Analysis
4.13. Luciferase Assay
4.14. Statistical Analysis
5. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Mirzaali o’g’li, A.J.; Doniyor Turg’un o’g, X.; Fozil o’g’li, N.A. Liver Structure and Functions, Hepatocytes Information about. Am. J. Econ. Bus. Manag. 2022, 5, 215–216. [Google Scholar]
- Cao, L.; Quan, X.-B.; Zeng, W.-J.; Yang, X.-O.; Wang, M.-J. Mechanism of hepatocyte apoptosis. J. Cell Death 2016, 9, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sifuentes-Franco, S.; Padilla-Tejeda, D.E.; Carrillo-Ibarra, S.; Miranda-Díaz, A.G. Oxidative stress, apoptosis, and mitochondrial function in diabetic nephropathy. Int. J. Endocrinol. 2018, 2018, 1875870. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Gross, A. BCL-2 family proteins as regulators of mitochondria metabolism. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2016, 1857, 1243–1246. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The molecular mechanisms regulating the KEAP1-NRF2 pathway. Mol. Cell. Biol. 2020, 40, e00099-00020. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1–Nrf2 system. Free. Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Vriend, J.; Reiter, R.J. The Keap1-Nrf2-antioxidant response element pathway: A review of its regulation by melatonin and the proteasome. Mol. Cell. Endocrinol. 2015, 401, 213–220. [Google Scholar] [CrossRef]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1–Nrf2–ARE pathway as a potential preventive and therapeutic target: An update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Ghanim, B.Y.; Qinna, N.A. Nrf2/ARE axis signalling in hepatocyte cellular death. Mol. Biol. Rep. 2022, 49, 4039–4053. [Google Scholar] [CrossRef]
- Kučera, O.; Endlicher, R.; Roušar, T.; Lotková, H.; Garnol, T.; Drahota, Z.; Červinková, Z. The effect of tert-butyl hydroperoxide-induced oxidative stress on lean and steatotic rat hepatocytes in vitro. Oxidative Med. Cell. Longev. 2014, 2014, 752506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liese, W.; Kohl, M. Bamboo. In The Plant and Its Uses; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Shukla, R.; Sumit, G.; Sajal, S.; Dwivedi, P.; Mishra, A. Medicinal importance of bamboo. Int. J. Biopharm Phytochem. Res. 2012, 1, 9–15. [Google Scholar]
- Qi, X.F.; Kim, D.H.; Yoon, Y.S.; Song, S.B.; Teng, Y.C.; Cai, D.Q.; Lee, K.J. Bambusae caulis in Liquamen Suppresses the Expression of Thymus and Activation-Regulated Chemokine and Macrophage-Derived Chemokine in Human Keratinocytes due to Antioxidant Effect. Evid. Based Complement Altern. Med. 2012, 2012, 617494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.H.; Lee, M.W. Anti-oxidative, anti-inflammatory and whitening effects of phenolic compounds from Bambusae Caulis in Liquamen. Nat. Prod. Res. 2012, 26, 1687–1691. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Modi, H. Comparative study of DPPH, ABTS and FRAP assays for determination of antioxidant activity. Int. J. Res. Appl. Sci. Eng. Technol. 2015, 3, 636–641. [Google Scholar]
- Ross, D.; Siegel, D. The diverse functionality of NQO1 and its roles in redox control. Redox Biol. 2021, 41, 101950. [Google Scholar] [CrossRef]
- Zhou, L.-l.; Zhou, C.; Liang, X.-w.; Feng, Z.; Liu, Z.-p.; Wang, H.-l.; Zhou, X.-p. Self-protection against triptolide-induced toxicity in human hepatic cells via Nrf2-ARE-NQO1 pathway. Chin. J. Integr. Med. 2017, 23, 929–936. [Google Scholar] [CrossRef]
- Liu, T.; Lv, Y.-F.; Zhao, J.-L.; You, Q.-D.; Jiang, Z.-Y. Regulation of Nrf2 by phosphorylation: Consequences for biological function and therapeutic implications. Free. Radic. Biol. Med. 2021, 168, 129–141. [Google Scholar] [CrossRef]
- Liu, T.; Sun, L.; Zhang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2022, 36, e22942. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalmi, S.K.; Sinha, A.K. ROS mediated MAPK signaling in abiotic and biotic stress-striking similarities and differences. Front. Plant Sci. 2015, 6, 769. [Google Scholar]
- Sangeetha, R.; Diea, Y.; Chaitra, C.; Malvi, P.; Shinomol, G. The amazing bamboo: A review on its medicinal and pharmacological potential. Indian J. Nutr. 2015, 2, 1–7. [Google Scholar]
- Del Olmo, A.; Calzada, J.; Nuñez, M. Benzoic acid and its derivatives as naturally occurring compounds in foods and as additives: Uses, exposure, and controversy. Crit. Rev. Food Sci. Nutr. 2017, 57, 3084–3103. [Google Scholar] [PubMed]
- Khalili, S.T.; Mohsenifar, A.; Beyki, M.; Zhaveh, S.; Rahmani-Cherati, T.; Abdollahi, A.; Bayat, M.; Tabatabaei, M. Encapsulation of Thyme essential oils in chitosan-benzoic acid nanogel with enhanced antimicrobial activity against Aspergillus flavus. LWT-Food Sci. Technol. 2015, 60, 502–508. [Google Scholar]
- Yerlikaya, O.; Gucer, L.; Akan, E.; Meric, S.; Aydin, E.; Kinik, O. Benzoic acid formation and its relationship with microbial properties in traditional Turkish cheese varieties. Food Biosci. 2021, 41, 101040. [Google Scholar]
- Al-Haidari, R.A.; Al-Oqail, M.M. New benzoic acid derivatives from Cassia italica growing in Saudi Arabia and their antioxidant activity. Saudi Pharm. J. 2020, 28, 1112–1117. [Google Scholar] [CrossRef]
- AlSalhi, M.S.; Elangovan, K.; Ranjitsingh, A.J.A.; Murali, P.; Devanesan, S. Synthesis of silver nanoparticles using plant derived 4-N-methyl benzoic acid and evaluation of antimicrobial, antioxidant and antitumor activity. Saudi J. Biol. Sci. 2019, 26, 970–978. [Google Scholar]
- Ross, D.; Siegel, D. NQO1 in protection against oxidative stress. Curr. Opin. Toxicol. 2018, 7, 67–72. [Google Scholar] [CrossRef]
- Berk, M.; Ng, F.; Dean, O.; Dodd, S.; Bush, A.I. Glutathione: A novel treatment target in psychiatry. Trends Pharmacol. Sci. 2008, 29, 346–351. [Google Scholar]
- Hu, Y.; Yu, C.; Yao, M.; Wang, L.; Liang, B.; Zhang, B.; Huang, X.; Zhang, A. The PKCδ-Nrf2-ARE signalling pathway may be involved in oxidative stress in arsenic-induced liver damage in rats. Environ. Toxicol. Pharmacol. 2018, 62, 79–87. [Google Scholar] [PubMed]
- Yue, J.; López, J.M. Understanding MAPK signaling pathways in apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar]
- Liu, J.; Chang, F.; Li, F.; Fu, H.; Wang, J.; Zhang, S.; Zhao, J.; Yin, D. Palmitate promotes autophagy and apoptosis through ROS-dependent JNK and p38 MAPK. Biochem. Biophys. Res. Commun. 2015, 463, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Fu, H.; Gong, W.; Cao, F.; Wu, T.; Hu, F. Plumbagin protects H9c2 cardiomyocytes against TBHP-induced cytotoxicity by alleviating ROS-induced apoptosis and modulating autophagy. Exp. Ther. Med. 2022, 24, 1–8. [Google Scholar]
- Cho, S.S.; Lee, J.H.; Kim, K.M.; Park, E.Y.; Ku, S.K.; Cho, I.J.; Yang, J.H.; Ki, S.H. REDD1 attenuates hepatic stellate cell activation and liver fibrosis via inhibiting of TGF-β/Smad signaling pathway. Free Radic. Biol. Med. 2021, 176, 246–256. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | DPPH (IC50 μg/mL) | ABTS (IC50 μg/mL) | TPC | TFC |
|---|---|---|---|---|
| GAE mg/g | QUE mg/g | |||
| BCL | 99.97 ± 1.23 | 88.17 ± 0.82 | 274.13 ± 15.49 | 352.27 ± 1.28 |
| Gallic acid | 22.30 ± 2.25 | 22.55 ± 1.12 | - | - |
| Quercetin | 46.21 ± 1.72 | 41.97 ± 1.89 | - | - |
| No. | Standard | BCL Concentration (mg/g) |
|---|---|---|
| 1 | 4-Hydroxy benzoic acid | 0.10 |
| 2 | Syringic acid | 0.50 |
| 3 | Coumaric acid | 0.33 |
| 4 | Benzoic acid | 23.14 |
| 5 | Nicotinic acid | 0.14 |
| 6 | Protocatechuic acid | 0.08 |
| 7 | Biochanin A | 0.01 |
| 8 | Catechin | 0.08 |
| 9 | Ethyl gallate | 0.04 |
| 10 | Epigallocatechin gallate | 0.04 |
| Total | 24.47 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.H. Cytoprotective Effect of Bambusae caulis in Liquamen by Blocking Oxidative Stress in Hepatocytes. Molecules 2023, 28, 5862. https://doi.org/10.3390/molecules28155862
Yang JH. Cytoprotective Effect of Bambusae caulis in Liquamen by Blocking Oxidative Stress in Hepatocytes. Molecules. 2023; 28(15):5862. https://doi.org/10.3390/molecules28155862
Chicago/Turabian StyleYang, Ji Hye. 2023. "Cytoprotective Effect of Bambusae caulis in Liquamen by Blocking Oxidative Stress in Hepatocytes" Molecules 28, no. 15: 5862. https://doi.org/10.3390/molecules28155862
APA StyleYang, J. H. (2023). Cytoprotective Effect of Bambusae caulis in Liquamen by Blocking Oxidative Stress in Hepatocytes. Molecules, 28(15), 5862. https://doi.org/10.3390/molecules28155862