

(−)-Epicatechin Inhibits Metastatic-Associated Proliferation, Migration, and Invasion of Murine Breast Cancer Cells In Vitro

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Cell Survival
2.2. Cell Death Bax/Bcl2 Ratio
2.3. Cell Migration Wound Healing
2.4. Cell Invasion Assay
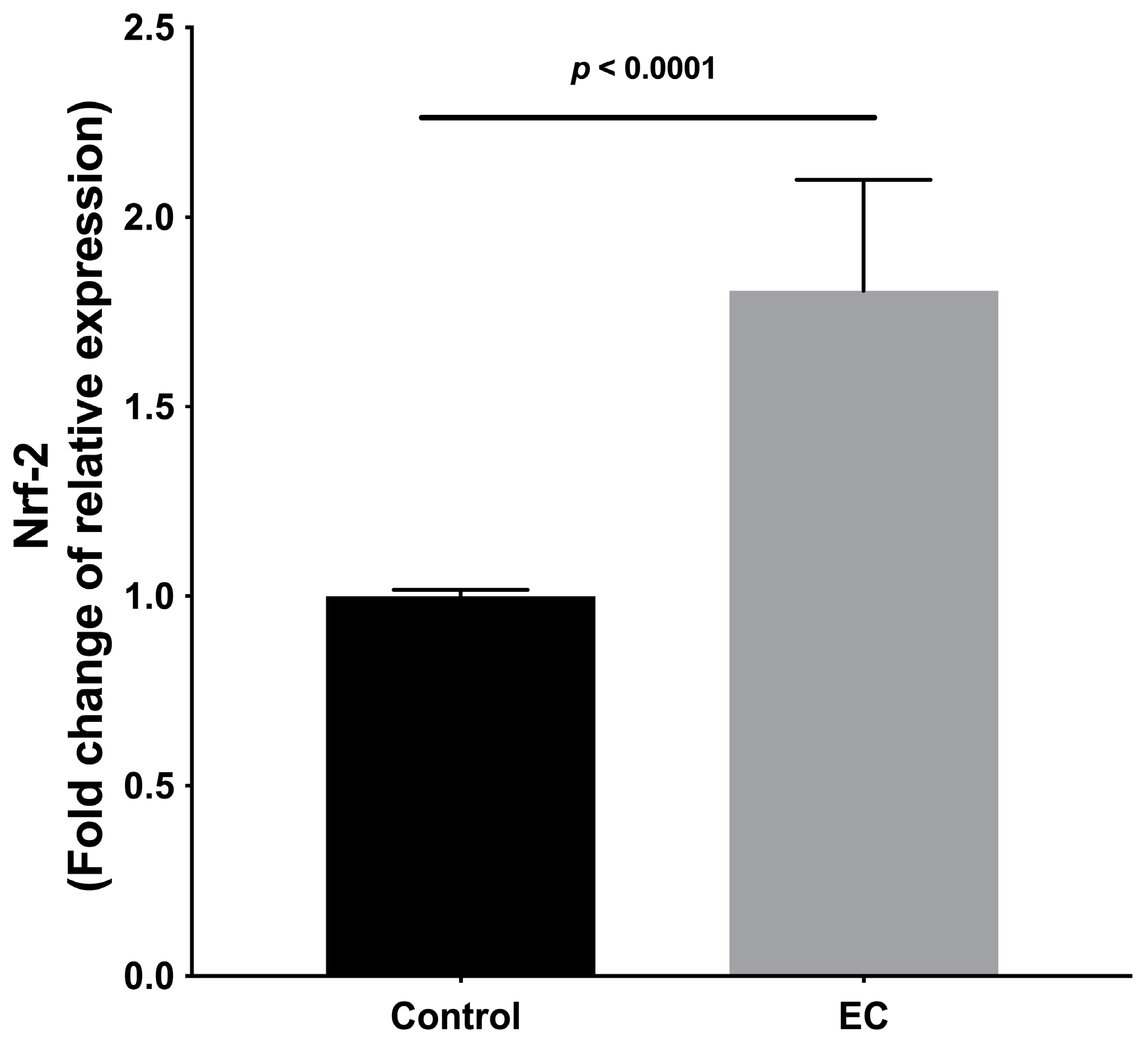
2.5. Expression of Genes Related to the Metastasis Process
3. Discussion
4. Materials and Methods
4.1. 4T1 Cell Culture
4.2. Cytotoxicity Test in 4T1 Cells in Culture
4.3. Cytotoxicity Test in C2C12 Cells in Culture
4.4. Wound Healing
4.5. Cell Invasion Assay
4.6. Analysis of Genes Related to Proliferation and Metastasis by Q-PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Azamjah, N.; Soltan-Zadeh, Y.; Zayeri, F. Global Trend of Breast Cancer Mortality Rate: A 25-Year Study. Asian Pac. J. Cancer Prev. 2019, 20, 2015–2020. [Google Scholar] [CrossRef] [PubMed]
- Ghoncheh, M.; Pournamdar, Z.; Salehiniya, H. Incidence and Mortality and Epidemiology of Breast Cancer in the World. Asian Pac. J. Cancer Prev. 2016, 17, 43–46. [Google Scholar] [CrossRef]
- Merino Bonilla, J.A.; Torres Tabanera, M.; Ros Mendoza, L.H. Breast Cancer in the 21st Century: From Early Detection to New Therapies. Radiología 2017, 59, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Kim, M.Y. Breast Cancer Metastasis. Adv. Exp. Med. Biol. 2021, 1187, 183–204. [Google Scholar] [PubMed]
- Prieto-Callejero, B.; Rivera, F.; Fagundo-Rivera, J.; Romero, A.; Romero-Martín, M.; Gómez-Salgado, J.; Ruiz-Frutos, C. Relationship between Chemotherapy-Induced Adverse Reactions and Health-Related Quality of Life in Patients with Breast Cancer. Medicine 2020, 99, e21695. [Google Scholar] [CrossRef]
- Moreno-Ulloa, A.; Cid, A.; Rubio-Gayosso, I.; Ceballos, G.; Villarreal, F.; Ramirez-Sanchez, I. Effects of (−)-Epicatechin and Derivatives on Nitric Oxide Mediated Induction of Mitochondrial Proteins. Bioorg. Med. Chem. Lett. 2013, 23, 4441–4446. [Google Scholar] [CrossRef]
- Chun, J.H.; Henckel, M.M.; Knaub, L.A.; Hull, S.E.; Pott, G.B.; Walker, L.A.; Reusch, J.E.-B.; Keller, A.C. (−)-Epicatechin Improves Vasoreactivity and Mitochondrial Respiration in Thermoneutral-Housed Wistar Rat Vasculature. Nutrients 2022, 14, 1097. [Google Scholar] [CrossRef]
- Nogueira, L.; Ramirez-Sanchez, I.; Perkins, G.A.; Murphy, A.; Taub, P.R.; Ceballos, G.; Villarreal, F.J.; Hogan, M.C.; Malek, M.H. (−)-Epicatechin Enhances Fatigue Resistance and Oxidative Capacity in Mouse Muscle. J. Physiol. 2011, 589, 4615–4631. [Google Scholar] [CrossRef]
- Dower, J.I.; Geleijnse, J.M.; Gijsbers, L.; Schalkwijk, C.; Kromhout, D.; Hollman, P.C. Supplementation of the Pure Flavonoids Epicatechin and Quercetin Affects Some Biomarkers of Endothelial Dysfunction and Inflammation in (Pre)Hypertensive Adults: A Randomized Double-Blind, Placebo-Controlled, Crossover Trial. J. Nutr. 2015, 145, 1459–1463. [Google Scholar] [CrossRef]
- Gutiérrez-Salmeán, G.; Meaney, E.; Lanaspa, M.A.; Cicerchi, C.; Johnson, R.J.; Dugar, S.; Taub, P.; Ramí-rez-Sánchez, I.; Villarreal, F.; Schreiner, G.; et al. A Randomized, Placebo-Controlled, Double-Blind Study on the Effects of (−)-Epicatechin on the Triglyceride/HDLc Ratio and Cardiometabolic Profile of Subjects with Hypertri-glyceridemia: Unique in Vitro Effects. Int. J. Cardiol. 2016, 223, 500–506. [Google Scholar] [CrossRef]
- McDonald, C.M.; Ramirez-Sanchez, I.; Oskarsson, B.; Joyce, N.; Aguilar, C.; Nicorici, A.; Dayan, J.; Goude, E.; Abresch, R.T.; Villarreal, F.; et al. (−)-Epicatechin Induces Mitochondrial Biogenesis and Markers of Muscle Regeneration in Adults with Becker Muscular Dystrophy. Muscle Nerve 2021, 63, 239–249. [Google Scholar] [CrossRef]
- Almaguer, G.; Ortiz-Vilchis, P.; Cordero, P.; Martinez-Vega, R.; Perez-Durán, J.; Meaney, E.; Villarreal, F.; Ceballos, G.; Nájera, N. Anticancer Potential of (−)-Epicatechin in a Triple-Negative Mammary Gland Model. J. Pharm. Pharmacol. 2021, 73, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Dong, J. (−)-Epicatechin Acts as a Potent Agonist of the Membrane Androgen Receptor, ZIP9 (SLC39A9), to Promote Apoptosis of Breast and Prostate Cancer Cells. J. Steroid Biochem. Mol. Biol. 2021, 211, 105906. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Kuzuhara, T.; Echigo, N.; Suganuma, M.; Fujiki, H. New Role of (−)-Epicatechin in Enhancing the Induction of Growth Inhibition and Apoptosis in Human Lung Cancer Cells by Curcumin. Cancer Prev. Res. 2010, 3, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Sanchez, I.; Maya, L.; Ceballos, G.; Villarreal, F. (−)-Epicatechin Activation of Endothelial Cell Endothelial Nitric Oxide Synthase, Nitric Oxide, and Related Signaling Pathways. Hypertension 2010, 55, 1398–1405. [Google Scholar] [CrossRef]
- Dugar, S.; Villarreal, F.; Hollinger, F.H.; Mahajan, D.; Ramirez-Sanchez, I.; Moreno-Ulloa, A.; Ceballos, G.; Schreiner, G. 11-β-Hydroxysterols as Possible Endogenous Stimulators of Mitochondrial Biogenesis as Inferred from Epicatechin Molecular Mimicry. Pharmacol. Res. 2020, 151, 104540. [Google Scholar] [CrossRef]
- Varela-Castillo, O.; Cordero, P.; Gutiérrez-Iglesias, G.; Palma, I.; Rubio-Gayosso, I.; Meaney, E.; Ramirez-Sanchez, I.; Villarreal, F.; Ceballos, G.; Nájera, N. Characterization of the Cytotoxic Effects of the Combination of Cisplatin and Flavanol (−)-Epicatechin on Human Lung Cancer Cell Line A549. An Isobolographic Approach. Exp. Oncol. 2018, 40, 19–23. [Google Scholar] [CrossRef]
- Utama, K.; Khamto, N.; Meepowpan, P.; Aobchey, P.; Kantapan, J.; Meerak, J.; Roytrakul, S.; Sangthong, P. 2′,4′-Dihydroxy-6′-methoxy-3′,5′-Dimethylchalcone and Its Amino Acid-Conjugated Derivatives Induce G0/G1 Cell Cycle Arrest and Apoptosis via BAX/BCL2 Ratio Upregulation and in Silico Insight in SiHa Cell Lines. Eur. J. Pharm. Sci. 2023, 184, 106390. [Google Scholar] [CrossRef]
- Farhadi, F.; Jahanpour, S.; Hazem, K.; Aghbali, A.; Baradran, B.; Vahid Pakdel, S.M. Garlic (Allium sativum) Fresh Juice Induces Apoptosis in Human Oral Squamous Cell Carcinoma: The Involvement of Caspase-3, Bax and Bcl-2. J. Dent. Res. Dent. Clin. Dent. Prospect. 2015, 9, 267–273. [Google Scholar] [CrossRef]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Orfanos, C.E.; Geilen, C.C.; Wieder, T.; Sturm, I.; Daniel, P.T. The Bax/Bcl-2 Ratio Determines the Susceptibility of Human Melanoma Cells to CD95/Fas-Mediated Apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Sánchez, I.; Rodríguez, A.; Moreno-Ulloa, A.; Ceballos, G.; Villarreal, F. (-)-Epicatechin-Induced Re-covery of Mitochondria from Simulated Diabetes: Potential Role of Endothelial Nitric Oxide Synthase. Diab. Vasc. Dis. Res. 2016, 13, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Daussin, F.N.; Heyman, E.; Burelle, Y. Effects of (−)-Epicatechin on Mitochondria. Nutr. Rev. 2021, 79, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Gong, Y.; Liang, X. Role of FAT1 in Health and Disease (Review). Oncol. Lett. 2021, 21, 398. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Shen, W.; Gao, B.; Xu, J.; Gong, B. Metastasis Suppressor 1 Acts as a Tumor Suppressor by Inhibiting Epithelial-to-Mesenchymal Transition in Triple-Negative Breast Cancer. Int. J. Biol. Markers 2020, 35, 74–81. [Google Scholar] [CrossRef]
- Dawson, J.C.; Bruche, S.; Spence, H.J.; Braga, V.M.M.; Machesky, L.M. Mtss1 Promotes Cell-Cell Junction As-sembly and Stability through the Small GTPase Rac1. PLoS ONE 2012, 7, e31141. [Google Scholar] [CrossRef]
- Chen, J.; Sun, J.; Wang, Q.; Du, Y.; Cheng, J.; Yi, J.; Xie, B.; Jin, S.; Chen, G.; Wang, L.; et al. Systemic Deficiency of PTEN Accelerates Breast Cancer Growth and Metastasis. Front. Oncol. 2022, 12, 825484. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Chen, C.-Z.; Jin, L.-P.; Ji, Q.-Q.; Chen, Y.-Z.; Li, Q.; Zhang, X.-H.; Qu, J.-M. Effect and Mechanism of the Metastasis Suppressor Gene BRMS1 on the Migration of Breast Cancer Cells. Int. J. Clin. Exp. Med. 2013, 6, 908–916. [Google Scholar]
- Cao, Y.; Tan, S.; Tu, Y.; Zhang, G.; Liu, Y.; Li, D.; Xu, S.; Le, Z.; Xiong, J.; Zou, W.; et al. MicroRNA-125a-5p Inhibits Invasion and Metastasis of Gastric Cancer Cells by Targeting BRMS1 Expression. Oncol. Lett. 2018, 15, 5119–5130. [Google Scholar] [CrossRef]
- Phadke, P.A.; Vaidya, K.S.; Nash, K.T.; Hurst, D.R.; Welch, D.R. BRMS1 Suppresses Breast Cancer Experimental Metastasis to Multiple Organs by Inhibiting Several Steps of the Metastatic Process. Am. J. Pathol. 2008, 172, 809–817. [Google Scholar] [CrossRef]
- Sporn, M.B.; Liby, K.T. NRF2 and Cancer: The Good, the Bad and the Importance of Context. Nat. Rev. Cancer 2012, 12, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gordon, G.B. A Strategy for Cancer Prevention: Stimulation of the Nrf2-ARE Signaling Pathway. Mol. Cancer Ther. 2004, 3, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kumar, R.M.; Bhattacharjee, D.; Somanna, P.; Jain, V. Role of Nrf2 Signaling Cascade in Breast Cancer: Strategies and Treatment. Front. Pharmacol. 2022, 13, 720076. [Google Scholar] [CrossRef]
- Kim, W.D.; Kim, Y.W.; Cho, I.J.; Lee, C.H.; Kim, S.G. E-Cadherin Inhibits Nuclear Accumulation of Nrf2: Im-plications for Chemoresistance of Cancer Cells. J. Cell Sci. 2012, 125, 1284–1295. [Google Scholar] [CrossRef]
- Lee, S.B.; Sellers, B.N.; DeNicola, G.M. The Regulation of NRF2 by Nutrient-Responsive Signaling and Its Role in Anabolic Cancer Metabolism. Antioxid. Redox Signal. 2018, 29, 1774–1791. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Baird, L.; Zhang, Y.; Hargreaves, I.; Chalasani, A.; Land, J.M.; Stanyer, L.; Yamamoto, M.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 Impacts Cellular Bioenergetics by Controlling Substrate Availability for Mitochondrial Respiration. Biol. Open 2013, 2, 761–770. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Brose, R.D.; Shin, G.; McGuinness, M.C.; Schneidereith, T.; Purvis, S.; Dong, G.X.; Keefer, J.; Spencer, F.; Smith, K.D. Activation of the Stress Proteome as a Mechanism for Small Molecule Therapeutics. Hum. Mol. Genet. 2012, 21, 4237–4252. [Google Scholar] [CrossRef]
- Chidambaram, S.B.; Bhat, A.; Ray, B.; Sugumar, M.; Muthukumar, S.P.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M.; Guillemin, G.J.; Sakharkar, M.K. Cocoa Beans Improve Mitochondrial Biogenesis via PPARγ/PGC1α Dependent Signalling Pathway in MPP + Intoxicated Human Neuroblastoma Cells (SH-SY5Y). Nutr. Neurosci. 2020, 23, 471–480. [Google Scholar] [CrossRef]
- He, Q.; Yang, J.; Pan, Z.; Zhang, G.; Chen, B.; Li, S.; Xiao, J.; Tan, F.; Wang, Z.; Chen, P.; et al. Biochanin A Protects against Iron Overload Associated Knee Osteoarthritis via Regulating Iron Levels and NRF2/System Xc-/GPX4 Axis. Biomed. Pharmacother. 2023, 157, 113915. [Google Scholar] [CrossRef]
- Zhou, R.; Xu, L.; Ye, M.; Liao, M.; Du, H.; Chen, H. Formononetin Inhibits Migration and Invasion of MDA-MB-231 and 4T1 Breast Cancer Cells by Suppressing MMP-2 and MMP-9 through PI3K/AKT Signaling Pathways. Horm. Metab. Res. 2014, 46, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhou, W.; Zhang, X.; Wang, Y.; Yang, D.; Li, S. Protective Effect of Blood Cora Polysaccharides on H9c2 Rat Heart Cells Injury Induced by Oxidative Stress by Activating Nrf2/HO-1 Signal Pathway. Front. Nutr. 2021, 8, 632161. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′ to 3′) | Reverse Primer (5′to 3′) | Reference |
|---|---|---|---|
| Bax | AGGATGCGTCCACCAAGAAGCT | TCCGTGTCCACGTCAGCAATCA | [40] |
| Bcl2 | CCTGTGGATGACTGAGTACCTG | AGCCAGGAGAAATCAAACAGAGG | [40] |
| Mmp9 | TGTTCCCGTTCATCTTTGAG | ATCCTGGTCATAGTTGGCTGT | [41] |
| Cdh1 | CTCCAGTCATAGGGAGCTGTC | TCTTCTGAGACCTGGGTACAC | ID: 118129809c3 |
| Mtss1 | ATGGAGGCTGTGATCGAGAAG | CCAAACTGGATAGCTCCCCT | ID: 226052051c1 |
| Pten | TGGATTCGACTTAGACTTGACCT | GCGGTGTCATAATGTCTCTCAG | ID: 6679523a1 |
| Brms | GGTGGACTACGCGGAGAAC | CACCTGACTCAACCGCTCTTT | ID: 27754014a1 |
| Fat1 | CTACGGAGGAACGTGCATGG | ATCTTTGCAGTACGGACTAAGC | ID: 27697111a1 |
| Smad4 | ACACCAACAAGTAACGATGCC | GCAAAGGTTTCACTTTCCCCA | ID: 28201436a1 |
| Nrf2 | TAGATGACCATGAGTCGCTTGC | GCCAAACTTGCTCCATGTCC | [42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Durán, J.; Luna, A.; Portilla, A.; Martínez, P.; Ceballos, G.; Ortíz-Flores, M.Á.; Solis-Paredes, J.M.; Nájera, N. (−)-Epicatechin Inhibits Metastatic-Associated Proliferation, Migration, and Invasion of Murine Breast Cancer Cells In Vitro. Molecules 2023, 28, 6229. https://doi.org/10.3390/molecules28176229
Pérez-Durán J, Luna A, Portilla A, Martínez P, Ceballos G, Ortíz-Flores MÁ, Solis-Paredes JM, Nájera N. (−)-Epicatechin Inhibits Metastatic-Associated Proliferation, Migration, and Invasion of Murine Breast Cancer Cells In Vitro. Molecules. 2023; 28(17):6229. https://doi.org/10.3390/molecules28176229
Chicago/Turabian StylePérez-Durán, Javier, Aglaé Luna, Andrés Portilla, Pamela Martínez, Guillermo Ceballos, Miguel Ángel Ortíz-Flores, Juan Mario Solis-Paredes, and Nayelli Nájera. 2023. "(−)-Epicatechin Inhibits Metastatic-Associated Proliferation, Migration, and Invasion of Murine Breast Cancer Cells In Vitro" Molecules 28, no. 17: 6229. https://doi.org/10.3390/molecules28176229
APA StylePérez-Durán, J., Luna, A., Portilla, A., Martínez, P., Ceballos, G., Ortíz-Flores, M. Á., Solis-Paredes, J. M., & Nájera, N. (2023). (−)-Epicatechin Inhibits Metastatic-Associated Proliferation, Migration, and Invasion of Murine Breast Cancer Cells In Vitro. Molecules, 28(17), 6229. https://doi.org/10.3390/molecules28176229