
An Attention towards the Prophylactic and Therapeutic Options of Phytochemicals for SARS-CoV-2: A Molecular Insight


 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Are Immunological and Pathophysiological Findings Helping in the Treatment, Prevention and Control of SARS-CoV-2?
3. Is the Drug-Repurposing Approach Sufficient? Or Novel Drug Development Is the Need to Curb SARS-CoV-2
4. Mucormycosis and SARS-CoV-2 Comorbidity
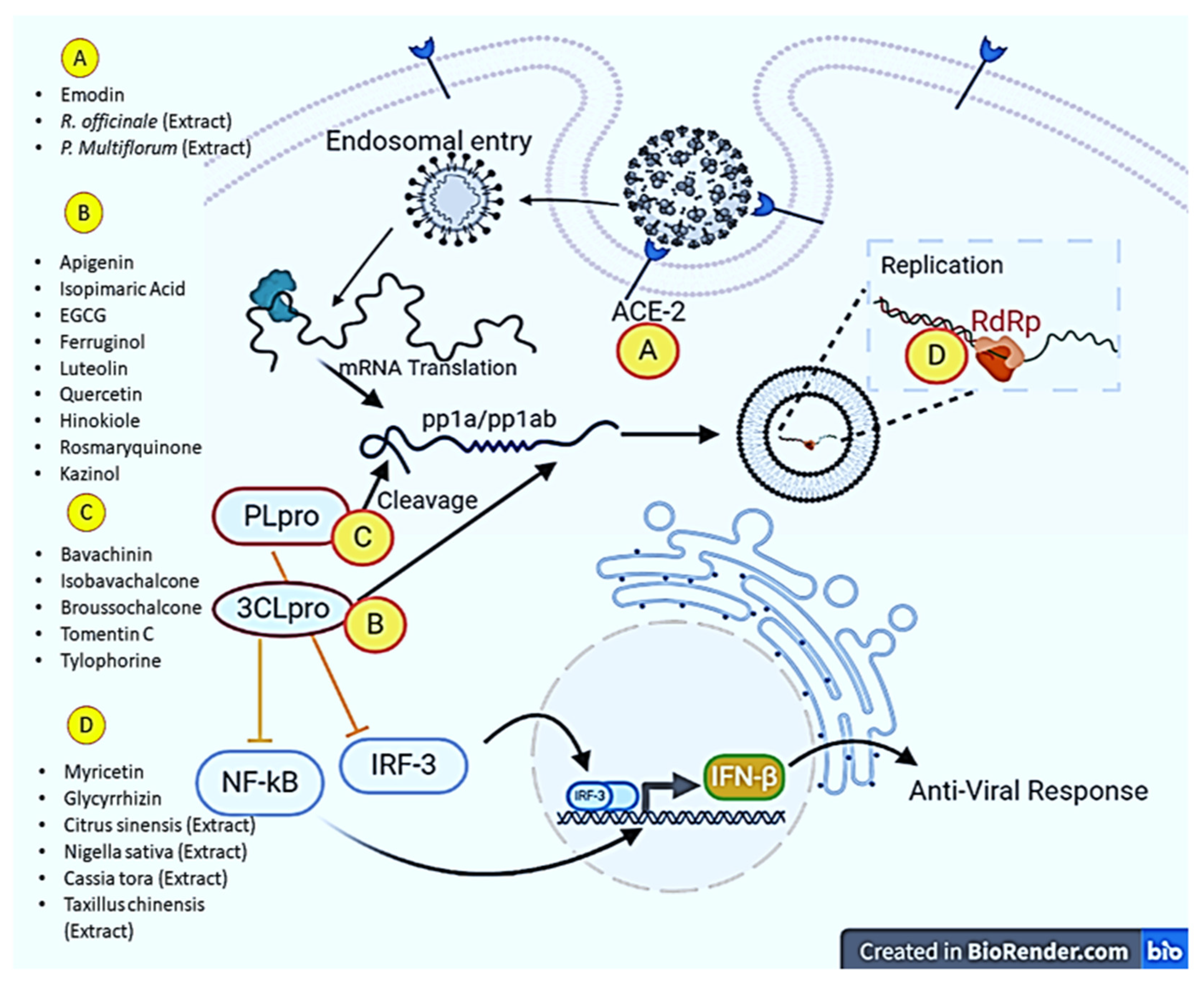
5. Can Plant Products Overcome the Burden of SARS-CoV-2?
5.1. Plant Products with Anti-Inflammatory and Antioxidant Potential in Various Diseases

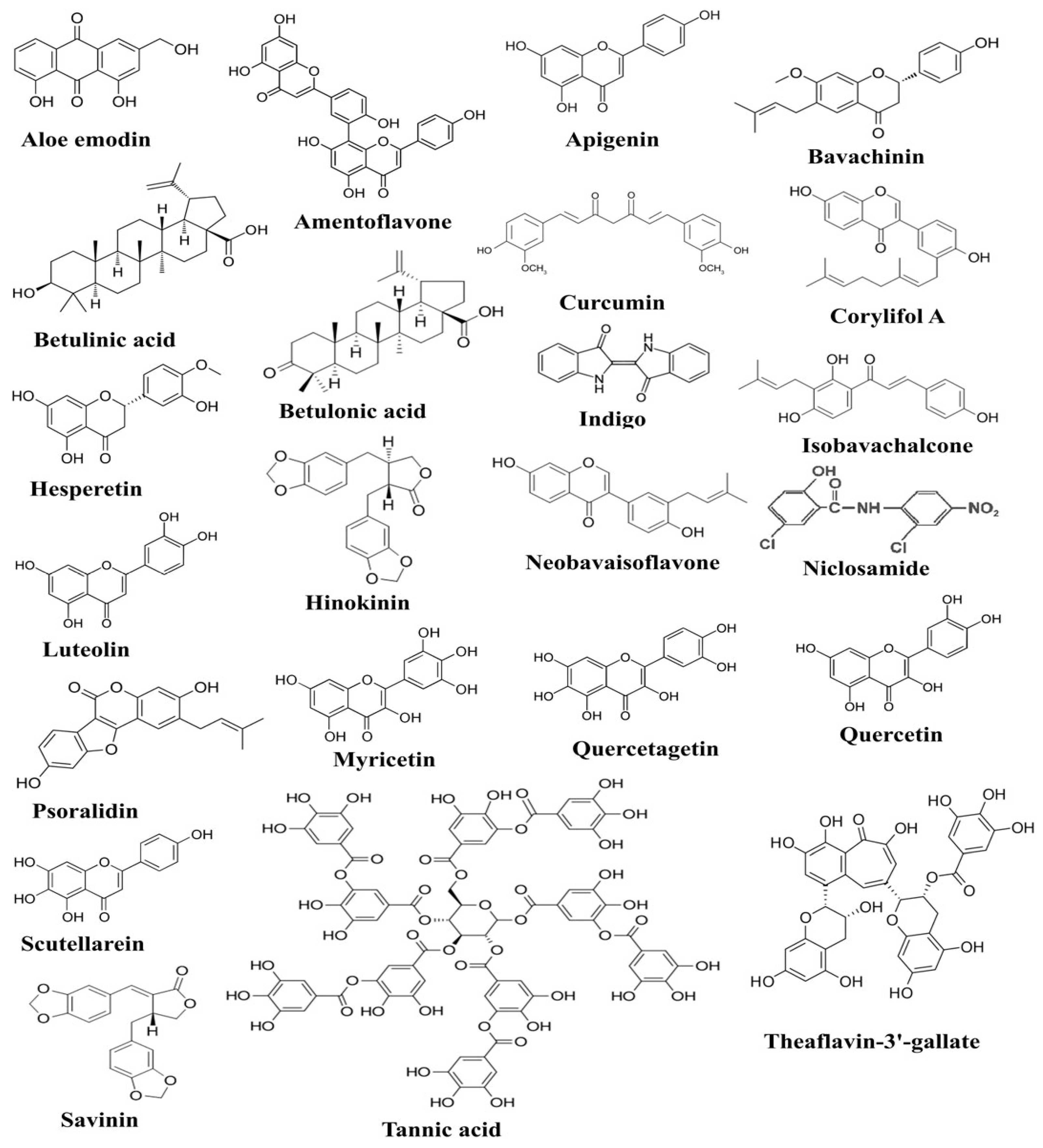{kind=link}
{kind=link}
| Class | Plant Product | Effects | Reference |
|---|---|---|---|
| Phenolics | Curcumin | Inhibits TNF-α, serum IL-6 and CRP production. | [65,67] |
| Phenolics | Chloroquine | Inhibits IFN-γ, IL-1, IL-6 and TNF-α production. | [68,69] |
| Phenolics | Punicalagin | Inhibits IL-6 and TNF-α production. | [81,82] |
| Phenolics | Ascorbic acid | Inhibits IFN-γ, TNF-α and IL-6 production. | [73,74] |
| Anthraquinone | Emodin | Inhibits binding of spike protein to ACE2 receptor. | [77] |
| Flavanol | EGCG | Inhibits IL-6, TNF-α and IL-8 production. | [78] |
| Phenolics | Hydroxychloroquine | Inhibits IL-1, IL-6 IFN-γ and TNF-α production. | [70,71] |
| Flavone | Luteolin | Inhibits production of TNF-α-induced MCP-1, ICAM-1 and VCAM-1. | [87] |
| Flavone | Apigenin | Inhibits TNF-α, IL-6, IL-8, and GM-CSF expression. | [89,90] |
| Carotenoid | Fucoxanthin | Decreases TNF-α production. | [73] |
| Phenolics | Resveratrol | Inhibits TNF-α, IL-8 and IL-6 and MCP-1 expression. | [84] |
| Phenolics | Gallic acid | Inhibits IL-6 production. | [81] |
| Phenolics | Chebulagic acid (Triphala) | Inhibits IL-6, IL-8 and MCP-1 expression. | [83] |
| Phenolics | Chebulinic acid (Triphala) | Inhibits IL-6, IL-8 and MCP-1 expression. | [83] |
| Flavonol | Quercetin | Inhibits IL-6, TNF-α and IL-1β production. | [86] |
| Flavonol | Fisetin | Inhibits IL-6, TNF-α, MCP-1 and IL-8 production. | [91] |
| Flavone | Rutin | Inhibits expression of NO and TNF-α. | [92] |
| Flavonol | Kaempferol | Inhibits secretion of MCP-1. | [94] |
| Phenolics | Ellagic acid | Inhibits IL-6 production. | [81] |
| Flavonol | Myricetin | Inhibits IL-6, TNF-α, NO, and iNOS. | [96] |
| Flavanone | Hesperetin | Inhibits IL-1β, IL-6 and TNF-α production. | [97,98] |
| Flavanone | Naringenin | Inhibits IL-6 and TNF-α production. | [99] |
| Glycoside | Isorhamnetin | Inhibits IL-6 production. | [101] |
| Flavone | Chrysin | Inhibits IL-1β expression. | [102] |
| Flavonol | Galangin | Inhibits IL-1β expression. | [102] |
| Phenolics | Theaflavin | Inhibits expression of IL-6, MCP-1 and ICAM-1 | [103] |
| Isoflavonoid | Genistein | Inhibits IL-6 and TNF-α production. | [104] |
| Flavone | Baicalein | Inhibits production of IL-6, TNF-α, IL-1β and NO. | [106] |
| Phenolics | Ferulic acid | Inhibits production TNF-α, IL-6 and IL-10 | [107] |
| Flavonol | Kaempferol7-o-β-d-glucoside | Inhibits expression of TNF-α, IL-6, IL-1β, iNOS and COX-2. | [95] |
5.2. Antiviral Activities of Plant Products: An Insight into Molecular Mechanisms
| Plant/Plant Products | Effects | References |
|---|---|---|
| Myricetin | Inhibits nsP13 (SARS-CoV helicase). | [111] |
| Scutellarein | Inhibits nsP13 (SARS-CoV helicase). | [111] |
| Gallocatechin gallate | Inhibits SARS-CoV 3CL pro. | [112] |
| Luteolin | Inhibits SARS-CoV 3CL pro. | [115] |
| Hesperetin (Isatisindigotica) | Inhibits cleavage activity of the 3CL pro of SARS-CoV. | [118] |
| Glycyrrhizin (Glycyrrhiza glabra) | Inhibits SARS-CoV replication. | [120] |
| Cinanserin (Houttuynia cordata) | Inhibits SARS-CoV 3CL pro. | [122] |
| Lycorine (Lycoris radiata) | Inhibits SARS-CoV activities. | [123] |
| Quercetin | Inhibits SARS-CoV 3CL pro. | [112] |
| Isobavachalcone (Psoralea corylifolia) | Inhibits SARS-CoV PL pro. | [141] |
| Quercitrin (Houttuynia cordata) | Inhibits SARS-CoV growth. | [117] |
| Isoquercitrin (Houttuynia cordata) | Inhibits SARS-CoV growth. | [117] |
| Apigenin | Inhibits SARS-CoV 3CL pro. | [115] |
| Emodin | Inhibits interactions between spike protein and ACE2 receptor protein. | [77] |
| Sinigrin (Isatisindigotica) | Inhibits cleavage activity of the 3CL pro of SARS-CoV. | [118] |
| Tomentin A (Paulownia tomentosa) | Inhibits SARS-CoV papain-like protease. | [139] |
| Lycorisradiata (ethanolic extract) | Inhibits SARS-CoV activities. | [123] |
| Methyl dehydroabietate | Inhibits SARS-CoV 3CL pro. | [115] |
| 18-hydroxyferruginol | Inhibits SARS-CoV 3CL pro. | [115] |
| Anthemis hyaline | Inhibits SARS-CoV replication. | [130] |
| Aloe emodin (Isatisindigotica) | Inhibits cleavage activity of the 3CL pro of SARS-CoV. | [118] |
| Rheum palmatum L. | Inhibits SARS-CoV 3CL pro. | [127] |
| O-acetyl-18-hydroxyferruginol | Inhibits SARS-CoV 3CL pro. | [115] |
| Rosmariquinone (Salvia miltiorrhiza) | Inhibits SARS-CoV 3CL pro. | [140] |
| Prunella vulgaris | Blocks viral entry. | [129] |
| Tomentin B (Paulownia tomentosa) | Inhibits SARS-CoV papain-like protease. | [139] |
| Rheum officinale Baill | Inhibits interactions between spike protein and ACE2 receptor protein. | [77] |
| Houttuynia cordata Thunb | Inhibits SARS-CoV 3CL pro and RdRp. | [125] |
| Cibotium barometz | Inhibits SARS-CoV 3CL pro. | [126] |
| Dihydrotanshinone I (Salvia miltiorrhiza) | Inhibits SARS-CoV 3CL pro. | [140] |
| Toona sinensis Roem | Inhibits SARS-CoV replication. | [128] |
| Citrus sinensis | Inhibits SARS-CoV replication. | [130] |
| 18-oxoferruginol | Inhibits SARS-CoV 3CL pro. | [115] |
| Bavachinin (Psoralea corylifolia) | Inhibits SARS-CoV PL pro. | [141] |
| Polygonum multiflorum Thunb | Inhibits interactions between spike protein and ACE2 receptor protein. | [77] |
| Isopimaric acid | Inhibits SARS-CoV 3CL pro. | [115] |
| Pyrrosia lingua (chloroform extract) | Inhibits SARS-CoV activities. | [123] |
| EGCG | Inhibits SARS-CoV 3CL pro. | [112] |
| Nigella sativa | Inhibits SARS-CoV replication. | [130] |
| Hinokiol | Inhibits SARS-CoV 3CL pro. | [115] |
| Tomentin C (Paulownia tomentosa) | Inhibits SARS-CoV papain-like protease. | [139] |
| Amentoflavone | Inhibits SARS-CoV 3CL pro. | [115] |
| Artemisia annua L. (ethanolic extract) | Inhibits SARS-CoV activities. | [123] |
| Gentiana scabra | Inhibits viral replication and proliferation in Vero E6 cells. | [126] |
| Saussurealappa | Blocks viral entry. | [129] |
| Tomentin D (Paulownia tomentosa) | Inhibits SARS-CoV papain-like protease. | [139] |
| Psoralidin (Psoralea corylifolia) | Inhibits SARS-CoV PL pro. | [141] |
| Corylifol A (Psoralea corylifolia) | Inhibits SARS-CoV PL pro. | [141] |
| 4′-o-methylbavachalcone (Psoralea corylifolia) | Inhibits SARS-CoV PL pro. | [141] |
| Tomentin E (Paulownia tomentosa) | Inhibits SARS-CoV papain-like protease. | [139] |
| Cassia tora | Inhibits viral replication and proliferation in Vero E6 cells. | [126] |
| Papyriflavonol A (Broussonetiapapyrifera) | Inhibits SARS-CoV PL pro and 3CL pro. | [140] |
| Broussochalcone A (Broussonetiapapyrifera) | Inhibits SARS-CoV PL pro and 3CL pro. | [140] |
| Kayadiol | Inhibits SARS-CoV 3CL pro. | [115] |
| KazinolJ (Broussonetiapapyrifera) | Inhibits SARS-CoV PL pro and 3CL pro. | [140] |
| Tanshinone IIB (Salvia miltiorrhiza) | Inhibits SARS-CoV 3CL pro. | [140] |
| Neobavaisoflavone (Psoralea corylifolia) | Inhibits SARS-CoV PL pro. | [141] |
| Dioscorea batatas | Inhibits SARS-CoV 3CL pro. | [126] |
| Ferruginol | Inhibits SARS-CoV 3CL pro. | [115] |
| Broussochalcone B (Broussonetiapapyrifera) | Inhibits SARS-CoV PL pro and 3CL pro. | [140] |
| Tylophorine (Tylophora indica) | Inhibits SARS-CoV PL pro. | [142] |
| Taxillus chinensis | Inhibits viral replication and proliferation in Vero E6 cells. | [126] |
| Cryptotanshinone (Salvia miltiorrhiza) | Inhibits SARS-CoV PL pro. | [140] |
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bogoch, I.I.; Watts, A.; Thomas-Bachli, A.; Huber, C.; Kraemer, M.U.; Khan, K. Pneumonia of Unknown Etiology in Wuhan, China: Potential for International Spread Via Commercial Air Travel. J. Travel Med. 2020, 13, 27. [Google Scholar]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 13, 536–544.
- Hageman, J.R. The coronavirus disease 2019 (COVID-19). Ped. Ann. 2020, 49, e99–e100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor recognition by the novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS coronavirus. J. Viro. 2020, 94, e00127-20. [Google Scholar] [CrossRef] [Green Version]
- Jaimes, J.A.; Millet, J.K.; Stout, A.E.; André, N.M.; Whittaker, G.R. A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses. Viruses 2020, 12, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, J.; Li, J.; Li, X.; Qi, X. CT imaging of the 2019 novel coronavirus (2019-nCoV)pneumonia. Radiology 2020, 295, 18. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.R.; Cao, Q.D.; Hong, Z.S.; Tan, Y.Y.; Chen, S.D.; Jin, H.J.; Yan, Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak–an update on the status. Mil. Med. Res. 2020, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Poutanen, S.M.; Low, D.E.; Henry, B.; Finkelstein, S.; Rose, D.; Green, K.; Chan, A.K. Identification of severe acute respiratory syndrome in Canada. N. Eng. J. Med. 2003, 348, 1995–2005. [Google Scholar] [CrossRef]
- Wang, W.; Tang, J.; Wei, F. Updated understanding of the outbreak of 2019 novel coronavirus (2019-nCoV) in Wuhan, China. J. Med. Virol. 2020, 92, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.L.; Wang, Y.M.; Wu, Z.Q.; Xiang, Z.C.; Guo, L.; Xu, T.; Fan, G.H. Identification of a novel coronavirus causing severe pneumonia in human: A descriptive study. Chin. Med. J. 2020, 133, 1015–1024. [Google Scholar] [CrossRef]
- Carlos, W.G.; Dela Cruz, C.S.; Cao, B.; Pasnick, S.; Jamil, S. Novel Wuhan (2019-nCoV)coronavirus. Am. J. Respir. Crit. Care Med. 2020, 201, P7–P8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Cheng, Z. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72 314 cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Xing, X. Early transmission dynamics in Wuhan, China, of novel coronavirus–infected pneumonia. N. Eng. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Müller, M.A. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Chen, H.D. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.K.; Huang, I.C.; Xu, K.; Vasilieva, N.; et al. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J. 2005, 24, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Yu, T. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Wang, Y.; Zhu, T.; Xia, L. CT features of coronavirus disease 2019 (COVID-19) pneumonia in 62 patients in Wuhan, China. Am. J. Roent. 2020, 214, 1287–1294. [Google Scholar] [CrossRef]
- Mustafić, S.; Jusufović, E.; Hukić, F.; Trnačević, E.; Divković, A.; Trnačević, A. Early predictors of severity and mortality in COVID-19 hospitalized patients. Med. Glas. 2021, 18, 384–393. [Google Scholar]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Guan, L. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensiv. Care Med. 2020, 46, 846–848. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Qi, F.; Shi, K.; Li, Y.; Li, J.; Chen, Y.; Luo, Y. Thalidomide combined with low-dose glucocorticoid in the treatment of COVID-19 pneumonia. Preprints 2020, 2020020395. Available online: https://www.preprints.org/manuscript/202002.0395/v1 (accessed on 20 December 2022).
- Zhang, W.; Du, R.H.; Li, B.; Zheng, X.S.; Yang, X.L.; Hu, B.; Zhou, P. Molecular and serological investigation of 2019-nCoV infected patients: Implication of multiple shedding routes. Emer. Micr. Inf. 2020, 9, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [Green Version]
- Madjid, M.; Safavi-Naeini, P.; Solomon, S.D.; Vardeny, O. Potential effects of coronaviruses on the cardiovascular system: A review. JAMA Cardiol. 2020, 5, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Zhao, Y. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Grasselli, G.; Zangrillo, A.; Zanella, A.; Antonelli, M.; Cabrini, L.; Castelli, A.; Iotti, G. Baseline Characteristics and Outcomes of 1591 Patients Infected With SARS-CoV-2 Admitted to ICUs of the Lombardy Region, Italy. JAMA 2020, 323, 1574–1581. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhao, Y.; Zhang, F.; Wang, Q.; Li, T.; Liu, Z.; Zeng, X. The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The experience of clinical immunologists from China. Clin. Immunol. 2020, 214, 108393. [Google Scholar] [CrossRef]
- Bhat, S.A.; Sher, F.; Kumar, R.; Karahmet, E.; Haq, S.A.U.; Zafar, A.; Lima, E.C. Environmental and health impacts of spraying COVID-19 disinfectants with associated challenges. Environ. Sci. Pollut. Res. 2022, 29, 85648–85657. [Google Scholar] [CrossRef] [PubMed]
- Arabi, Y.M.; Balkhy, H.H.; Hayden, F.G.; Bouchama, A.; Luke, T.; Baillie, J.K.; Nguyen-Van-Tam, J.S. Middle East respiratory syndrome. N. Eng. J. Med. 2017, 376, 584–594. [Google Scholar] [CrossRef]
- Li, G.; De Clercq, E. Therapeutic options for the 2019 novel coronavirus (2019-nCoV). Nature 2020, 19, 149–150. [Google Scholar] [CrossRef] [PubMed]
- Eltahla, A.A.; Luciani, F.; White, P.A.; Lloyd, A.R.; Bull, R.A. Inhibitors of the hepatitis C virus polymerase; mode of action and resistance. Viruses 2015, 7, 5206–5224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.; Kotian, P.; Warren, T.; Panchal, R.; Bavari, S.; Julander, J.; Babu, Y. BCX4430–a broad-spectrum antiviral adenosine nucleoside analog under development for the treatment of Ebola virus disease. J. Infect. Public. Health 2016, 9, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Ray, A.S. Coronavirus susceptibility to the antiviral remdesivir (GS-5734) is mediated by the viral polymerase and the proofreading exoribonuclease. mBio 2018, 9, e00221-18. [Google Scholar] [CrossRef] [Green Version]
- Warren, T.K.; Jordan, R.; Lo, M.K.; Ray, A.S.; Mackman, R.L.; Soloveva, V.; Larson, N. Therapeutic efficacy of the small molecule GS-5734 against Ebola virus in rhesus monkeys. Nature 2016, 531, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Sheahan, T.P.; Sims, A.C.; Graham, R.L.; Menachery, V.D.; Gralinski, L.E.; Case, J.B.; Bannister, R. Broad-spectrum antiviral GS-5734 inhibits both epidemic and zoonotic coronaviruses. Sci. Transl. Med. 2017, 9, eaal3653. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, Y.M.; Xu, J.Y.; Cao, B. Potential antiviral therapeutics for 2019 Novel Coronavirus. ZhonghuaJie He He Hu Xi Za Zhi Chin. J. Tuber. Resp. Dis. 2020, 43, E002. [Google Scholar]
- Savarino, A.; Di Trani, L.; Donatelli, I.; Cauda, R.; Cassone, A. New insights into the antiviral effects of chloroquine. Lanc. Infect. Dis. 2006, 6, 67–69. [Google Scholar] [CrossRef]
- Yan, Y.; Zou, Z.; Sun, Y.; Li, X.; Xu, K.F.; Wei, Y.; Jiang, C. Anti-malaria drug chloroquine is highly effective in treating avian influenza A H5N1 virus infection in an animal model. Cell Res. 2013, 23, 300–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, M.J.; Bergeron, E.; Benjannet, S.; Erickson, B.R.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol. J. 2005, 2, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Chan, K.W.; Wong, V.T.; Tang, S.C.W. COVID-19: An update on the epidemiological, clinical, preventive and therapeutic evidence and guidelines of integrative Chinese–Western medicine for the management of 2019 novel coronavirus disease. Am. J. Chin. Med. 2020, 48, 737–762. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, H.K.; Mehra, M.R. COVID-19 illness in native and immunosuppressed states: A clinical-therapeutic staging proposal. J. Heart. Lung. Transp. 2020, 39, 405–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinaei, R.; Pezeshki, S.; Asadipour, A.; Shiari, R.; Sinaei, R.; Sinaei, A. Anti-rheumatic drugs as potential anti-inflammatory, immunomodulatory agents against COVID-19: A systematic review. Pharm. Sci. 2021, 27, S13–S28. [Google Scholar] [CrossRef]
- Stebbing, J.; Phelan, A.; Griffin, I.; Tucker, C.; Oechsle, O.; Smith, D.; Richardson, P. COVID-19: Combining antiviral and anti-inflammatory treatments. Lancet Infect. Dis. 2020, 20, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F. Nitazoxanide: A first-in-class broad-spectrum antiviral agent. Antivir. Res. 2014, 110, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Shen, R.; He, J.; Li, X.; Guo, X. Molecular Modeling Evaluation of the Binding Effect of Ritonavir, Lopinavir and Darunavir to Severe Acute Respiratory Syndrome Coronavirus 2 Proteases. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Jeon, S.; Shin, H.Y.; Kim, M.J.; Seong, Y.M.; Lee, W.J.; Park, S.J. Case of the index patient who caused tertiary transmission of COVID-19 infection in Korea: The application of lopinavir/ritonavir for the treatment of COVID-19 infected pneumonia monitored by quantitative RT-PC. J. Kor. Med. Sci. 2020, 35, e79. [Google Scholar] [CrossRef]
- Bian, H.; Zheng, Z.H.; Wei, D.; Zhang, Z.; Kang, W.Z.; Hao, C.Q.; Dong, K.; Kang, W.; Xia, J.L.; Miao, J.L.; et al. Meplazumab treats COVID-19 pneumonia: An open-labelled, concurrent controlled add-on clinical trial. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Chen, W.; Zhou, Y.S.; Lian, J.Q.; Zhang, Z.; Du, P.; Gong, L.; Zhang, Y.; Cui, H.Y.; Geng, J.J.; et al. SARS-CoV-2 invades host cells via a novel route: CD147-spike protein. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wu, Z.; Li, J.W.; Zhao, H.; Wang, G.Q. The cytokine release syndrome (CRS) of severe COVID-19 and Interleukin-6 receptor (IL-6R) antagonist Tocilizumab may be the key to reduce the mortality. Int. J. Antimicrob. Age 2020, 55, 105954. [Google Scholar] [CrossRef]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhang, X. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Chappell, J.D. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Trans. Med. 2020, 12, eabb5883. [Google Scholar] [CrossRef] [Green Version]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved Drug Ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef]
- Pushparaj, K.; Bhotla, H.K.; Arumugam, V.A.; Pappusamy, M.; Easwaran, M.; Liu, W.C.; Issara, U.; Rengasamy, K.R.R.; Meyyazhagan, A.; Balasubramanian, B. Mucormycosis (black fungus) ensuing COVID-19 and comorbidity meets—Magnifying global pandemic grieve and catastrophe begins. Sci. Total Environ. 2022, 805, 150355. [Google Scholar] [CrossRef]
- Arjmand, G.; Askari, E.; Kazemi, A.; Zarei, E.; Haseli, S.; Sadraei, N. Bronchial wall necrosis secondary to mucormycosis following SARS-CoV-2 infection: A case report. Radiol. Case Rep. 2022, 17, 2956–2959. [Google Scholar] [CrossRef]
- Ahmed, O.F.; Al-Neaimy, S.; Kakamad, F.H.; Mikael, T.M.; Hamasaeed, A.G.; Salih, R.Q.; Salh, A.M.; Hamasaeed, M.G. COVID-19 associated with pulmonary mucormycosis; a case series. Ann. Med. Surg. 2022, 76, 103434. [Google Scholar] [CrossRef]
- Dabas, A.; Swarnim, S.; Sagar, T.; Mantan, M. Mucormycosis with Severe Acute Respiratory Syndrome Coronavirus-2 Patients on Hemodialysis: A Case Series with Review of Literature. Saudi J. Kidney Dis. Transplant. 2021, 32, 1152. [Google Scholar]
- Mekonnen, Z.K.; Ashraf, D.C.; Jankowski, T.; Grob, S.R.; Vagefi, M.R.; Kersten, R.C.; Simko, J.P.; Winn, B.J. Acute invasive rhino-orbital mucormycosis in a patient with COVID-19-associated acute respiratory distress syndrome. Ophthalmic Plast. Reconstr. Surg. 2020, 37, e40–e80. [Google Scholar] [CrossRef] [PubMed]
- Reddy, Y.M.; Yeduguri, S.; Parida, S.; Kamatham, S.N.; Pidaparthi, L.; Jaiswal, S.K.; Sadhvani, B.; Tourani, V.; Kumar, S.; Challa, S.; et al. Pathogenetic factors fanning the flames of COVID-19 to cause rhino-orbito-cerebral mucormycosis: An observational study. J. Med. Mycol. 2022, 32, 101252. [Google Scholar] [CrossRef] [PubMed]
- Majnooni, M.B.; Fakhri, S.; Bahrami, G.; Naseri, M.; Farzaei, M.H.; Echeverría, J. Alkaloids as potential phytochemicals against SARS-CoV-2: Approaches to the associated pivotal mechanisms. Evid.-Based Complement. Altern. Med. 2021, 2021, 6632623. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Yao, J.; Li, Y.; Hu, X.; Shao, H.; Tian, X. Anti-inflammatory and antioxidant effects of curcumin on acute lung injury in a rodent model of intestinal ischemia reperfusion by inhibiting the pathway of NF-Kb. Int. J. Clin. Exp. Pathol. 2015, 8, 3451. [Google Scholar]
- Ghandadi, M.; Sahebkar, A. Curcumin: An effective inhibitor of interleukin-6. Curr. Pharm. Des. 2017, 23, 921–931. [Google Scholar] [CrossRef]
- Mohammadi, S.; Kayedpoor, P.; Karimzadeh-Bardei, L.; Nabiuni, M. The effect of curcumin on TNF-α, IL-6 and CRP expression in a model of polycystic ovary syndrome as an inflammation state. J. Reprod. Infertil. 2017, 18, 352. [Google Scholar]
- Oh, S.; Shin, J.H.; Jang, E.J.; Won, H.Y.; Kim, H.K.; Jeong, M.G.; Hwang, E.S. Anti-inflammatory activity of chloroquine and amodiaquine through p21-mediated suppression of T cell proliferation and Th1 cell differentiation. Biochem. Biophys. Res. Commun. 2016, 474, 345–350. [Google Scholar] [CrossRef]
- Jang, C.H.; Choi, J.H.; Byun, M.S.; Jue, D.M. Chloroquin inhibits production of TNFalpha, interleukin-1 and interleukin-6 from lipopolysaccharide-stimulated human monocytes/macrophages by different modes. Rheumatology 2006, 45, 703–710. [Google Scholar] [CrossRef] [Green Version]
- Sperber, K.; Quraishi, H.U.M.A.; Kalb, T.H.; Panja, A.S.I.T.; Stecher, V.; Mayer, L. Selective regulation of cytokine secretion by hydroxychloroquine: Inhibition of interleukin 1 alpha (IL-1-alpha) and IL-6 in human monocytes and T cells. J. Rheumatol. 1993, 20, 803–808. [Google Scholar]
- Dijkmans, B.A.; Verweij, C.L. Chloroquine and hydroxychloroquine equally affect tumor necrosis factor-alpha, interleukin 6, and interferon-gamma production by peripheral blood mononuclear cells. J. Rheumatol. 1997, 24, 55–60. [Google Scholar]
- Ohishi, T.; Hishiki, T.; Baig, M.S.; Rajpoot, S.; Saqib, U.; Takasaki, T.; Hara, Y. Epigallocatechin gallate (EGCG) attenuates severe acute respiratory coronavirus disease 2 (SARS-CoV-2) infection by blocking the interaction of SARS-CoV-2 spike protein receptor-binding domain to human angiotensin-converting enzyme 2. PLoS ONE 2022, 17, e0271112. [Google Scholar] [CrossRef]
- Molina, N.; Morandi, A.C.; Bolin, A.P.; Otton, R. Comparative effect of fucoxanthin and vitamin C on oxidative and functional parameters of human lymphocytes. Int. Immunol. 2014, 22, 41–50. [Google Scholar] [CrossRef]
- Härtel, C.; Strunk, T.; Bucsky, P.; Schultz, C. Effects of vitamin C on intracytoplasmic cytokine production in human whole blood monocytes and lymphocytes. Cytokine 2004, 27, 101–106. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, G.; Yuan, J.; Wang, Y.; Yang, X.; Wang, X.; Zhong, N. Vitamin C mitigates oxidative stress and tumor necrosis factor-alpha in severe community-acquired pneumonia and LPS-induced macrophages. Mediat. Inflamm. 2014, 2014, 426740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhm, F.; Settergren, M.; Pernow, J. Vitamin C blocks vascular dysfunction and release of interleukin-6 induced by endothelin-1 in humans in vivo. Atherosclerosis 2007, 190, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.Y.; Wu, S.L.; Chen, J.C.; Li, C.C.; Hsiang, C.Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef]
- Shin, H.Y.; Kim, S.H.; Jeong, H.J.; Kim, S.Y.; Shin, T.Y.; Um, J.Y.; Kim, H.M. Epigallocatechin-3-gallate inhibits secretion of TNF-α, IL-6 and IL-8 through the attenuation of ERK and NF-κB in HMC-1 cells. Int. Arch. Allergy Immunol. 2007, 142, 335–344. [Google Scholar] [CrossRef]
- Li, M.; Liu, J.T.; Pang, X.M.; Han, C.J.; Mao, J.J. Epigallocatechin-3-gallate inhibits angiotensin II and interleukin-6-induced C-reactive protein production in macrophages. Pharmacol. Rep. 2012, 64, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.; Jha, A.K.; Hazra, B. Plant products as inhibitors of coronavirus 3CL protease. Front. Pharmacol. 2021, 12, 583387. [Google Scholar] [CrossRef] [PubMed]
- Ben Saad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A&B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in LPS-Induced RAW264. 7 Macrophages by Inhibiting FoxO3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 2794. [Google Scholar] [CrossRef] [Green Version]
- Shanmuganathan, S.; Angayarkanni, N. Chebulagic acid Chebulinic acid and Gallic acid, the active principles of Triphala, inhibit TNFα induced pro-angiogenic and pro-inflammatory activities in retinal capillary endothelial cells by inhibiting p38, ERK and NF-kB phosphorylation. Vasc. Pharmacol. 2018, 108, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, I.; Takeda, Y. Inhibitory effects of resveratrol on MCP-1, IL-6, and IL-8 production in human coronary artery smooth muscle cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2013, 386, 835–839. [Google Scholar] [CrossRef]
- Wang, X.L.; Li, T.; Li, J.H.; Miao, S.Y.; Xiao, X.Z. The effects of resveratrol on inflammation and oxidative stress in a rat model of chronic obstructive pulmonary disease. Molecules 2017, 22, 1529. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Diao, P.; Shu, X.; Li, L.; Xiong, L. Quercetin and Quercitrin Attenuates the Inflammatory Response and Oxidative Stress in LPS-Induced RAW264. 7 Cells: In vitro Assessment and a Theoretical Model. BioMed Res. Int. 2019, 2019, 7039802. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Nallasamy, P.; Liu, D.; Shah, H.; Li, J.Z.; Chitrakar, R.; Li, Y. Luteolin protects against vascular inflammation in mice and TNF-alpha-induced monocyte adhesion to endothelial cells via suppressing IΚBα/NF-κB signaling pathway. J. Nutr. Biochem. 2015, 26, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, X.; Zhang, L.; Virgous, C.; Si, H. Combination of curcumin and luteolin synergistically inhibits TNF-α-induced vascular inflammation in human vascular cells and mice. J. Nutr. Biochem. 2019, 73, 108222. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Lee, J.H.; Kwon, D.Y. Apigenin inhibits release of inflammatory mediators by blocking the NF-κB activation pathways in the HMC-1 cells. Immunopharmacol. Immunotoxicol. 2011, 33, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, G.; Gurley, E.C.; Zhou, H. Flavonoid apigenin inhibits lipopolysaccharide-induced inflammatory response through multiple mechanisms in macrophages. PLoS ONE 2014, 9, e107072. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.L.; Huang, W.C.; Cheng, S.C.; Liou, C.J. Fisetin inhibits the generation of inflammatory mediators in interleukin-1β–induced human lung epithelial cells by suppressing the Nf-κb and Erk1/2 pathways. Int. Immunopharmacol. 2018, 60, 202–210. [Google Scholar] [CrossRef]
- Guruvayoorappan, C.; Kuttan, G. Rutin inhibits nitric oxide and tumor necrosis factor-α production in lipopolysaccharide and concanavalin-A stimulated macrophages. Drug Metab. Drug Interact. 2007, 22, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Su, K.Y.; Yu, C.Y.; Chen, Y.P.; Hua, K.F.; Chen, Y.L.S. 3, 4-Dihydroxytoluene, a metabolite of rutin, inhibits inflammatory responses in lipopolysaccharide-activated macrophages by reducing the activation of NF-κBsignaling. BMC Complement. Altern. Med. 2014, 14, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilczok, T. Effect of kaempferol on the production and gene expression of monocyte chemoattractant protein-1 in J774. 2 macrophages. Pharmacol. Rep. 2005, 57, 107–112. [Google Scholar]
- Lee, S.B.; Shin, J.S.; Han, H.S.; Lee, H.H.; Park, J.C.; Lee, K.T. Kaempferol 7-O-β-D-glucoside isolated from the leaves of Cudraniatricuspidata inhibits LPS-induced expression of pro-inflammatory mediators through inactivation of NF-κB, AP-1, and JAK-STAT in RAW 264.7 macrophages. Chem.-Biol. Interact. 2018, 284, 101–111. [Google Scholar] [CrossRef]
- Cho, B.O.; Yin, H.H.; Park, S.H.; Byun, E.B.; Ha, H.Y.; Jang, S.I. Anti-inflammatory activity of myricetin from Diospyros lotus through suppression of NF-κB and STAT1 activation and Nrf2-mediated HO-1 induction in lipopolysaccharide-stimulated RAW264. 7 macrophages. Biosci. Biotechnol. Biochem. 2016, 80, 1520–1530. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Feng, X.; Ding, S. Hesperetin attenuates ventilator-induced acute lung injury through inhibition of NF-κB-mediated inflammation. Eur. J. Pharmacol. 2015, 769, 333–341. [Google Scholar] [CrossRef]
- Ren, H.; Hao, J.; Liu, T.; Zhang, D.; Lv, H.; Song, E.; Zhu, C. Hesperetin suppresses inflammatory responses in lipopolysaccharide-induced RAW 264.7 cells via the inhibition of NF-κB and activation of Nrf2/HO-1 pathways. Inflammation 2016, 39, 964–973. [Google Scholar] [CrossRef]
- Jin, L.; Zeng, W.; Zhang, F.; Zhang, C.; Liang, W. Naringenin ameliorates acute inflammation by regulating intracellular cytokine degradation. J. Immunol. 2017, 199, 3466–3477. [Google Scholar] [CrossRef] [Green Version]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Mizokami, S.S.; Borghi, S.M.; Bordignon, J.; Silva, R.L.; VerriJr, W.A. The citrus flavonone naringenin reduces lipopolysaccharide-induced inflammatory pain and leukocyte recruitment by inhibiting NF-κB activation. J. Nutr. Biochem. 2016, 33, 8–14. [Google Scholar] [CrossRef]
- Jin, J.Y.; Choi, E.Y.; Park, H.R.; Choi, J.I.; Choi, I.S.; Kim, S.J. Isorhamnetin inhibits P revotella intermedia lipopolysaccharide-induced production of interleukin-6 in murine macrophages via anti-inflammatory heme oxygenase-1 induction and inhibition of nuclear factor-κB and signal transducer and activator of transcription 1 activation. J. Periodontal Res. 2013, 48, 687–695. [Google Scholar]
- Bronska, M.; Czuba, Z.P.; Krol, W. Effect of flavone derivatives on interleukin 1β mRNA expression and IL-1β protein synthesis in stimulated RAW 264.7 macrophages. Scand. J. Immunol. 2003, 57, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Joo, Y.E. Theaflavin inhibits LPS-induced IL-6, MCP-1, and ICAM-1 expression in bone marrow-derived macrophages through the blockade of NF-κB and MAPK signaling pathways. Chonnam Med. J. 2011, 47, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Zhang, Y.; Yang, Q.; Cheng, S.; Hao, J.; Zhao, X.; Jiang, Z. Genistein suppresses LPS-induced inflammatory response through inhibiting NF-κB following AMP kinase activation in RAW 264.7 macrophages. PLoS ONE 2012, 7, e53101. [Google Scholar] [CrossRef]
- Choi, E.Y.; Bae, S.H.; Ha, M.H.; Choe, S.H.; Hyeon, J.Y.; Choi, J.I.; Kim, S.J. Genistein suppresses Prevotella intermedia lipopolysaccharide-induced inflammatory response in macrophages and attenuates alveolar bone loss in ligature-induced periodontitis. Arch. Oral Biol. 2016, 62, 70–79. [Google Scholar] [CrossRef]
- Qi, Z.; Yin, F.; Lu, L.; Shen, L.; Qi, S.; Lan, L.; Yin, Z. Baicalein reduces lipopolysaccharide-induced inflammation via suppressing JAK/STATs activation and ROS production. Inflamm. Res. 2013, 62, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Lampiasi, N.; Montana, G. The molecular events behind ferulic acid mediated modulation of IL-6 expression in LPS-activated Raw 264.7 cells. Immunobiology 2016, 221, 486–493. [Google Scholar] [CrossRef]
- Cheng, F.J.; Huynh, T.K.; Yang, C.S.; Hu, D.W.; Shen, Y.C.; Tu, C.Y.; Wu, Y.-C.; Tang, C.-H.; Huang, W.-C.; Chen, Y.; et al. Hesperidin Is a Potential Inhibitor against SARS-CoV-2 Infection. Nutrients 2021, 13, 2800. [Google Scholar] [CrossRef]
- Salgado-Benvindo, C.; Leijs, A.A.; Thaler, M.; Tas, A.; Arbiser, J.L.; Snijder, E.J.; van Hemert, M.J. Honokiol inhibits SARS-CoV-2 replication in cell culture. bioRxiv 2022. [Google Scholar] [CrossRef]
- Mitra, S.S.; Nandy, S.; Dey, A. Promising Plant-Based Bioactive Natural Products in Combating SARS-CoV-2 Novel Corona (COVID-19) Virus Infection. In Medicinal Plants for Lung Diseases; Springer: Singapore, 2021; pp. 497–514. [Google Scholar]
- Yu, M.S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.W.; Jee, J.G.; Jeong, Y.J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorg. Med. Chem. Lett. 2012, 22, 4049–4054. [Google Scholar] [CrossRef]
- Nguyen, T.T.H.; Woo, H.J.; Kang, H.K.; Kim, Y.M.; Kim, D.W.; Ahn, S.A.; Kim, D. Flavonoid-mediated inhibition of SARS coronavirus 3C-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Elkousy, R.H.; Said, Z.N.; Ali, M.A.; Kutkat, O. Anti-SARS-CoV-2 in vitro potential of Castor Oil Plant (Ricinus communis) Leaf Extract: In-Silico Virtual Evidence. Res. Sq. 2022, 1–34. [Google Scholar] [CrossRef]
- Ansari, W.A.; Ahamad, T.; Khan, M.A.; Khan, Z.A.; Khan, M.F. Exploration of Luteolin as Potential Anti-COVID-19 Agent: Molecular Docking, Molecular Dynamic Simulation, ADMET and DFT Analysis. Lett. Drug Des. Discov. 2022, 19, 741–756. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.Y.; Kim, D.; Rho, M.C. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef]
- Swargiary, A.; Mahmud, S.; Saleh, M.A. Screening of phytochemicals as potent inhibitor of 3-chymotrypsin and papain-like proteases of SARS-CoV-2: An in silico approach to combat COVID-19. J. Biomol. Struct. Dyn. 2022, 40, 2067–2081. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Yang, Z.; Wang, J.; Xu, Y.; Tan, R.X.; Li, E. Houttuynia cordata blocks HSV infection through inhibition of NF-κBactivation. Antivir. Res. 2011, 92, 341–345. [Google Scholar] [CrossRef]
- Lin, C.W.; Tsai, F.J.; Tsai, C.H.; Lai, C.C.; Wan, L.; Ho, T.Y.; Chao, P.D.L. Anti-SARS coronavirus 3C-like protease effects of Isatisindigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- Saravanan, K.M.; Zhang, H.; Senthil, R.; Vijayakumar, K.K.; Sounderrajan, V.; Wei, Y.; Shakila, H. Structural basis for the inhibition of SARS-CoV-2 main protease by Indian medicinal plant-derived antiviral compounds. J. Biomol. Struct. Dyn. 2022, 40, 1970–1978. [Google Scholar] [CrossRef]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, A.A.; Abdel-Wadood, Y.A. The potential of glycyrrhizin and licorice extract in combating COVID-19 and associated conditions. Phytomed. Plus 2021, 1, 100043. [Google Scholar] [CrossRef]
- Chen, L.; Gui, C.; Luo, X.; Yang, Q.; Günther, S.; Scandella, E.; Chen, J. Cinanserin is an inhibitor of the 3C-like proteinase of severe acute respiratory syndrome coronavirus and strongly reduces virus replication in vitro. J. Virol. 2005, 79, 7095–7103. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Y.; Chen, C.; Zhang, H.Q.; Guo, H.Y.; Wang, H.; Wang, L.; Li, R.S. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23. [Google Scholar] [CrossRef]
- Boozari, M.; Hosseinzadeh, H. Natural products for COVID-19 prevention and treatment regarding to previous coronavirus infections and novel studies. Phytother. Res. 2021, 35, 864–876. [Google Scholar] [CrossRef]
- Lau, K.M.; Lee, K.M.; Koon, C.M.; Cheung, C.S.F.; Lau, C.P.; Ho, H.M.; Tsui, S.K.W. Immunomodulatory and anti-SARS activities of Houttuynia cordata. J. Ethnopharmacol. 2008, 118, 79–85. [Google Scholar] [CrossRef]
- Wen, C.C.; Shyur, L.F.; Jan, J.T.; Liang, P.H.; Kuo, C.J.; Arulselvan, P.; Yang, N.S. Traditional Chinese medicine herbal extracts of Cibotium barometz, Gentiana scabra, Dioscorea batatas, Cassia tora, and Taxillus chinensis inhibit SARS-CoVreplication. J. Tradit. Complement. Med. 2011, 1, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Su, X.; Gong, S.; Qin, Y.; Liu, W.; Li, J.; Xu, Q. Anti-SARS coronavirus 3C-like protease effects of Rheum palmatum L. extracts. Biosci. Trends 2009, 3, 124–126. [Google Scholar]
- Chen, C.J.; Michaelis, M.; Hsu, H.K.; Tsai, C.C.; Yang, K.D.; Wu, Y.C.; Doerr, H.W. Toona sinensis Roem tender leaf extract inhibits SARS coronavirus replication. J. Ethnopharmacol. 2008, 120, 108–111. [Google Scholar] [CrossRef]
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Chen, L. Small molecules blocking the entry of severe acute respiratory syndrome coronavirus into host cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulasli, M.; Gurses, S.A.; Bayraktar, R.; Yumrutas, O.; Oztuzcu, S.; Igci, M.; Arslan, A. The effects of Nigella sativa (Ns), Anthemis hyalina(Ah) and Citrus sinensis (Cs) extracts on the replication of coronavirus and the expression of TRP genes family. Mol. Biol. Rep. 2014, 41, 1703–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jena, A.B.; Kanungo, N.; Nayak, V.; Chainy, G.B.N.; Dandapat, J. Catechin and curcumin interact with S protein of SARS-CoV-2 and ACE2 of human cell membrane: Insights from computational studies. Sci. Rep. 2021, 11, 2043. [Google Scholar] [CrossRef] [PubMed]
- Ungarala, R.; Munikumar, M.; Sinha, S.N.; Kumar, D.; Sunder, R.S.; Challa, S. Assessment of antioxidant, immunomodulatory activity of oxidised Epigallocatechin-3-Gallate (green tea polyphenol) and its action on the main protease of SARS-CoV-2—An in vitro and in silico approach. Antioxidants 2022, 11, 294. [Google Scholar] [CrossRef]
- Imam, S.S.; Imam, S.T.; Mdwasifathar, K.; Ammar, M.Y. Interaction Between Ace 2 and SARS-CoV-2, And Use of Egcg And Theaflavin to Treat Covid 19 in Initial Phases. Int. J. Curr. Pharm. Res. 2022, 14, 5–10. [Google Scholar] [CrossRef]
- Tuli, H.; Sood, S.; Pundir, A.; Choudhary, D.; Dhama, K.; Kaur, G.; Seth, P.; Vashishth, A.; Kumar, P. Molecular Docking studies of Apigenin, Kaempferol, and Quercetin as potential target against spike receptor protein of SARS COV. J. Exp. Biol. Agric. Sci. 2022, 10, 144–149. [Google Scholar] [CrossRef]
- Flores-Félix, J.D.; Gonçalves, A.C.; Alves, G.; Silva, L.R. Consumption of phenolic-rich food and dietary supplements as a key tool in SARS-CoV-19 infection. Foods 2021, 10, 2084. [Google Scholar] [CrossRef]
- Rossi, G.A.; Sacco, O.; Capizzi, A.; Mastromarino, P. Can resveratrol-inhaled formulations be considered potential adjunct treatments for COVID-19? Front. Immunol. 2021, 12, 670955. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Jung, J.-H.; Kim, M.-K.; Lim, S.; Choi, J.-M.; Chung, B.; Kim, D.-W.; Kim, D. The inhibitory effects of plant derivate polyphenols on the main protease of SARS coronavirus 2 and their structure–activity relationship. Molecules 2021, 26, 1924. [Google Scholar] [CrossRef] [PubMed]
- Clementi, N.; Scagnolari, C.; D’Amore, A.; Palombi, F.; Criscuolo, E.; Frasca, F.; Pierangeli, A.; Mancini, N.; Antonelli, G.; Clementi, M.; et al. Naringenin is a powerful inhibitor of SARS-CoV-2 infection in vitro. Pharmacol. Res. 2021, 163, 105255. [Google Scholar] [CrossRef]
- Cho, J.K.; Curtis-Long, M.J.; Lee, K.H.; Kim, D.W.; Ryu, H.W.; Yuk, H.J.; Park, K.H. Geranylated flavonoids displaying SARS-CoV papain-like protease inhibition from the fruits of Paulownia tomentosa. Bioorg. Med. Chem. 2013, 21, 3051–3057. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Lee, W.S. Evaluation of polyphenols from Broussonetiapapyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Seo, K.H.; Curtis-Long, M.J.; Oh, K.Y.; Oh, J.W.; Cho, J.K.; Park, K.H. Phenolic phytochemical displaying SARS-CoV papain-like protease inhibition from the seeds of Psoralea corylifolia. J. Enzym. Inhib. Med. Chem. 2014, 29, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Lee, Y.Z.; Kang, I.J.; Barnard, D.L.; Jan, J.T.; Lin, D.; Lee, S.J. Identification of phenanthroindolizines and phenanthroquinolizidines as novel potent anti-coronaviral agents for porcine enteropathogenic coronavirus transmissible gastroenteritis virus and human severe acute respiratory syndrome coronavirus. Antivir. Res. 2010, 88, 160–168. [Google Scholar] [CrossRef]
- Park, J.Y.; Kim, J.H.; Kim, Y.M.; Jeong, H.J.; Kim, D.W.; Park, K.H.; Ryu, Y.B. Tanshinones as selective and slow-binding inhibitors for SARS-CoV cysteine proteases. Bioorg. Med. Chem. 2012, 20, 5928–5935. [Google Scholar] [CrossRef]
- Zhuang, M.; Jiang, H.; Suzuki, Y.; Li, X.; Xiao, P.; Tanaka, T.; Qin, C. Procyanidins and butanol extract of Cinnamomi Cortex inhibit SARS-CoVinfection. Antivir. Res. 2009, 82, 73–81. [Google Scholar] [CrossRef]
- Wen, C.C.; Kuo, Y.H.; Jan, J.T.; Liang, P.H.; Wang, S.Y.; Liu, H.G.; Hou, C.C. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus. J. Med. Chem. 2007, 50, 4087–4095. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raddad, L.J.; Chemaitelly, H.; Butt, A.A. Effectiveness of the BNT162b2 Covid-19 Vaccine against the B. 1.1. 7 and B. 1.351 Variants. N. Engl. J. Med. 2021, 385, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Chemaitelly, H.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; Tang, P.; Hasan, M.R.; Malek, J.A.; Coyle, P.; Ayoub, H.H.; Al Kanaani, Z.; et al. mRNA-1273 COVID-19 vaccine effectiveness against the B. 1.1. 7 and B. 1.351 variants and severe COVID-19 disease in Qatar. Nat. Med. 2021, 27, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Sapkal, G.N.; Yadav, P.D.; Ella, R.; Deshpande, G.R.; Sahay, R.R.; Gupta, N.; Vadrevu, K.M.; Abraham, P.; Panda, S.; Bhargava, B. Inactivated COVID-19 vaccine BBV152/COVAXIN effectively neutralizes recently emerged B. 1.1. 7 variant of SARS-CoV-2. J. Travel Med. 2021, 28, taab051. [Google Scholar] [CrossRef]
- Shinde, V.; Bhikha, S.; Hoosain, Z.; Archary, M.; Bhorat, Q.; Fairlie, L.; Lalloo, U.; Masilela, M.S.L.; Moodley, D.; Madhi, S.A.; et al. Efficacy of NVX-CoV2373 Covid-19 vaccine against the B. 1.351 variant. N. Engl. J. Med. 2021, 384, 1899–1909. [Google Scholar] [CrossRef]
- Gushchin, V.A.; Dolzhikova, I.V.; Shchetinin, A.M.; Odintsova, A.S.; Siniavin, A.E.; Nikiforova, M.A.; Pochtovyi, A.A.; Shidlovskaya, E.V.; Kuznetsova, N.A.; Burgasova, O.A.; et al. Neutralizing activity of sera from Sputnik V-vaccinated people against variants of concern (VOC: B. 1.1. 7, B. 1.351, P. 1, B. 1.617. 2, B. 1.617. 3) and Moscow endemic SARS-CoV-2 variants. Vaccines 2021, 9, 779. [Google Scholar] [CrossRef]
- Hitchings, M.D.; Ranzani, O.T.; Torres, M.S.S.; de Oliveira, S.B.; Almiron, M.; Said, R.; Borg, R.; Schulz, W.L.; de Oliveira, R.D.; da Silva, P.V.; et al. Effectiveness of CoronaVac in the setting of high SARS-CoV-2 P. 1 variant transmission in Brazil: A test-negative case-control study. Lancet Reg. Health Am. 2021, 1, 100025. [Google Scholar]
- Clemens, S.A.C.; Folegatti, P.M.; Emary, K.R.; Weckx, L.Y.; Ratcliff, J.; Bibi, S.; De Almeida Mendes, A.V.; Milan, E.P.; Pittella, A.; Schwarzbold, A.V.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 lineages circulating in Brazil. Nat. Commun. 2021, 12, 5861. [Google Scholar] [CrossRef] [PubMed]
- Lopez Bernal, J.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of Covid-19 vaccines against the B. 1.617. 2 (Delta) variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Werner, A.P.; Koch, M.; Choi, A.; Narayanan, E.; Stewart-Jones, G.B.; Colpitts, T.; Bennett, H.; Boyoglu-Barnum, S.; Shi, W.; et al. Serum neutralizing activity elicited by mRNA-1273 vaccine. N. Engl. J. Med. 2021, 384, 1468–1470. [Google Scholar] [CrossRef] [PubMed]
- Zani, A.; Caccuri, F.; Messali, S.; Bonfanti, C.; Caruso, A. Serosurvey in BNT162b2 vaccine-elicited neutralizing antibodies against authentic B. 1, B. 1.1. 7, B. 1.351, B. 1.525 and P. 1 SARS-CoV-2 variants. Emerg. Microbes Infect. 2021, 10, 1241–1243. [Google Scholar] [CrossRef] [PubMed]
- Blanco, S.; Spinsanti, L.; Aguilar, J.J.; Diaz, A.; Rivarola, M.E.; Beranek, M.; Fernández, E.; Mangeaud, A.; Konigheim, B.S.; Gallego, S.V. Neutralizing response elicited by homologous and heterologous prime booster vaccination against ancestral SARS-CoV-2 B. 1, P. 1, C. 37 and B. 1.617. 2 variants. Vaccine 2022, 40, 6706–6710. [Google Scholar] [CrossRef]
- Tartof, S.Y.; Slezak, J.M.; Puzniak, L.; Hong, V.; Frankland, T.B.; Ackerson, B.K.; Takhar, H.; Ogun, O.A.; Simmons, S.; Zamparo, J.M.; et al. BNT162b2 vaccine effectiveness against SARS-CoV-2 omicron BA. 4 and BA. 5. Lancet Infect. Dis. 2022, 22, 1663–1665. [Google Scholar] [CrossRef]
- Huang, B.; Dai, L.; Wang, H.; Hu, Z.; Yang, X.; Tan, W.; Gao, G.F. Serum sample neutralisation of BBIBP-CorV and ZF2001 vaccines to SARS-CoV-2 501Y. V2. Lancet Microbe 2021, 2, e285. [Google Scholar] [CrossRef]


| SARS-CoV-2 Variant | Vaccines | References |
|---|---|---|
| B.1.1.7 | BNT162b2, mRNA-1273, BBV152/COVAXIN | [146,147,148] |
| B.1.351 | BNT162b2, mRNA-1273, NVX-CoV2373 | [146,147,149] |
| P.1 | Sputnik V, CoronaVac, ChAdOx1 nCoV-19 (AZD1222) | [150,151,152] |
| B.1.617.2 | Sputnik V, BNT162b2, ChAdOx1 nCoV-19 | [150,153] |
| B.1.427/B.1.429 | mRNA-1273 | [154] |
| P.2 | ChAdOx1 nCoV-19 (AZD1222) | [152] |
| B.1.525 | BNT162b2 | [155] |
| B.1.617.3 | Sputnik V | [150] |
| C.37 | Gam-COVID-Vac, ChAdOx1-S, Ad5-nCorV, BBIBP-CorV and mRNA-127 | [156] |
| B.1.1.28 | ChAdOx1 nCoV-19 (AZD1222) | [152] |
| BA.4 and BA.5 | BNT162b2 | [157] |
| 501Y.V2 | BBIBP-CorV, ZF2001 | [158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shoaib, S.; Ansari, M.A.; Kandasamy, G.; Vasudevan, R.; Hani, U.; Chauhan, W.; Alhumaidi, M.S.; Altammar, K.A.; Azmi, S.; Ahmad, W.; et al. An Attention towards the Prophylactic and Therapeutic Options of Phytochemicals for SARS-CoV-2: A Molecular Insight. Molecules 2023, 28, 795. https://doi.org/10.3390/molecules28020795
Shoaib S, Ansari MA, Kandasamy G, Vasudevan R, Hani U, Chauhan W, Alhumaidi MS, Altammar KA, Azmi S, Ahmad W, et al. An Attention towards the Prophylactic and Therapeutic Options of Phytochemicals for SARS-CoV-2: A Molecular Insight. Molecules. 2023; 28(2):795. https://doi.org/10.3390/molecules28020795
Chicago/Turabian StyleShoaib, Shoaib, Mohammad Azam Ansari, Geetha Kandasamy, Rajalakshimi Vasudevan, Umme Hani, Waseem Chauhan, Maryam S. Alhumaidi, Khadijah A. Altammar, Sarfuddin Azmi, Wasim Ahmad, and et al. 2023. "An Attention towards the Prophylactic and Therapeutic Options of Phytochemicals for SARS-CoV-2: A Molecular Insight" Molecules 28, no. 2: 795. https://doi.org/10.3390/molecules28020795
APA StyleShoaib, S., Ansari, M. A., Kandasamy, G., Vasudevan, R., Hani, U., Chauhan, W., Alhumaidi, M. S., Altammar, K. A., Azmi, S., Ahmad, W., Wahab, S., & Islam, N. (2023). An Attention towards the Prophylactic and Therapeutic Options of Phytochemicals for SARS-CoV-2: A Molecular Insight. Molecules, 28(2), 795. https://doi.org/10.3390/molecules28020795