
Walnut Kernel Oil and Defatted Extracts Enhance Mesenchymal Stem Cell Stemness and Delay Senescence
 , , ,
, , ,  , , ,
, , , 
Abstract
:1. Introduction
2. Results
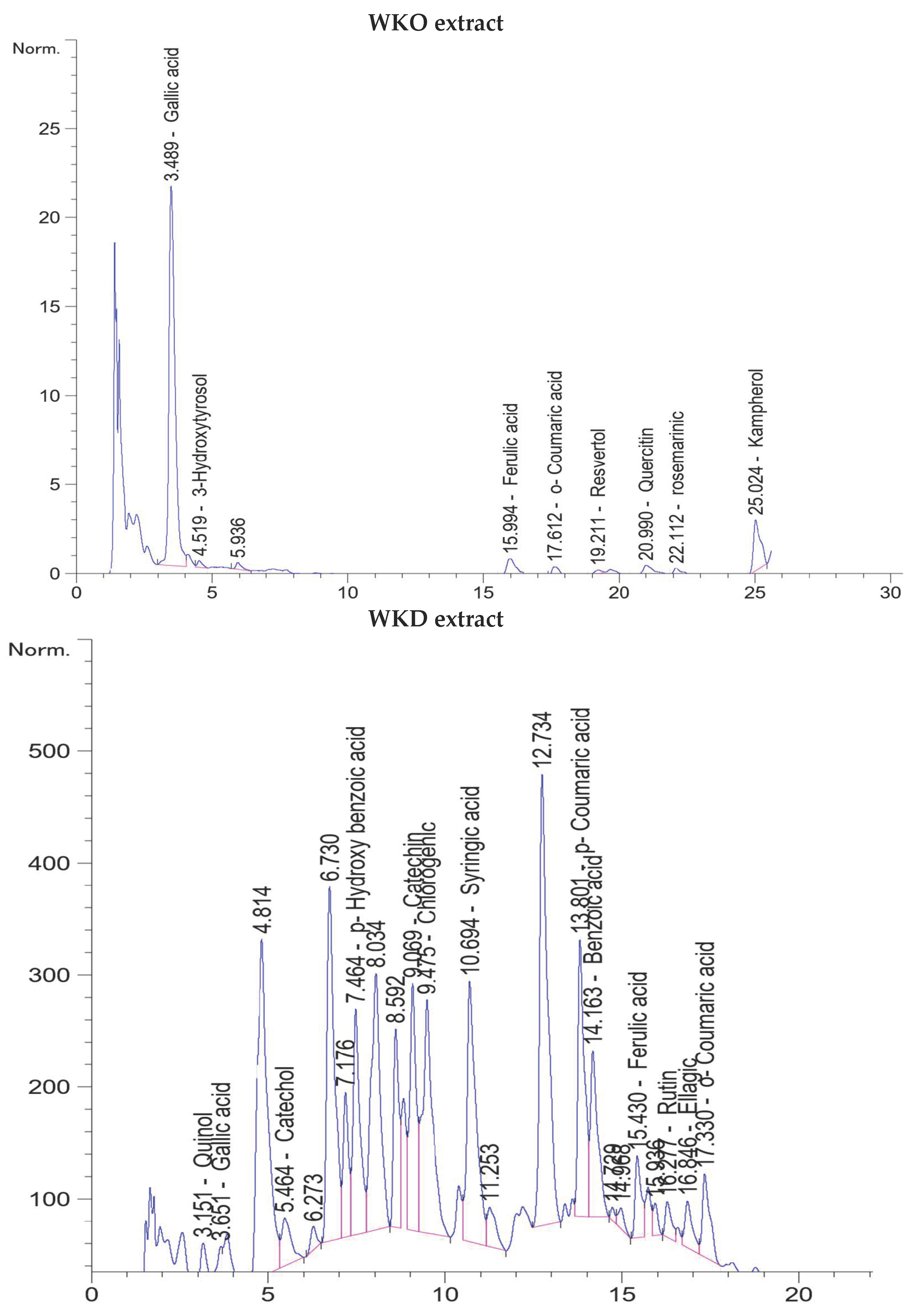
2.1. Flavonoids and Phenolic Compounds of WKO and WKD Extracts as Detected by HPLC
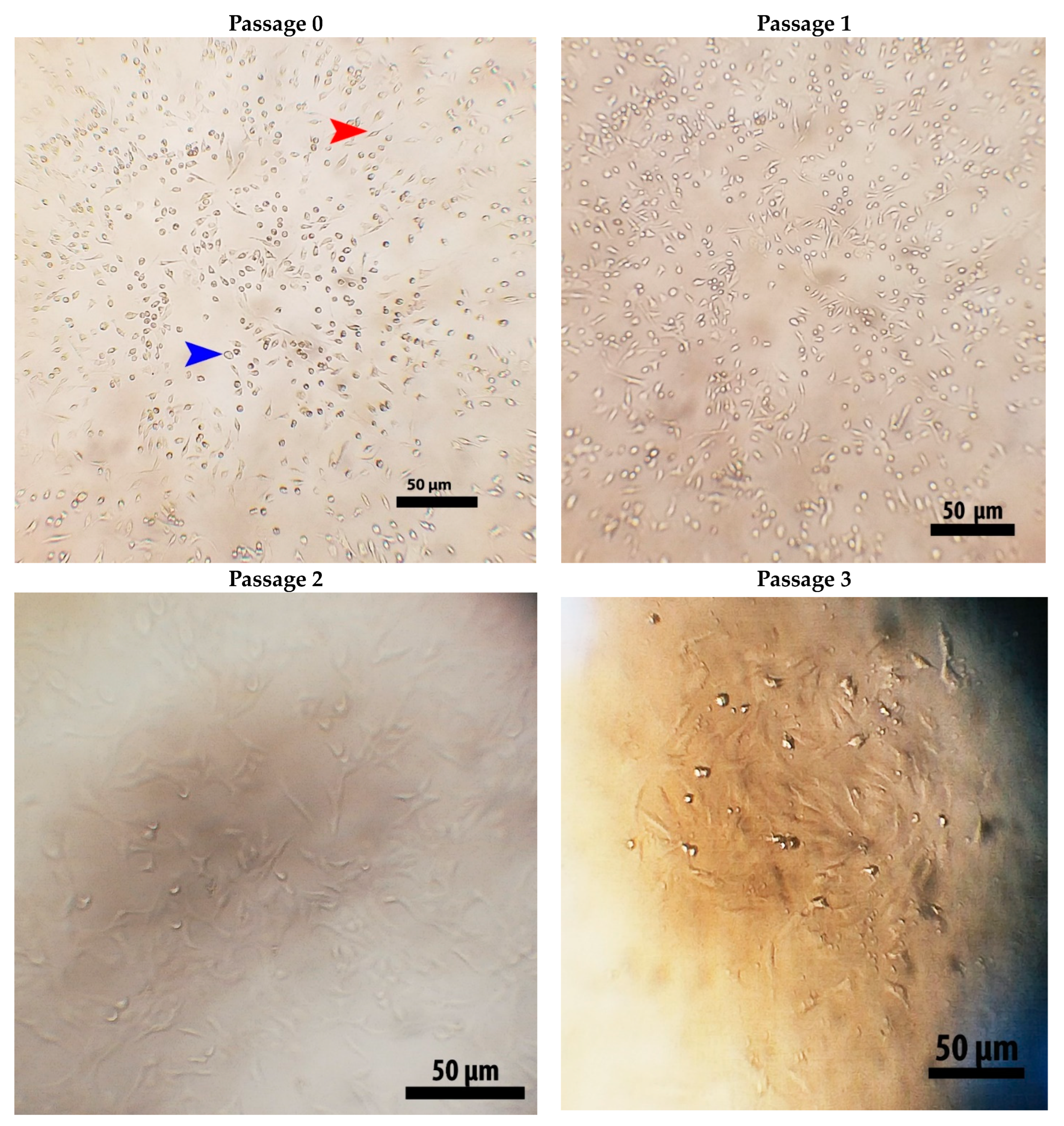
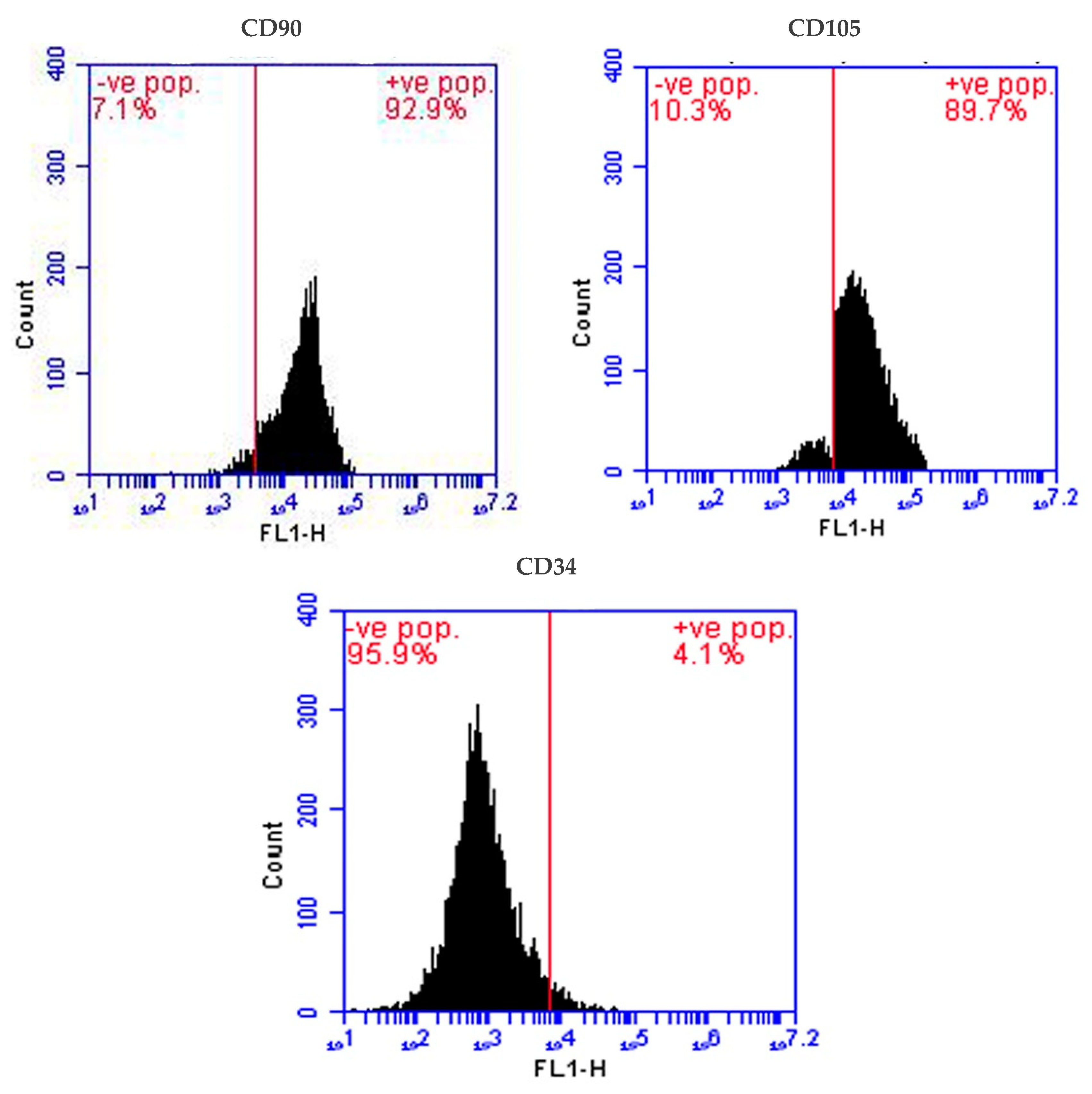
2.2. Isolation and Identification of Stem Cells
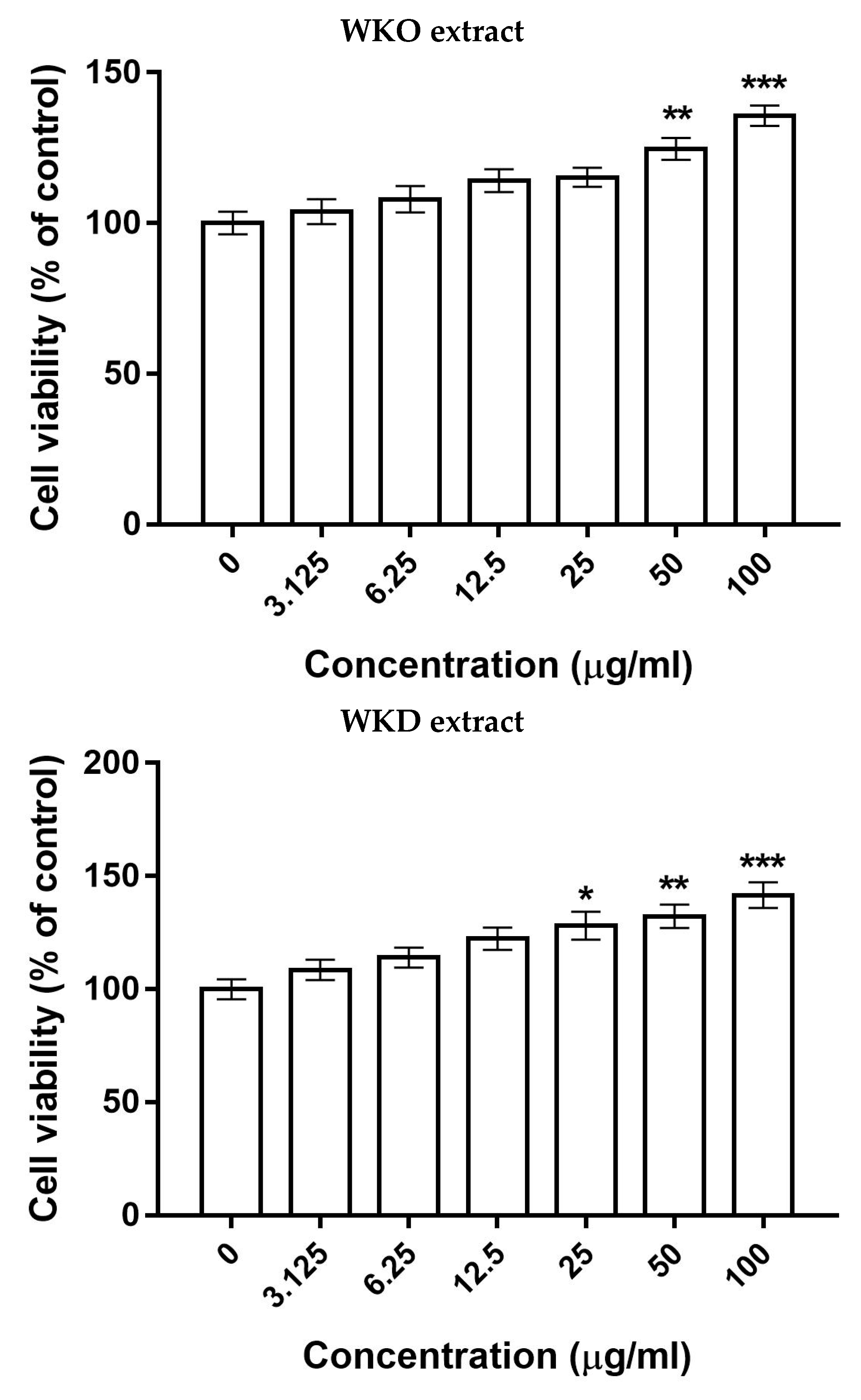
2.3. WKO and WKD Extracts Improved BM-MSC Stemness
2.3.1. WKO and WKD Extracts Enhanced BM-MSC Proliferation
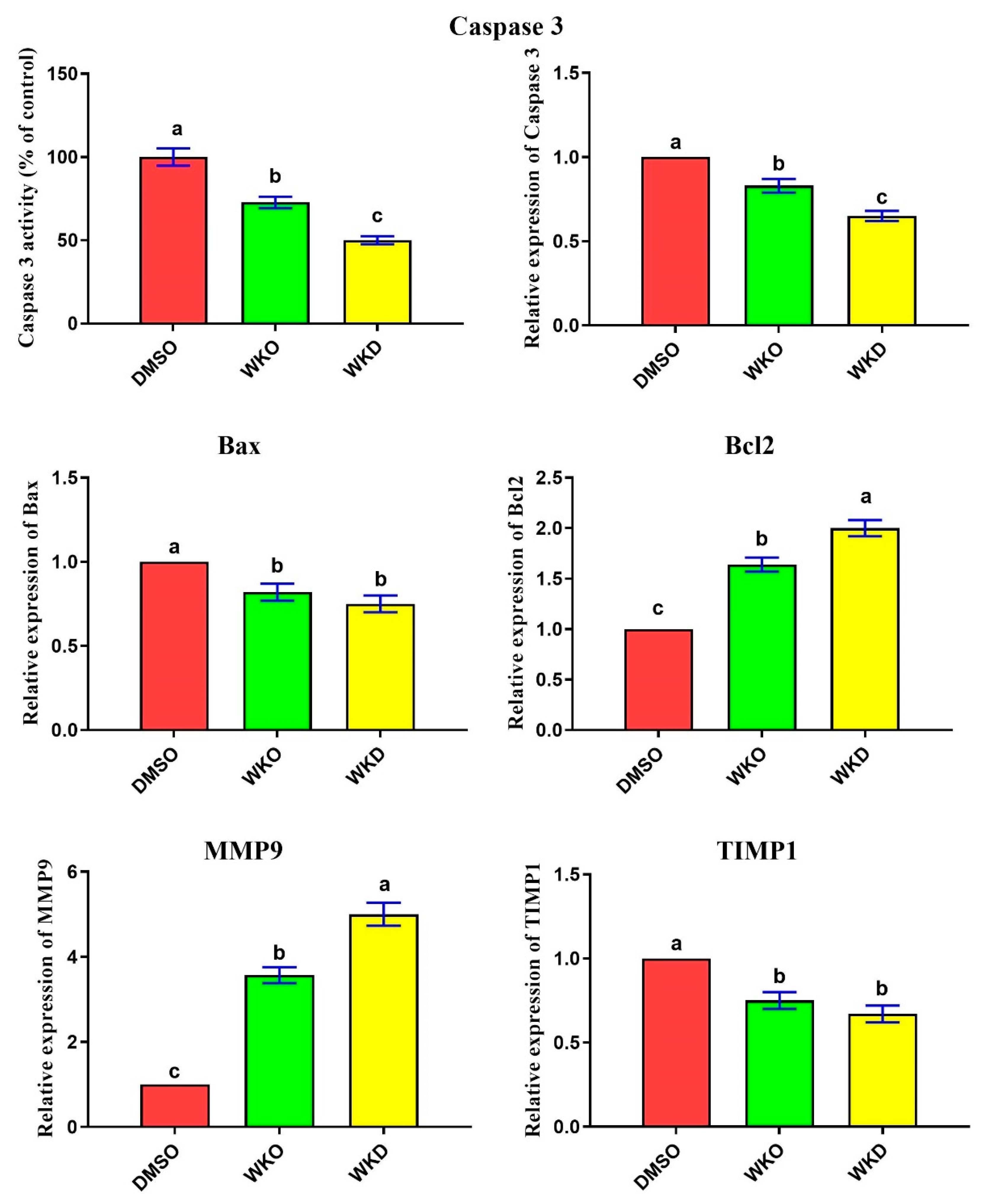
2.3.2. WKO and WKD Extracts Inhibited Apoptosis and Induced Migration of BM-MSCs
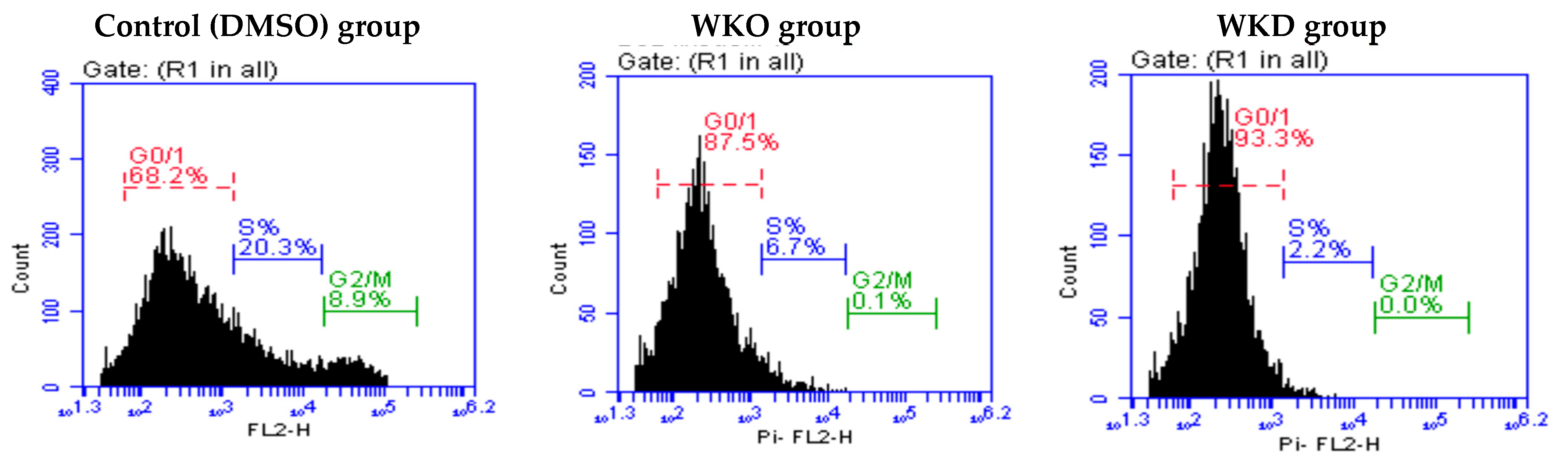
2.3.3. WKO and WKD Extracts Affected BM-MSC Cell Cycle
2.4. WKO and WKD Extracts Reduced BM-MSC H2O2-Induced Premature Senescence
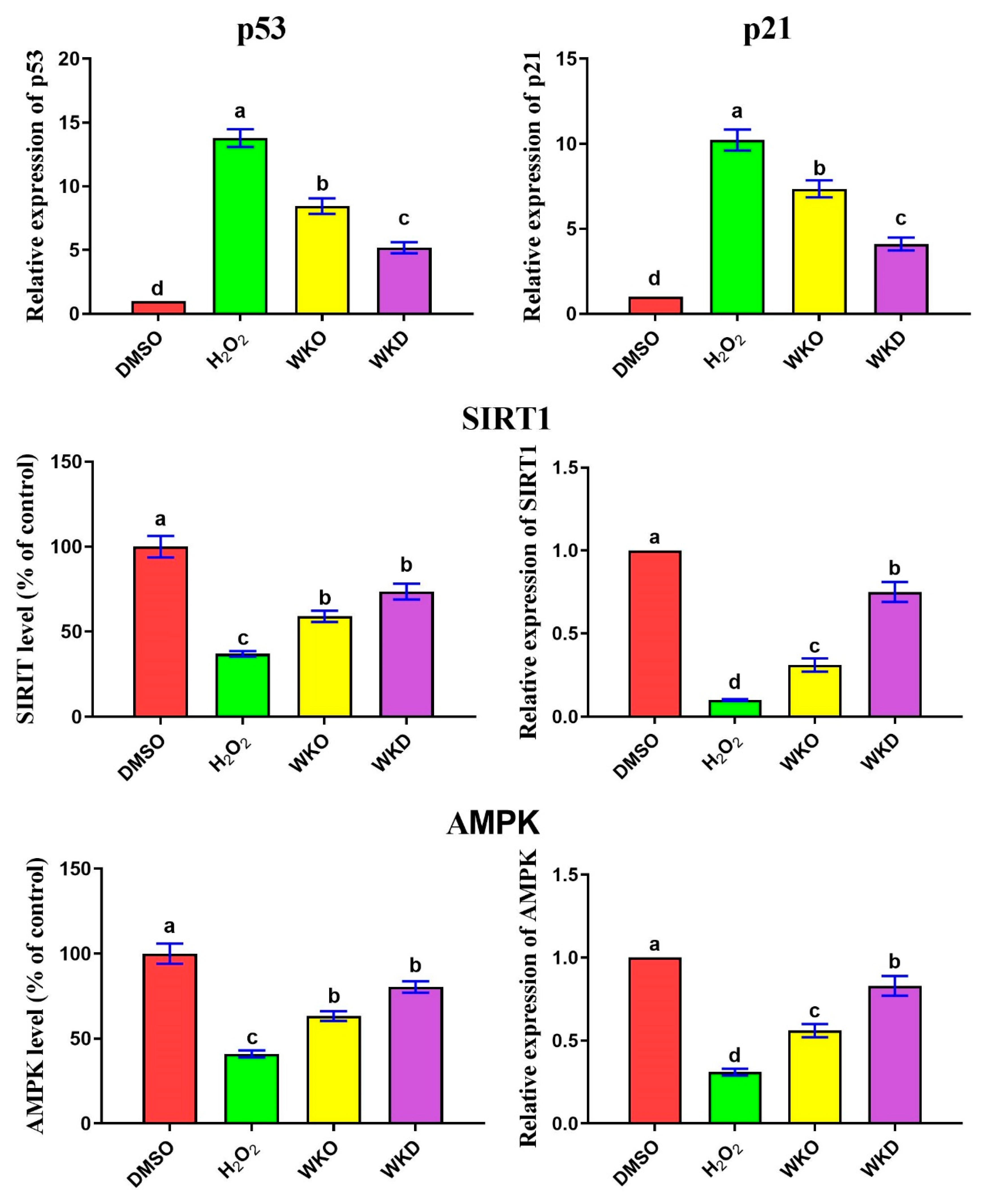
2.4.1. WKO and WKD Extracts Repressed BM-MSC Senescence-Related Markers
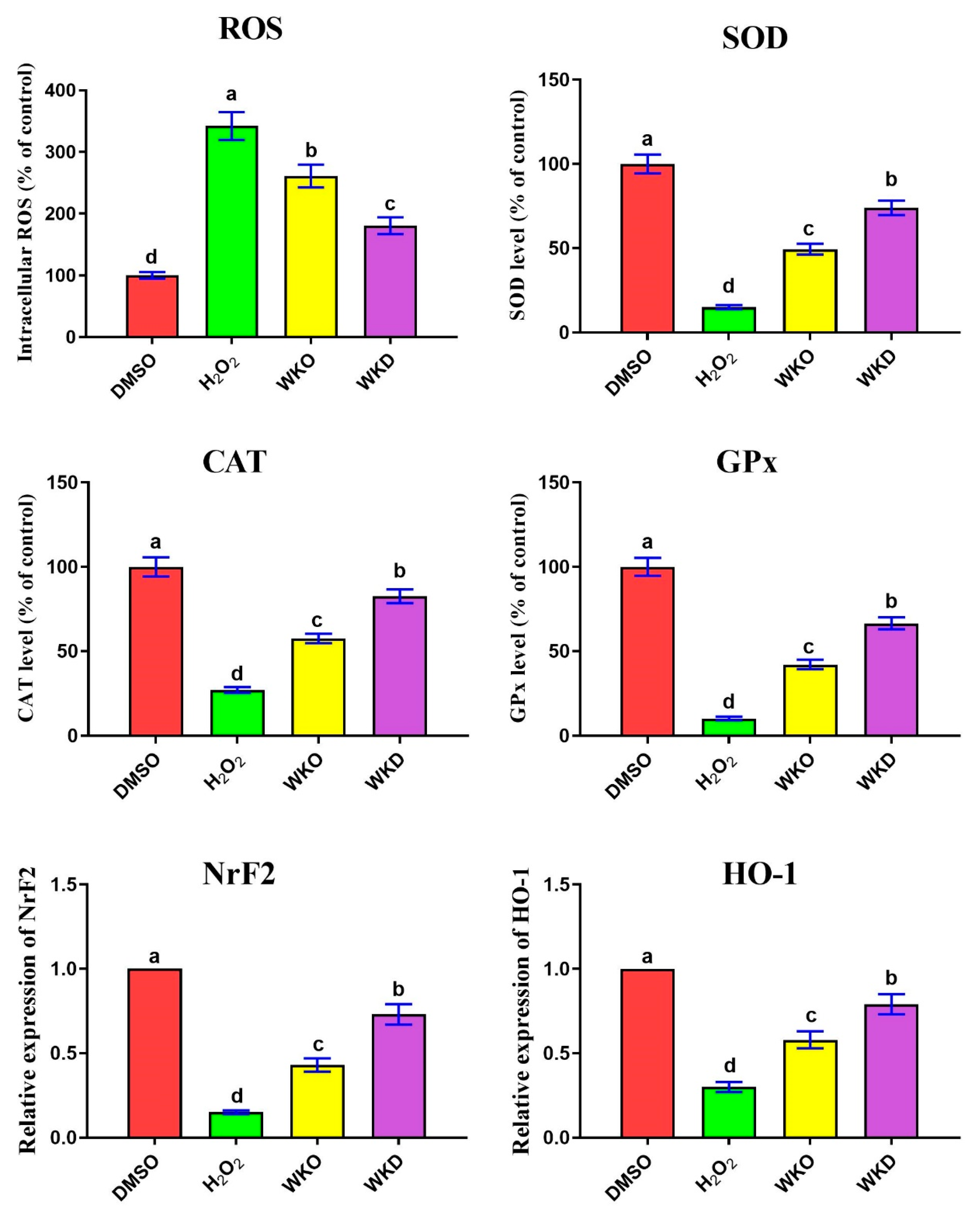
2.4.2. WKO and WKD Extracts Protected BM-MSCs against H2O2-Triggered ROS
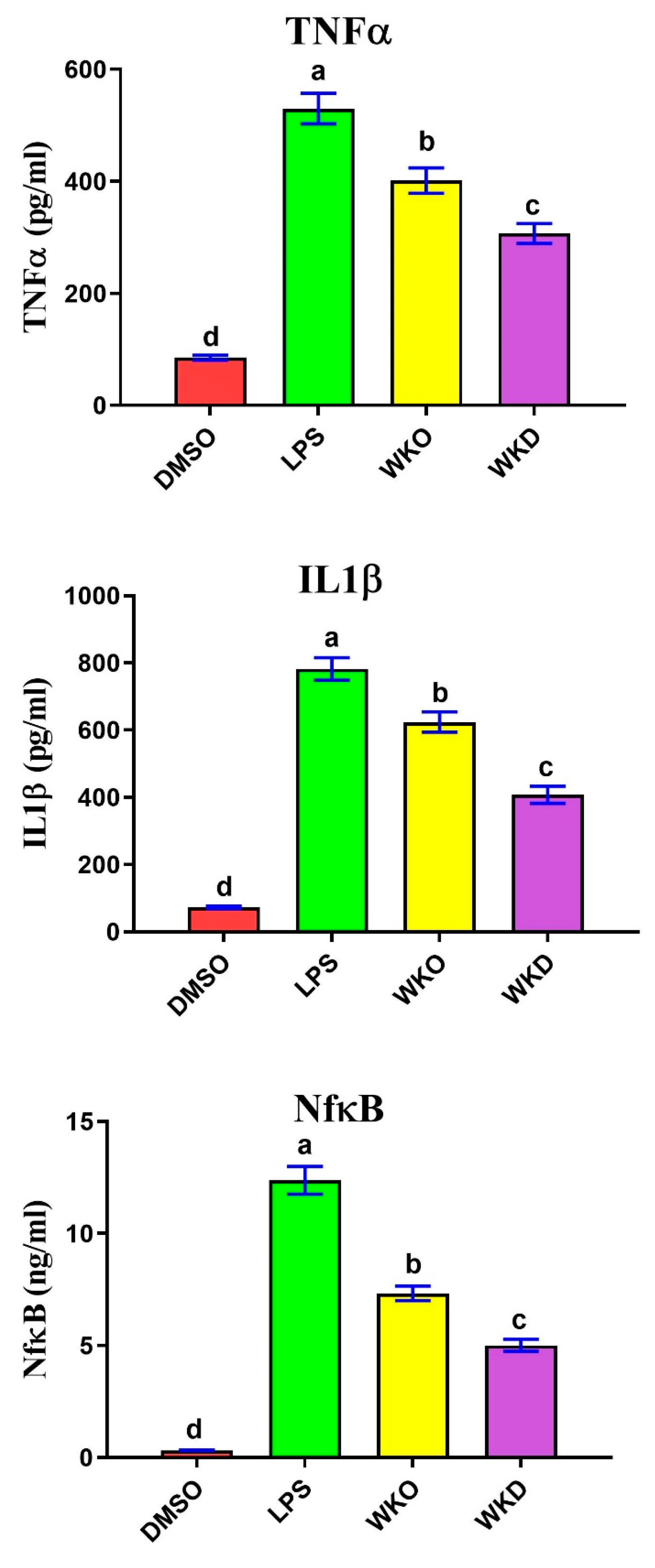
2.4.3. WKO and WKD Extracts Protected BM-MSCs against LPS-Induced Inflammation
3. Discussion
4. Materials and Methods
4.1. Preparation of Extract
4.2. Detection of Flavonoids and Phenolic Compounds by HPLC
4.3. Isolation and Identification of Stem Cells
4.4. MTT-Based Cell Proliferation Assay
4.5. Experimental Approach
4.6. Detection of Gene Expression by qPCR
4.7. Cell Cycle by Flow Cytometry
4.8. Intracellular Reactive Oxygen Species
4.9. Determination of Antioxidant Enzyme Activity
4.10. ELISA
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Daley, G.Q. Stem cells and the evolving notion of cellular identity. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20140376. [Google Scholar] [CrossRef] [PubMed]
- Abd-Allah, S.H.; Shalaby, S.M.; Abd-Elbary, E.; Saleh, A.A.; El-Magd, M.A. Human peripheral blood CD34+ cells attenuate oleic acid-induced acute lung injury in rats. Cytotherapy 2015, 17, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Biehl, J.K.; Russell, B. Introduction to stem cell therapy. J. Cardiovasc. Nurs. 2009, 24, 98–105. [Google Scholar] [CrossRef]
- Ewald, J.A.; Desotelle, J.A.; Wilding, G.; Jarrard, D.F. Therapy-induced senescence in cancer. JNCI J. Natl. Cancer Inst. 2010, 102, 1536–1546. [Google Scholar] [CrossRef]
- He, S.; Zhou, M.; Zheng, H.; Wang, Y.; Wu, S.; Gao, Y.; Chen, J. Resveratrol inhibits the progression of premature senescence partially by regulating v-rel avian reticuloendotheliosis viral oncogene homolog A (RELA) and sirtuin 1 (SIRT1). Ren. Fail. 2022, 44, 171–183. [Google Scholar] [CrossRef]
- Jones, K.; Elmore, L.; Jackson-Cook, C.; Demasters, G.; Povirk, L.; Holt, S.; Gewirtz, D. p53-Dependent accelerated senescence induced by ionizing radiation in breast tumour cells. Int. J. Radiat. Biol. 2005, 81, 445–458. [Google Scholar] [CrossRef]
- Al-Azab, M.; Idiiatullina, E.; Safi, M.; Hezam, K. Enhancers of mesenchymal stem cell stemness and therapeutic potency. Biomed. Pharmacother. Biomed. Pharmacother. 2023, 162, 114356. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Soflou, R.; Mohseni-Vadeghani, E.; Karkhaneh, A. Controlled release of resveratrol from a composite nanofibrous scaffold: Effect of resveratrol on antioxidant activity and osteogenic differentiation. J. Biomed. Mater. Res. Part A 2022, 110, 21–30. [Google Scholar] [CrossRef]
- Abed, É.; Reboul, P.; Lajeunesse, D. Sirtuins as Markers of Bone Disease: A Focus on Osteoarthritis and Osteoporosis. In Biomarkers in Bone Disease; Preedy, V.R., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 1–19. [Google Scholar]
- Ali, D.; Chen, L.; Kowal, J.M.; Okla, M.; Manikandan, M.; AlShehri, M.; AlMana, Y.; AlObaidan, R.; AlOtaibi, N.; Hamam, R. Resveratrol inhibits adipocyte differentiation and cellular senescence of human bone marrow stromal stem cells. Bone 2020, 133, 115252. [Google Scholar] [CrossRef]
- Chen, T.S.; Chuang, S.Y.; Shen, C.Y.; Ho, T.J.; Chang, R.L.; Yeh, Y.L.; Kuo, C.H.; Mahalakshmi, B.; Kuo, W.W.; Huang, C.Y. Antioxidant Sirt1/Akt axis expression in resveratrol pretreated adipose-derived stem cells increases regenerative capability in a rat model with cardiomyopathy induced by diabetes mellitus. J. Cell. Physiol. 2021, 236, 4290–4302. [Google Scholar] [CrossRef]
- Khanh, V.C.; Zulkifli, A.F.; Tokunaga, C.; Yamashita, T.; Hiramatsu, Y.; Ohneda, O. Aging impairs beige adipocyte differentiation of mesenchymal stem cells via the reduced expression of Sirtuin 1. Biochem. Biophys. Res. Commun. 2018, 500, 682–690. [Google Scholar] [CrossRef]
- Qin, W.; Gao, X.; Ma, T.; Weir, M.D.; Zou, J.; Song, B.; Lin, Z.; Schneider, A.; Xu, H.H. Metformin enhances the differentiation of dental pulp cells into odontoblasts by activating AMPK signaling. J. Endod. 2018, 44, 576–584. [Google Scholar] [CrossRef]
- Uğur, Y.; Erdoğan, S. FATTY ACİDS İN WALNUTS. 2021, p. 339. Available online: https://www.researchgate.net/publication/357152430_FATTY_ACIDS_IN_WALNUTS (accessed on 25 February 2023).
- Trandafir, I.; Cosmulescu, S.; Botu, M.; Nour, V. Antioxidant activity, and phenolic and mineral contents of the walnut kernel (Juglans regia L.) as a function of the pellicle color. Fruits 2016, 71, 177–184. [Google Scholar] [CrossRef]
- Jahanban-Esfahlan, A.; Davaran, S.; Moosavi-Movahedi, A.A.; Dastmalchi, S. Investigating the interaction of juglone (5-hydroxy-1, 4-naphthoquinone) with serum albumins using spectroscopic and in silico methods. J. Iran. Chem. Soc. 2017, 14, 1527–1540. [Google Scholar] [CrossRef]
- Martínez, M.L.; Labuckas, D.O.; Lamarque, A.L.; Maestri, D.M. Walnut (Juglans regia L.): Genetic resources, chemistry, by-products. J. Sci. Food Agric. 2010, 90, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.S.; Hussain, A.; Ganie, S.; Munshi, A.; Lal, E.; Gupta, R. Juglans regia—A review. Int. J. Latest Res. Sci. Technol 2016, 5, 90–97. [Google Scholar]
- Chung, J.; Kim, Y.S.; Lee, J.; Lee, J.H.; Choi, S.W.; Kim, Y. Compositional analysis of walnut lipid extracts and properties as an anti-cancer stem cell regulator via suppression of the self-renewal capacity. Food Sci. Biotechnol. 2016, 25, 623–629. [Google Scholar] [CrossRef]
- Carvalho, M.; Ferreira, P.J.; Mendes, V.S.; Silva, R.; Pereira, J.A.; Jerónimo, C.; Silva, B.M. Human cancer cell antiproliferative and antioxidant activities of Juglans regia L. Food Chem. Toxicol. 2010, 48, 441–447. [Google Scholar] [CrossRef]
- Shi, D.; Chen, C.; Zhao, S.; Ge, F.; Liu, D.; Song, H. Walnut polyphenols inhibit pancreatic lipase activity in vitro and have hypolipidemic effect on high-fat diet-induced obese Mice. J. Food Nutr. Res. 2014, 2, 757–763. [Google Scholar] [CrossRef]
- Esfahlan, A.J.; Jamei, R.; Esfahlan, R.J. The importance of almond (Prunus amygdalus L.) and its by-products. Food Chem. 2010, 120, 349–360. [Google Scholar] [CrossRef]
- Meshkini, A.; Tahmasbi, M. Antiplatelet aggregation activity of walnut hull extract via suppression of reactive oxygen species generation and caspase activation. J. Acupunct. Meridian Stud. 2017, 10, 193–203. [Google Scholar] [CrossRef]
- Salejda, A.M.; Janiewicz, U.; Korzeniowska, M.; Kolniak-Ostek, J.; Krasnowska, G. Effect of walnut green husk addition on some quality properties of cooked sausages. LWT-Food Sci. Technol. 2016, 65, 751–757. [Google Scholar] [CrossRef]
- Li, C.-Y.; Du, H.-J.; Su, X.-H.; Zhong, Y.-J.; Yuan, Z.-P.; Li, Y.-F.; Liang, B. Juglanones A and B: Two Novel Tetralone Dimers from Walnut Pericarp (Juglans regia). Helv. Chim. Acta 2013, 96, 1031–1035. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, B.; Jiang, Y.; Liu, Z.; Liu, Y.; Wang, X.; Kuang, H. Studies on Cytotoxic Activity against HepG-2 Cells of Naphthoquinones from Green Walnut Husks of Juglans mandshurica Maxim. Molecules 2015, 20, 15572–15588. [Google Scholar] [CrossRef] [PubMed]
- Tsasi, G.; Samara, P.; Tsitsilonis, O.; Jürgenliemk, G.; Skaltsa, H. Isolation, Identification and Cytotoxic Activity of Triterpenes and Flavonoids from Green Walnut (Juglans regia L.) Pericarps. Rec. Nat. Prod. 2015, 10, 83–92. [Google Scholar]
- Rusu, M.E.; Fizesan, I.; Pop, A.; Mocan, A.; Gheldiu, A.M.; Babota, M.; Vodnar, D.C.; Jurj, A.; Berindan-Neagoe, I.; Vlase, L.; et al. Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects. Molecules 2020, 25, 2187. [Google Scholar] [CrossRef] [PubMed]
- Aghapour SK, F.; Sisakhtnezhad, S. Effect of the Internal Septum Extract of the Walnut Kernel on the Mesenchymal Stem Cells Cycle and MSCs-derived Insulin-Producing Β-cells Differentiation and Glucose Uptake. Jentashapir. J. Cell. Mol. Biol. 2021, 12, e115014. [Google Scholar] [CrossRef]
- Lee, J.; Kim, Y.S.; Lee, J.; Heo, S.C.; Lee, K.L.; Choi, S.W.; Kim, Y. Walnut Phenolic Extract and Its Bioactive Compounds Suppress Colon Cancer Cell Growth by Regulating Colon Cancer Stemness. Nutrients 2016, 8, 439. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; El-Magd, M.A.; Abdelfattah-Hassan, A.; Saleh, A.A.; Saadeldin, I.M.; El-Shetry, E.S.; Badawy, A.A.; Alkarim, S. Potential Effect of Exosomes Derived from Cancer Stem Cells and MSCs on Progression of DEN-Induced HCC in Rats. Stem Cells Int. 2018, 2018, 8058979. [Google Scholar] [CrossRef]
- Hafeman, D.G.; Sunde, R.A.; Hoekstra, W.G. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutr. 1974, 104, 580–587. [Google Scholar] [CrossRef]
- Soleimani, M.; Nadri, S. A protocol for isolation and culture of mesenchymal stem cells from mouse bone marrow. Nat. Protoc. 2009, 4, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.L.; Yu, X.F.; Qu, S.C.; Qu, X.R.; Jiang, Y.F. Juglone, from Juglans mandshruica Maxim, inhibits growth and induces apoptosis in human leukemia cell HL-60 through a reactive oxygen species-dependent mechanism. Food Chem. Toxicol. 2012, 50, 590–596. [Google Scholar] [CrossRef]
- Zahran, R.; Ghozy, A.; Elkholy, S.S.; El-Taweel, F.; El-Magd, M.A. Combination therapy with melatonin, stem cells and extracellular vesicles is effective in limiting renal ischemia–reperfusion injury in a rat model. Int. J. Urol. 2020, 27, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Y.; Basyony, M.A.; El-Desouki, N.I.; Abdo, W.S.; El-Magd, M.A. The potential therapeutic effect for melatonin and mesenchymal stem cells on hepatocellular carcinoma. BioMedicine 2019, 9, 23–29. [Google Scholar] [CrossRef]
- Chen, X.; Lu, Y.; Zhang, Z.; Wang, J.; Yang, H.; Liu, G. Intercellular interplay between Sirt1 signalling and cell metabolism in immune cell biology. Immunology 2015, 145, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Basu-Roy, U.; Zavadil, J.; Basilico, C.; Mansukhani, A. Distinct functions of Sox2 control self-renewal and differentiation in the osteoblast lineage. Mol. Cell. Biol. 2011, 31, 4593–4608. [Google Scholar] [CrossRef] [PubMed]
- Simic, P.; Zainabadi, K.; Bell, E.; Sykes, D.B.; Saez, B.; Lotinun, S.; Baron, R.; Scadden, D.; Schipani, E.; Guarente, L. SIRT1 regulates differentiation of mesenchymal stem cells by deacetylating β-catenin. EMBO Mol. Med. 2013, 5, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.R.; Perrien, D.S.; Fleming, N.; Nyman, J.S.; Ono, K.; Connelly, L.; Moore, M.M.; Lwin, S.T.; Yull, F.E.; Mundy, G.R.; et al. Silent information regulator (Sir)T1 inhibits NF-κB signaling to maintain normal skeletal remodeling. J. Bone Miner. Res. 2013, 28, 960–969. [Google Scholar] [CrossRef]
- Herranz, D.; Muñoz-Martin, M.; Cañamero, M.; Mulero, F.; Martinez-Pastor, B.; Fernandez-Capetillo, O.; Serrano, M. Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat. Commun. 2010, 1, 3. [Google Scholar] [CrossRef]
- Rusu, M.E.; Georgiu, C.; Pop, A.; Mocan, A.; Kiss, B.; Vostinaru, O.; Fizesan, I.; Stefan, M.G.; Gheldiu, A.M.; Mates, L.; et al. Antioxidant Effects of Walnut (Juglans regia L.) Kernel and Walnut Septum Extract in a D-Galactose-Induced Aging Model and in Naturally Aged Rats. Antioxidants 2020, 9, 424. [Google Scholar] [CrossRef]
- El-Magd, M.A.; Mohamed, Y.; El-Shetry, E.S.; Elsayed, S.A.; Abo Gazia, M.; Abdel-Aleem, G.A.; Shafik, N.M.; Abdo, W.S.; El-Desouki, N.I.; Basyony, M.A. Melatonin maximizes the therapeutic potential of non-preconditioned MSCs in a DEN-induced rat model of HCC. Biomed. Pharmacother. 2019, 114, 108732. [Google Scholar] [CrossRef]
- Saad, E.; Marei, H.; El-Magd, M.; El-Fatiry, H. Molecular Characterization of Olfactory Bulb Neural Stem Cells during Proliferation and Differentiation. J. Appl. Pharm. Sci. 2018, 8, 087–092. [Google Scholar]
- Buhrmann, C.; Busch, F.; Shayan, P.; Shakibaei, M. Sirtuin-1 (SIRT1) is required for promoting chondrogenic differentiation of mesenchymal stem cells. J. Biol. Chem. 2014, 289, 22048–22062. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Han, W.; You, S.; Ye, D.; Wang, L.; Wang, S.; Zhao, J.; Zhang, W.; Jiang, X.; Zhang, X. In vitro study of enhanced osteogenesis induced by HIF-1α-transduced bone marrow stem cells. Cell Prolif. 2011, 44, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, N.H.; Lee, Y.M.; Kim, J.S. Preventive effect of chlorogenic acid on lens opacity and cytotoxicity in human lens epithelial cells. Biol. Pharm. Bull. 2011, 34, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Chen, X.; Whitener, R.J.; Boder, E.T.; Jones, J.O.; Porollo, A.; Chen, J.; Zhao, L. Effects of parabens on adipocyte differentiation. Toxicol.Sci. Off. J. Soc. Toxicol. 2013, 131, 56–70. [Google Scholar] [CrossRef]
- Pak, C.S.; Heo, C.Y.; Shin, J.; Moon, S.Y.; Cho, S.W.; Kang, H.J. Effects of a Catechol-Functionalized Hyaluronic Acid Patch Combined with Human Adipose-Derived Stem Cells in Diabetic Wound Healing. Int. J. Mol. Sci. 2021, 22, 2632. [Google Scholar] [CrossRef]
- Iso, H.; Date, C.; Wakai, K.; Fukui, M.; Tamakoshi, A. The relationship between green tea and total caffeine intake and risk for self-reported type 2 diabetes among Japanese adults. Ann. Intern. Med. 2006, 144, 554–562. [Google Scholar] [CrossRef]
- Venables, M.C.; Hulston, C.J.; Cox, H.R.; Jeukendrup, A.E. Green tea extract ingestion, fat oxidation, and glucose tolerance in healthy humans. Am. J. Clin. Nutr. 2008, 87, 778–784. [Google Scholar] [CrossRef]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef]
- El-Magd, M.A.; Zedan, A.M.G.; Zidan, N.S.; Sakran, M.I.; Bahattab, O.; Oyouni, A.A.A.; Al-Amer, O.M.; Alalawy, A.I.; Elmoslemany, A.M. Avocado Seeds-Mediated Alleviation of Cyclosporine A-Induced Hepatotoxicity Involves the Inhibition of Oxidative Stress and Proapoptotic Endoplasmic Reticulum Stress. Molecules 2022, 27, 7859. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, F.; Saadeldin, I.; Alkarim, S. Ameliorative Effect of Mesenchymal Stem Cells-derived Exosomes on Diethylnitrosamine-induced Liver Injury in Albino Rats. Int. J. Pharmacol. 2018, 14, 1128–1135. [Google Scholar]
- Carey, A.N.; Fisher, D.R.; Bielinski, D.F.; Cahoon, D.S.; Shukitt-Hale, B. Walnut-Associated Fatty Acids Inhibit LPS-Induced Activation of BV-2 Microglia. Inflammation 2020, 43, 241–250. [Google Scholar] [CrossRef]
- Saleh, A.A.; Amber, K.; El-Magd, M.A.; Atta, M.S.; Mohammed, A.A.; Ragab, M.M.; Abd El-Kader, H. Integrative effects of feeding Aspergillus awamori and fructooligosaccharide on growth performance and digestibility in broilers: Promotion muscle protein metabolism. BioMed Res. Int. 2014, 2014, 946859. [Google Scholar] [CrossRef] [PubMed]
- Zedan, A.M.G.; Sakran, M.I.; Bahattab, O.; Hawsawi, Y.M.; Al-Amer, O.; Oyouni, A.A.A.; Nasr Eldeen, S.K.; El-Magd, M.A. Oriental Hornet (Vespa orientalis) Larval Extracts Induce Antiproliferative, Antioxidant, Anti-Inflammatory, and Anti-Migratory Effects on MCF7 Cells. Molecules 2021, 26, 3303. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Retention Time (min) | Amount (µg/g) |
|---|---|---|
| WKO extract | ||
| Gallic acid | 3.489 | 21.81 |
| 3-Hydroxytyrosol | 4.519 | 3.49 |
| Ferulic acid | 15.994 | 1.76 |
| o-Coumaric acid | 17.6 | 1.12 |
| Resveratrol | 19.21 | 9.41 |
| Quercitin | 20.9 | 1.70 |
| Rosemarinic | 22.1 | 1.43 |
| Kaempferol | 25.024 | 3.21 |
| Total | 43.93 | |
| WKD extract | ||
| Quinol | 3.15 | 52.26 |
| Gallic acid | 3.65 | 58.50 |
| Catechol | 5.464 | 21.93 |
| p-Hydroxy benzoic acid | 7.46 | 73.74 |
| Catechin | 9.069 | 11.70 |
| Chlorogenic acid | 9.47 | 38.47 |
| Syringic acid | 10.6 | 30.14 |
| p-Coumaric acid | 13.8 | 16.41 |
| Benzoic acid | 14.163 | 280.00 |
| Ferulic acid | 15.4 | 5.80 |
| Rutin | 16.27 | 19.02 |
| Ellagic | 16.8 | 6.50 |
| o-Coumaric acid | 17.3 | 3.90 |
| Total | 618.37 | |
| Phase | Control (DMSO) Group | WKO Group | WKD Group |
|---|---|---|---|
| G0/G1 | 70.72± 2.50 b | 88.43± 0.9 a | 92.2± 1.12 a |
| S | 18.25± 2.05 a | 6.00 ± 0.07 b | 2.70 ± 0.05 b |
| G2/M | 8.50 ± 0.40 a | 0.15 ± 0.01 b | 0.02 ± 0.005 b |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Bax | ACACCTGAGCTGACCTTG | AGCCCATGATGGTTCTGATC |
| Caspase3 | GGTATTGAGACAGACAGTGG | CATGGGATCTGTTTCTTTGC |
| Bcl2 | ATCGCTCTGTGGATGACTGAGTAC | AGAGACAGCCAGGAGAAATCAAAC |
| MMP9 | TCGAAGGCGACCTCAAGTG | TTCGGTGTAGCTTTGGATCCA |
| TIMP1 | CGCAGCGAGGAGGTTTCTCAT | GGCAGTGATGTGCAAATTTCC |
| p53 | ATGGCTTCCACCTGGGCTTC | TGACCCACAACTGCACAGGGC |
| p21 | GAGGCCTCTTCCCCATCTTCT | AATTAAGACACACTGAATGAAGGCTAAG |
| SIRT1 | GGCACCGATCCTCGAACAAT | CGCTTTGGTGGTTCTGAAAGG |
| AMPK | TGAAGCCAGAGAACGTGTTG | ATAATTTGGCGATCCACAGC |
| NrF2 | CACATCCAGACAGACACCAGT | CTACAAATGGGAATGTCTCTGC |
| HO-1 | GGAAAGCAGTCATGGTCAGTCA | CCCTTCCTGTGTCTTCCTTTGT |
| B-actin | AAGTCCCTCACCCTCCCAAAAG | AAGCAATGCTGTCACCTTCCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsied, M.A.; Sharawi, Z.W.; Al-Amrah, H.; Hegazy, R.A.; Mohamed, A.E.; Saleh, R.M.; El-kholy, S.S.; Farrag, F.A.; Fayed, M.H.; El-Magd, M.A. Walnut Kernel Oil and Defatted Extracts Enhance Mesenchymal Stem Cell Stemness and Delay Senescence. Molecules 2023, 28, 6281. https://doi.org/10.3390/molecules28176281
Elsied MA, Sharawi ZW, Al-Amrah H, Hegazy RA, Mohamed AE, Saleh RM, El-kholy SS, Farrag FA, Fayed MH, El-Magd MA. Walnut Kernel Oil and Defatted Extracts Enhance Mesenchymal Stem Cell Stemness and Delay Senescence. Molecules. 2023; 28(17):6281. https://doi.org/10.3390/molecules28176281
Chicago/Turabian StyleElsied, Marwa A., Zeina W. Sharawi, Hadba Al-Amrah, Rabab A. Hegazy, Amro E. Mohamed, Rasha M. Saleh, Sanad S. El-kholy, Foad A. Farrag, Masoud H. Fayed, and Mohammed A. El-Magd. 2023. "Walnut Kernel Oil and Defatted Extracts Enhance Mesenchymal Stem Cell Stemness and Delay Senescence" Molecules 28, no. 17: 6281. https://doi.org/10.3390/molecules28176281
APA StyleElsied, M. A., Sharawi, Z. W., Al-Amrah, H., Hegazy, R. A., Mohamed, A. E., Saleh, R. M., El-kholy, S. S., Farrag, F. A., Fayed, M. H., & El-Magd, M. A. (2023). Walnut Kernel Oil and Defatted Extracts Enhance Mesenchymal Stem Cell Stemness and Delay Senescence. Molecules, 28(17), 6281. https://doi.org/10.3390/molecules28176281