

Investigation of NO Role in Neural Tissue in Brain and Spinal Cord Injury

Abstract
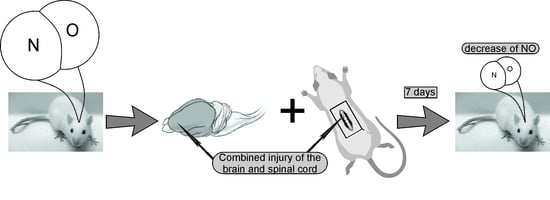
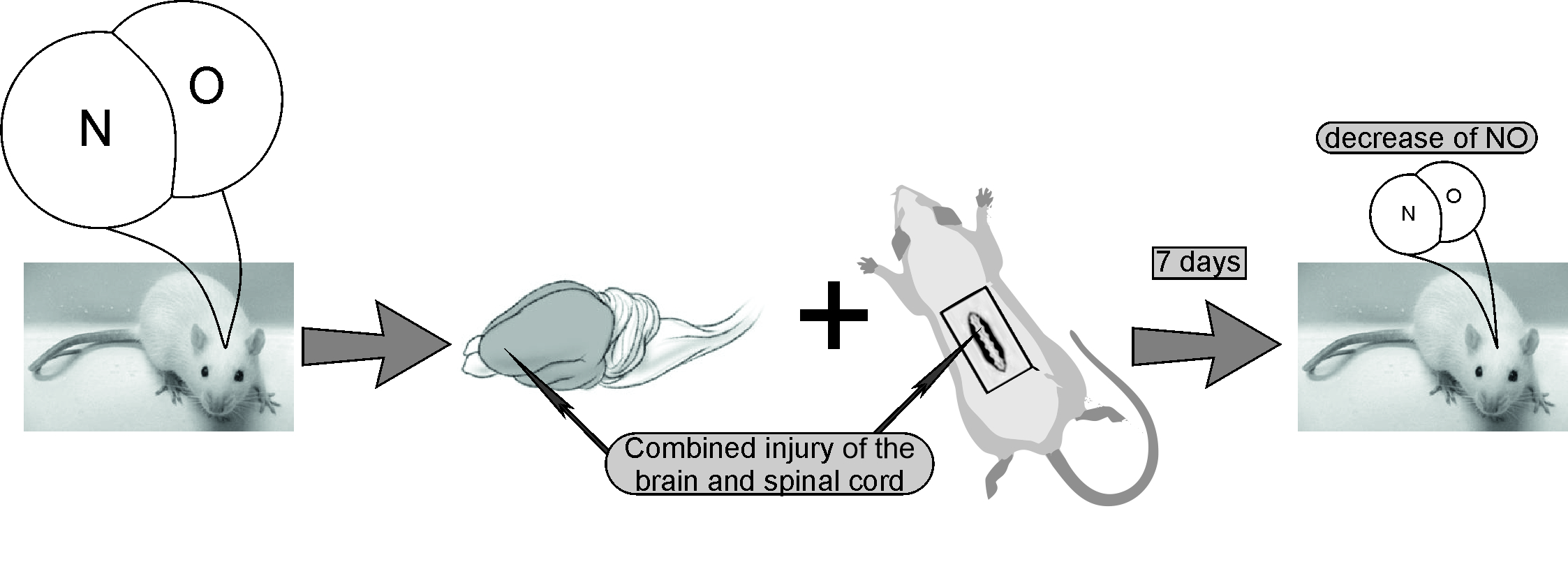
:
{kind=link}
{kind=link}
{kind=link}
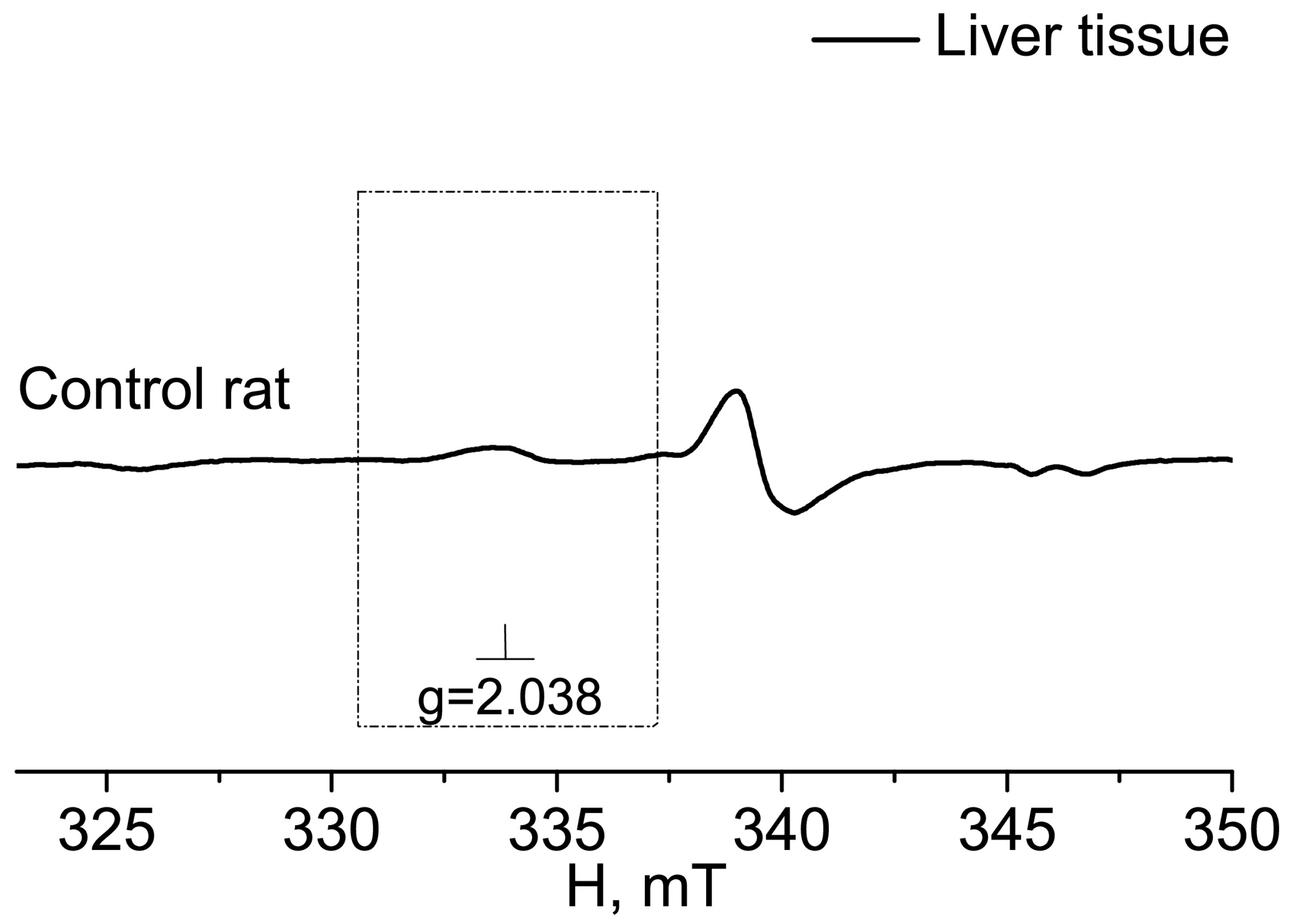{kind=link}
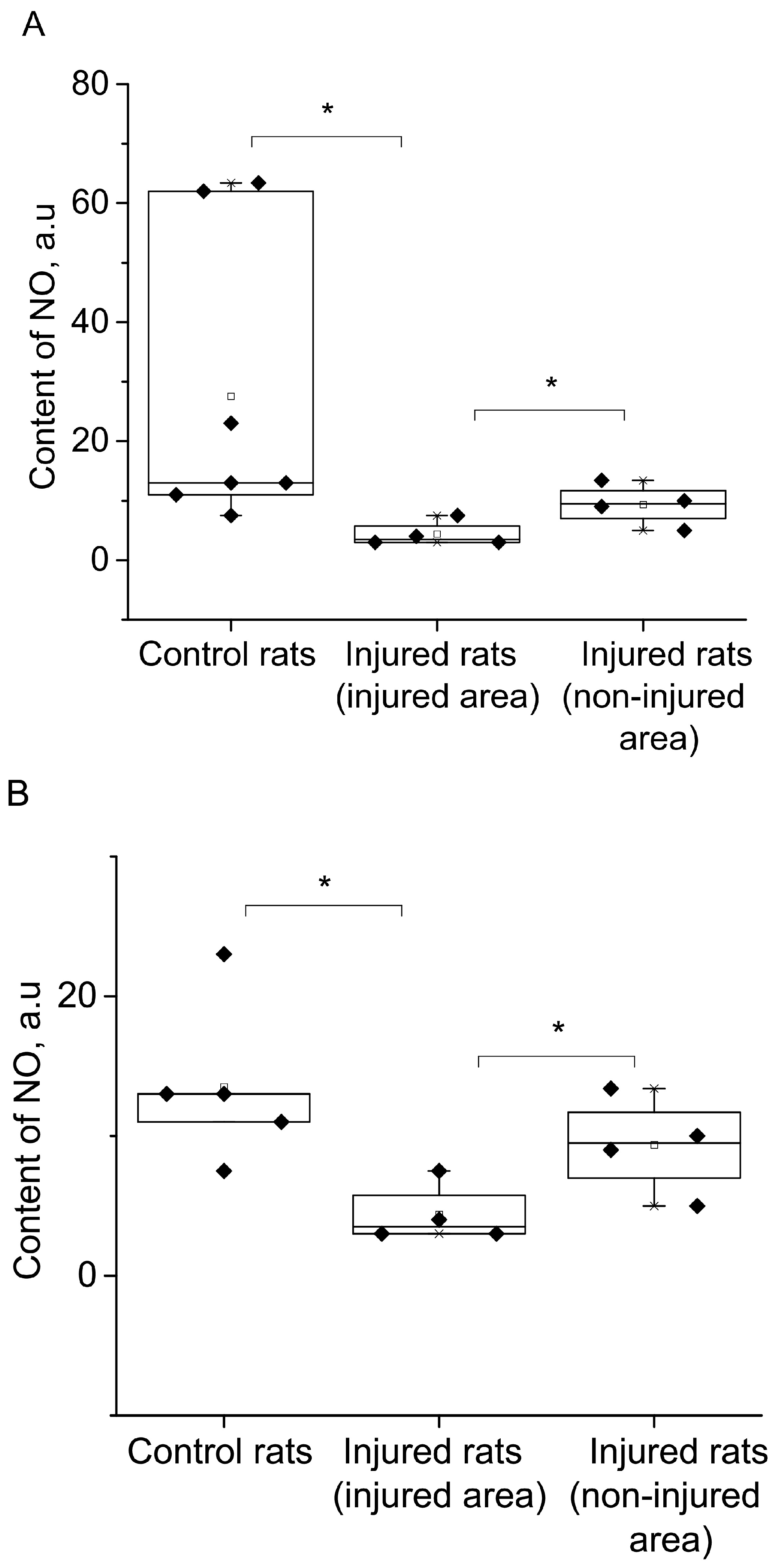{kind=link}
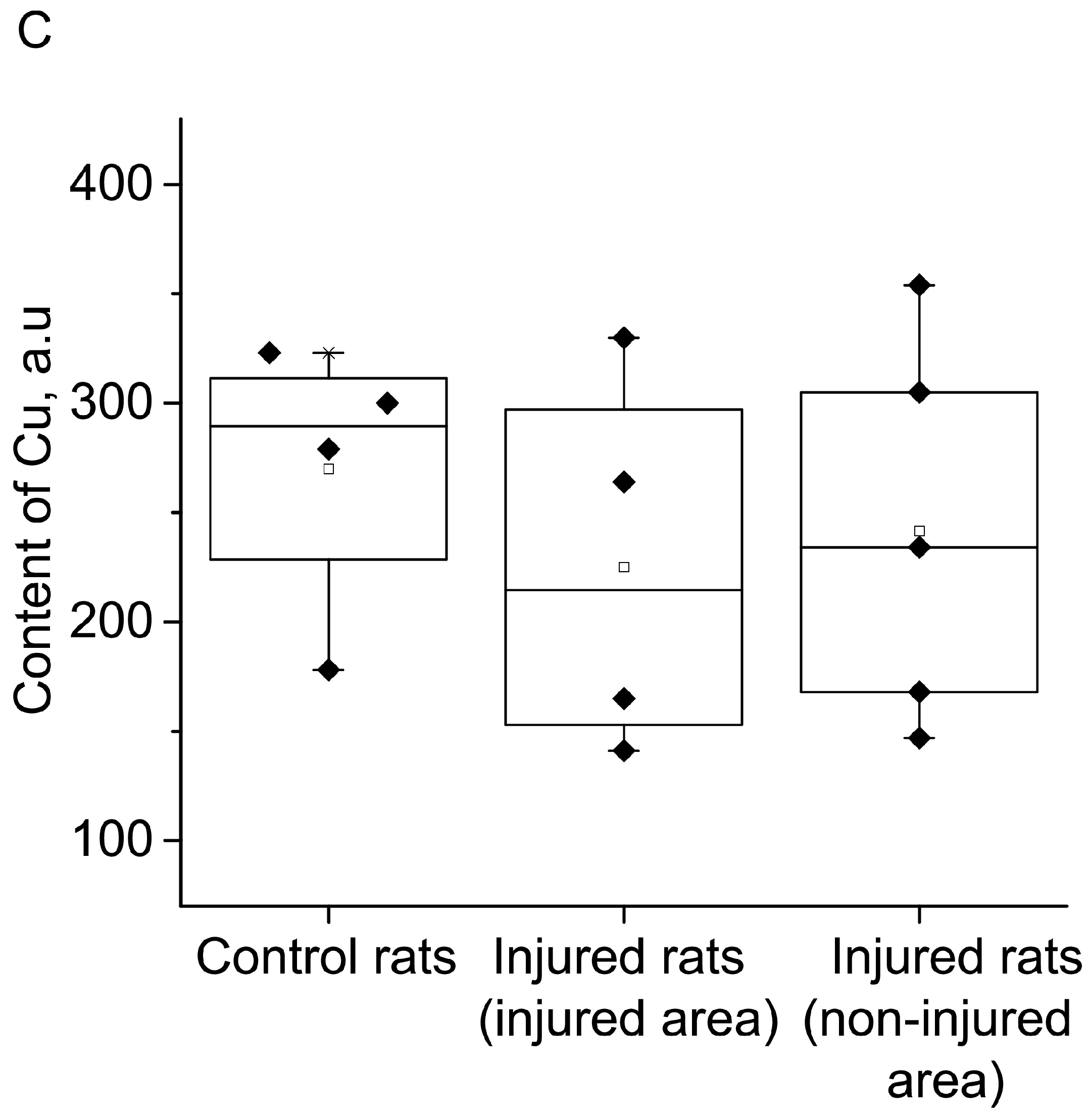{kind=link}
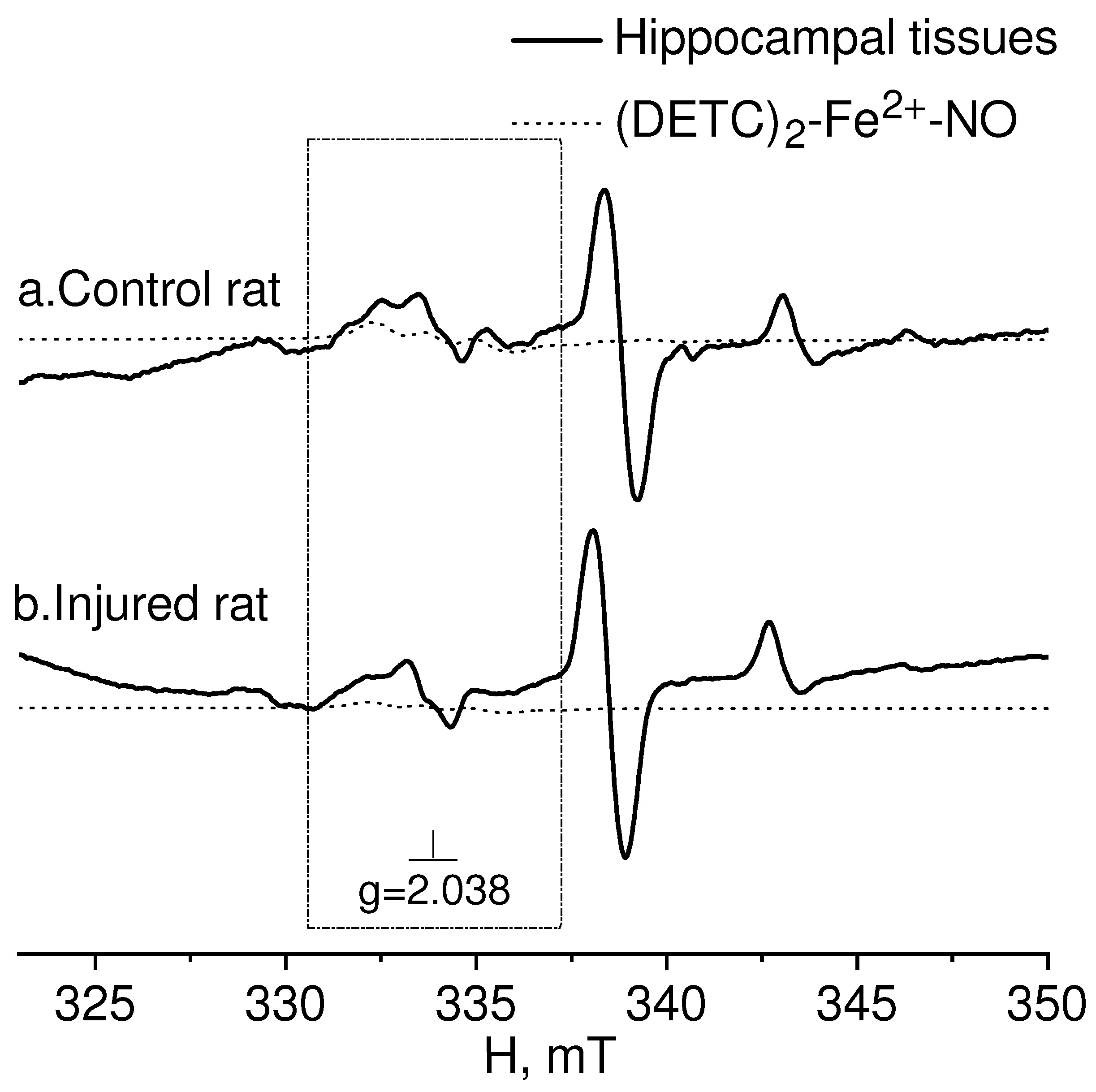{kind=link}
{kind=link}
1. Introduction
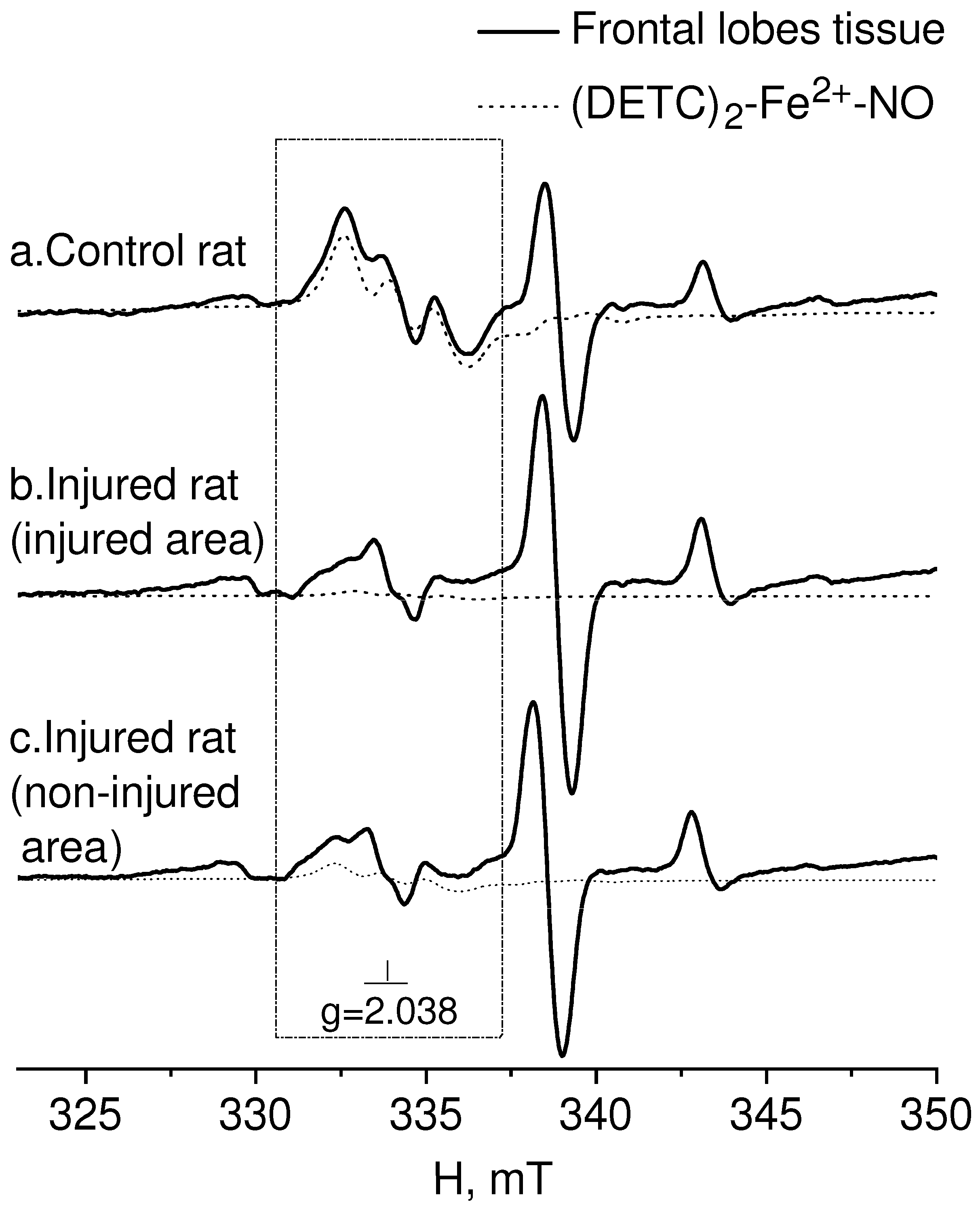
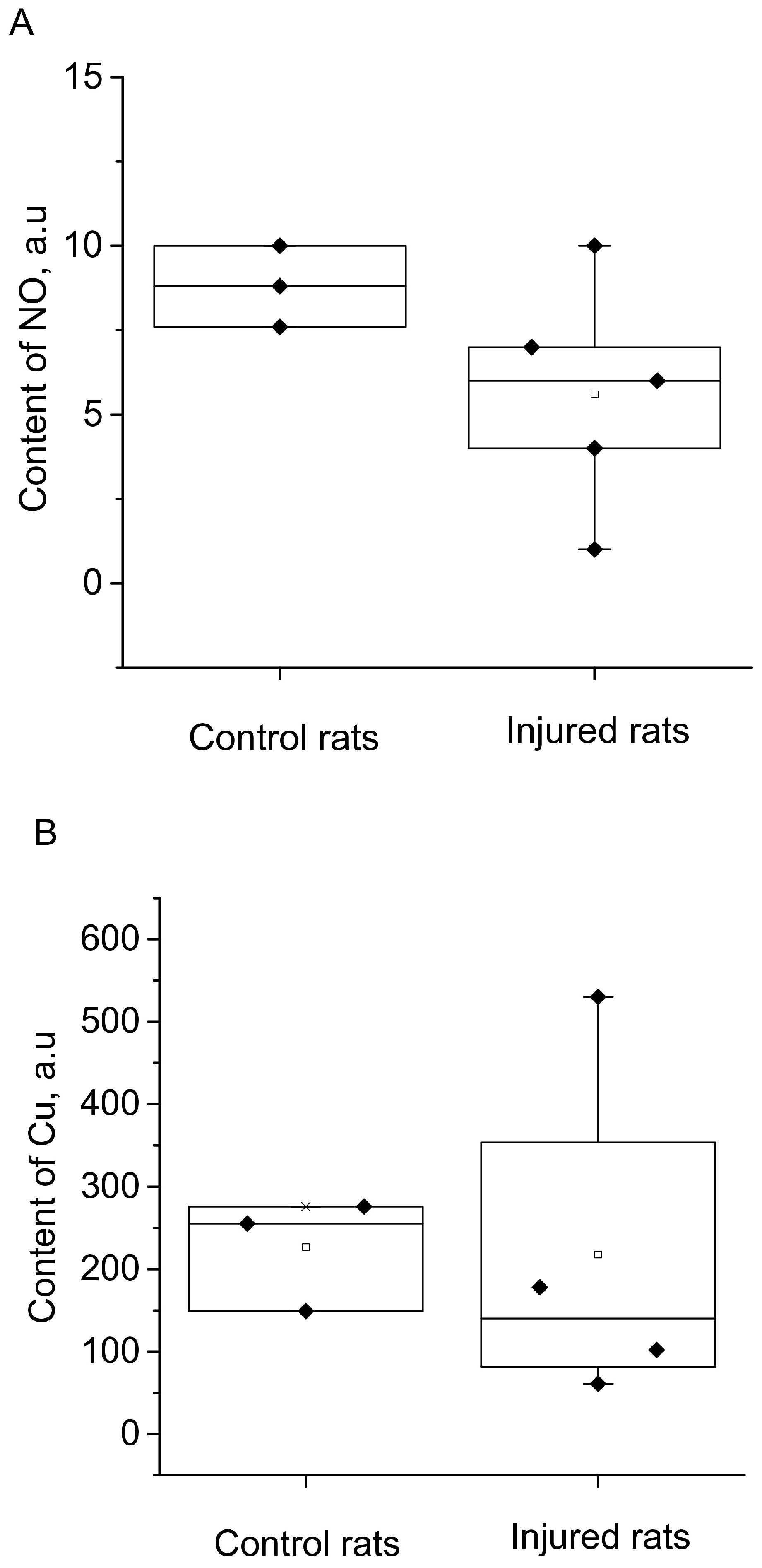
2. Results
3. Discussion
4. Experimental Procedures
4.1. Animals
4.2. Experiment Protocol: Modeling of Combined Trauma of the Brain and Spinal Cord in Rats
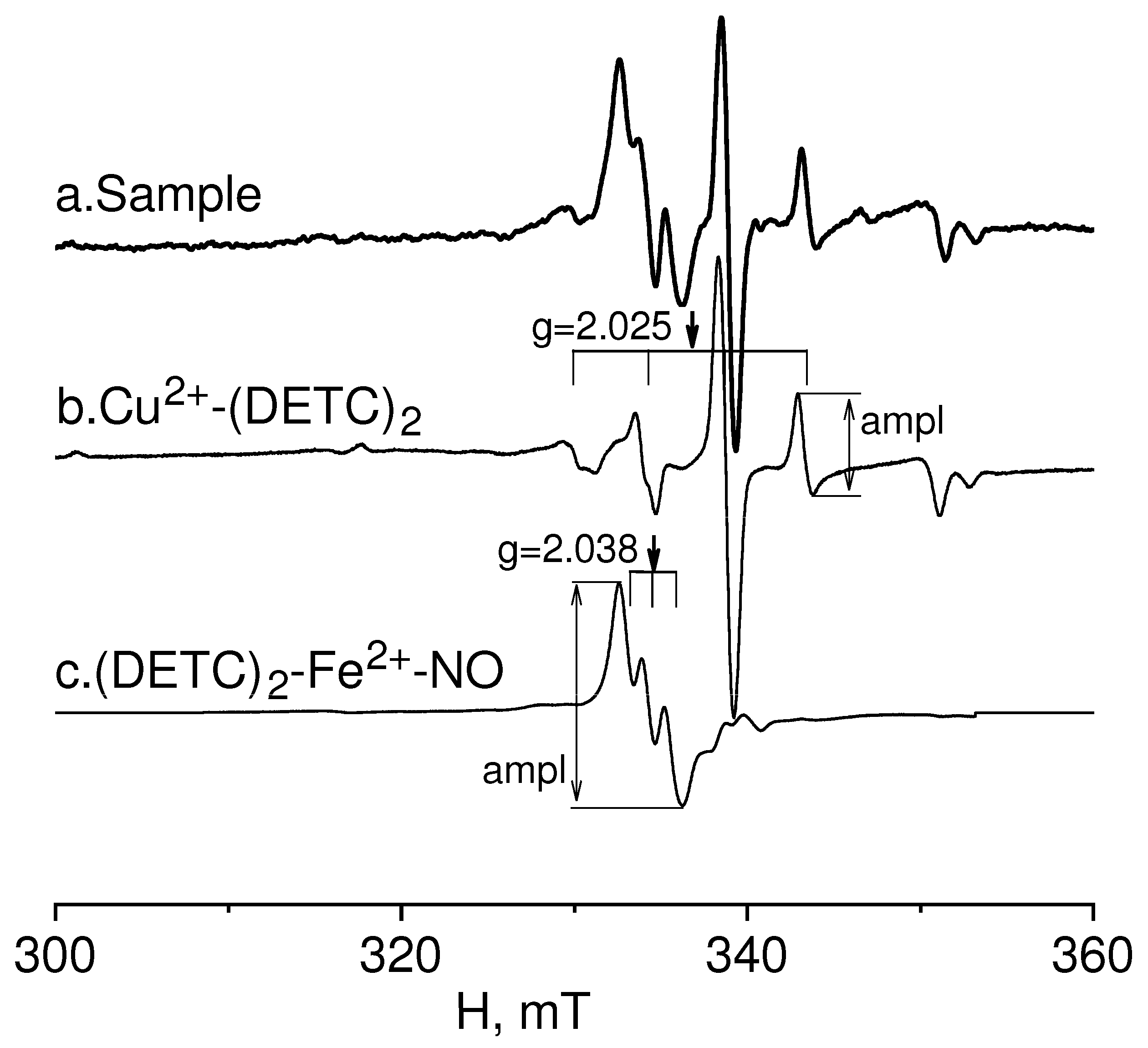
4.3. Formation of a (DETC)2-Fe2+-NO Complex with a Spin Trap in Rat Tissues
4.4. Measurements of a (DETC)2-Fe2+-NO and Cu(DETC)2 Complexes in Rats Tissues
4.5. Statistical Processing of the Result
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garthwaite, J. Concepts of neural nitric oxide-mediated transmission. Eur. J. Neurosci. 2008, 27, 2783–2802. [Google Scholar] [CrossRef] [PubMed]
- Steinert, J.R.; Chernova, T.; Forsythe, I.D. Nitric oxide signaling in brain function, dysfunction, and dementia. Neuroscientist 2010, 16, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.M.; Lourenco, C.F.; Ledo, A.; Barbosa, R.M.; Laranjinha, J. Nitric oxide inactivation mechanisms in the brain: Role in bioenergetics and neurodegeneration. Intern. J. Cell Biol. 2012, 2012, 391914. [Google Scholar] [CrossRef]
- Maggio, D.M.; Singh, A.; Iorgulescu, J.B.; Bleicher, D.H.; Ghosh, M.; Lopez, M.M.; Tuesta, L.M.; Flora, G.; Dietrich, W.D.; Pearse, D.D. Identifying the long-term role of inducible nitric oxide synthase after contusive spinal cord injury using a transgenic mouse model. Int. J. Mol. Sci. 2017, 18, 245. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Weitzberg, E. Nitric oxide signaling in health and disease. Cell 2022, 185, 2853–2878. [Google Scholar] [CrossRef] [PubMed]
- Boehning, D.; Snyder, S.H. Novel neural modulators. Annu. Rev. Neurosci. 2003, 26, 105–131. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Rizzarelli, E.; Owen, J.B.; Dinkova-Kostova, A.T.; Butterfield, D.A. Nitric oxide in cell survival: A janus molecule. Antioxid. Redox Signal. 2009, 11, 2717–2739. [Google Scholar] [CrossRef]
- Balaban, P.M.; Roshchin, M.V.; Timoshenko, A.K.; Gainutdinov, K.L.; Bogodvid, T.K.; Muranova, L.N.; Zuzina, A.B.; Korshunova, T.A. Nitric oxide is necessary for labilization of a consolidated context memory during reconsolidation in terrestrial snails. Eur. J. Neurosci. 2014, 40, 2963–2970. [Google Scholar] [CrossRef]
- Moroz, L.L. Gaseous transmission across time and species. Am. Zool. 2015, 41, 304–320. [Google Scholar] [CrossRef]
- Reutov, V.P.; Okhotin, V.E.; Shuklin, A.V.; Sorokina, E.G.; Kosicin, N.S.; Gurin, V.N. Nitric oxide and the cycle in the myocardium: Molecular, biochemical and physiological aspects. Usp. Fiziol. Nauk. 2007, 38, 39–58. (In Russian) [Google Scholar]
- Andrianov, V.V.; Sitdikov, F.G.; Gainutdinov, K.L.; Yurtaeva, S.V.; Muranova, L.N.; Obynochnyi, A.A.; Karimov, F.K.; Chiglintsev, V.M.; Iyudin, V.S. Changes in nitric oxide in heart of intact and sympathectomized rats of different age. Russ. J. Dev. Biol. 2008, 38, 352–356. [Google Scholar] [CrossRef]
- Baider, L.M.; Reutov, V.P.; Krushinsky, A.L.; Kuzenkov, V.S.; Sorokina, E.G.; Koshelev, V.B.; Fadyukova, O.E.; Djumbaeva, T.T.; Komissarova, L.K.; Pinelis, V.G.; et al. Investigation by method of EPR of influence of hypoxia on nitric oxide (NO) production in blood of rats Krushinskii-Molodkina. Biofizika 2009, 54, 894–899. (In Russian) [Google Scholar] [PubMed]
- Tennyson, A.G.; Lippard, S.J. Generation, translocation, and action of nitric oxide in living systems. Chem. Biol. 2011, 18, 1211–1220. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Zhao, F.; Wang, H.; Qu, Y.; Mu, D. Nitric oxide synthase in hypoxic or ischemic brain injury. Rev. Neurosci. 2015, 26, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Garry, P.S.; Ezra, M.; Rowland, M.J.; Westbrook, J.; Pattinson, K.T. The role of the nitric oxide pathway in brain injury and its treatment—From bench to bedside. Exp. Neurol. 2015, 263, 235–243. [Google Scholar] [CrossRef]
- Sukmanskiy, O.I.; Reutov, V.P. Gasotransmitters: Physiological role and involvement in the pathogenesis of diseases. Uspekhi Fiziol. Nauk. 2016, 47, 30–58. (In Russian) [Google Scholar]
- Sobrevia, L.; Ooi, L.; Ryan, S.; Steinert, J.R. Nitric oxide: A regulator of cellular function in health and disease. Oxidative Med. Cell. Longev. 2016, 2016, 9782346. [Google Scholar] [CrossRef]
- Balez, R.; Ooi, L. Getting to NO Alzheimers Disease: Neuroprotection versus neurotoxicity mediated by nitric oxide. Oxidative Med. Cell. Longev. 2016, 2016, 3806157. [Google Scholar] [CrossRef]
- Dubey, H.; Gulati, K.; Ray, A. Effects of Nitric Oxide (NO) Modulators on cognitive function and brain oxidative stress in experimental model of Alzheimers disease in rats. J. Pharmacol. Rep. 2017, 2, 2. [Google Scholar]
- Vanin, A.F. Dinitrosyl iron complexes and S-nitrosothiols are two possible forms of stabilization and transport of nitric oxide in biosystems. Biohimia 1998, 63, 924–938. (In Russian) [Google Scholar]
- Ignarro, L.J.; Cirino, G.; Casini, A.; Napoli, C. Nitric oxide as a signaling molecule in the vascular system: An overview. J. Cardiovasc. Pharmacol. 1999, 34, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Manukhina, E.B.; Malyshev, I.Y. Stress-limiting nitric oxide system. Ross. Fiziol. Zhurnal Im. I.M. Sechenova 2000, 86, 1283–1292. (In Russian) [Google Scholar]
- Luiking, Y.C.; Engelen, M.P.K.J.; Deutz, N.E.P. Regulation of nitric oxide production in health and disease. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, T.A.; da Silva, R.S.; Miranda, K.M.; Switzer, C.H.; Wink, D.A.; Fukuto, J.M. Biological nitric oxide signalling: Chemistry and terminology. Br. J. Pharmacol. 2013, 169, 1417–1429. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–427. [Google Scholar] [CrossRef]
- Serrano, J.; Fernández, A.P.; Martínez-Murillo, R.; Alonso, D.; Rodrigo, J.; Salas, E.; Mourelle, M.; Martínez, A. The nitric oxide donor LA 419 decreases ischemic brain damage. Int. J. Mol. Med. 2007, 19, 229–236. [Google Scholar] [CrossRef]
- Wierónska, J.M.; Cieślik, P.; Kalinowski, L. Nitric oxide-dependent pathways as critical factors in the consequences and recovery after brain ischemic hypoxia. Biomolecules 2021, 11, 1097. [Google Scholar] [CrossRef]
- Godecke, A.; Schrader, J. The Janus faces of NO. Circ. Res. 2004, 94, e55–e57. [Google Scholar] [CrossRef]
- Calabrese, V.; Mancuso, C.; Calvani, M.; Rizzarelli, E.; Butterfield, D.A.; Stella, A.M.G. Nitric oxide in the central nervous system: Neuroprotection versus neurotoxicity. Nat. Rev. Neurosci. 2007, 8, 767–775. [Google Scholar] [CrossRef]
- Vanin, A.F. What is the mechanism of nitric oxide conversion into nitrosonium ions ensuring S-nitrosating processes in living organisms. Cell Biochem. Biophys. 2019, 77, 279–292. [Google Scholar] [CrossRef]
- Samdani, A.F.; Dawson, T.M.; Dawson, V.L. Nitricoxide synthase in models of focal ischemia. Stroke 1997, 28, 1283. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C.; Zhang, F.; Casey, R.; Nagayama, M.; Ross, M.E. Delayed reduction of ischemic brain injury and neurological deficits in mice lacking the inducible nitric oxide synthase gene. J. Neurosci. 1997, 17, 9157–9164. [Google Scholar] [CrossRef] [PubMed]
- Malyshev, I.Y.; Zenina, T.A.; Golubeva, L.Y.; Saltykova, V.A.; Manukhina, E.B.; Mikoyan, V.D.; Kubrina, L.N.; Vanin, A.F. NO-dependent mechanisms of adaptation to hypoxia. Nitric Oxide 1999, 3, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Tominaga, T.; Ohnishi, T.; Ohnishi, S.T. Electron paramagnetic resonance study on nitric oxide production during brain focal ischemia and reperfusion in the rat. Brain Res. 1994, 647, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, T.; Sato, S.; Ohnishi, T.; Ohnishi, S.T. Electron paramagnetic resonance (EPR) detection of nitric oxide produced during forebrain ischemia of the rat. J. Cereb. Blood Flow Metab. 1994, 14, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Irie, Y.; Keung, W.M.; Maret, W. S-nitrosothiols react preferentially with zinc thiolate clasters of metallothionein III through transnitrosation. Biochemistry 2002, 41, 8360–8367. [Google Scholar] [CrossRef] [PubMed]
- Dawson, D.A.; Kusumoto, K.; Graham, D.I.; McCulloch, J.; Macrae, I.M. Inhibition of nitric oxide synthesis does not reduce infarct volume in a rat model of focal cerebral ischaemia. Neurosci. Lett. 1992, 142, 151–154. [Google Scholar] [CrossRef]
- Sancesario, G.; Iannone, M.; Morello, M.; Nistico, G.; Bernardi, G. Nitric-oxide inhibition aggravates ischemic damage of hippocampal but not of NADPH neurons in gerbils. Stroke 1994, 25, 436–443. [Google Scholar] [CrossRef]
- Yamamoto, S.; Golanov, E.V.; Berger, S.B.; Reis, D.J. Inhibition of nitric oxide synthesis increases focal ischemic infarction in rat. J. Cereb. Blood Flow Metab. 1992, 12, 717–726. [Google Scholar] [CrossRef]
- Willmot, M.; Gray, L.; Gibson, C.; Murphy, S.; Bath, P.M. A systematic review of nitric oxide donors and L-arginine in experimental stroke; effects on infarct size and cerebral blood flow. Nitric Oxide 2005, 12, 141–149. [Google Scholar] [CrossRef]
- Jung, K.H.; Chu, K.; Ko, S.Y.; Lee, S.T.; Sinn, D.I.; Park, D.K.; Kim, J.M.; Song, E.C.; Kim, M.; Roh, J.K. Early intravenous infusion of sodium nitrite protects brain against in vivo ischemia-reperfusion injury. Stroke 2006, 37, 2744–2750. [Google Scholar] [CrossRef] [PubMed]
- Evgenov, O.V.; Pacher, P.; Schmidt, P.M.; Haskó, G.; Schmidt, H.H.H.W.; Stasch, J.-P. NO-independent stimulators and activators of soluble guanylate cyclase: Discovery and therapeutic potential. Nat. Rev. Drug Discov. 2006, 5, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Godinez-Rubi, M.; Rojas-Mayorquin, A.E.; Ortuno-Sahagun, D. Nitric oxide donors as neuroprotective agents after an ischemic stroke-related inflammatory reaction. Oxid. Med. Cell. Longev. 2013, 2013, 297357. [Google Scholar] [CrossRef]
- Kuzenkov, V.S.; Krushinskiy, A.L. Protective effect of magnesium nitrate on cerebral ischemia. Vestn. Mosk. Univ. Biol. 2014, 4, 9–14. (In Russian) [Google Scholar]
- Wang, Y.Z.; Li, T.T.; Cao, H.L.; Yang, W.C. Recent advances in the neuroprotective effects of medical gases. Med. Gas Res. 2019, 9, 80–87. [Google Scholar] [CrossRef]
- Dou, C.; Han, X.; Hanbin, X.; Liao, H.; Xiao, X.; Huang, Z.; Luo, G.; Zhang, X.; Yao, W. Protective role of nitric oxide donors on endothelium in ischemia-reperfusion injury: A meta-analysis of randomized controlled trials. BMC Anesthesiol. 2023, 23, 189. [Google Scholar] [CrossRef]
- Jung, P.; Ha, E.; Zhang, M.; Fall, C.; Hwang, M.; Taylor, E.; Stetkevich, S.; Bhanot, A.; Wilson, C.G.; Figueroa, J.D.; et al. Neuroprotective role of nitric oxide inhalation and nitrite in a neonatal rat model of hypoxic-ischemic injury. PLoS ONE 2022, 17, e0268282. [Google Scholar] [CrossRef]
- Gainutdinov, K.L.; Gavrilova, S.A.; Iyudin, V.S.; Golubeva, A.V.; Davydova, M.P.; Jafarova, G.G.; Andrianov, V.V.; Koshelev, V.B. EPR study of the intensity of the nitric oxide production in rat brain after ischemic stroke. Appl. Magn. Reson. 2011, 40, 267–278. [Google Scholar] [CrossRef]
- Andrianov, V.V.; Pashkevich, S.G.; Yafarova, G.G.; Denisov, A.A.; Iyudin, V.S.; Bogodvid, T.K.; Dosina, M.O.; Kulchitsky, V.A.; Gainutdinov, K.L. Changes of nitric oxide content in the rat hippocampus, heart and liver in acute phase of ischemia. Appl. Magn. Reson. 2016, 47, 965–976. [Google Scholar] [CrossRef]
- Andrianov, V.V.; Yafarova, G.G.; Pashkevich, S.G.; Tokalchik, Y.P.; Dosina, M.O.; Zamaro, A.S.; Bogodvid, T.K.; Iyudin, V.S.; Bazan, L.V.; Denisov, A.A.; et al. Changes of the nitric oxide and copper content in the olfactory bulbs of rats brain after modeling of brain stroke and intranasal administration of mesenchymal stem cells. Appl. Magn. Reson. 2020, 51, 375–387. [Google Scholar] [CrossRef]
- Deryagin, O.G.; Gavrilova, S.A.; Buravkov, S.V.; Andrianov, V.V.; Yafarova, G.G.; Gainutdinov, K.L.; Koshelev, V.B. The role of ATP-sensitive potassium channels and nitric oxide in the protective effect of preconditioning of the brain. Neurosci. Behav. Physiol. 2018, 48, 58–63. [Google Scholar] [CrossRef]
- Deryagin, O.G.; Gavrilova, S.A.; Gainutdinov, K.L.; Golubeva, A.V.; Andrianov, V.V.; Yafarova, G.G.; Buravkov, S.V.; Koshelev, V.B. Molecular bases of brain preconditioning. Front. Neurosci. 2017, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, K.D.; Devarakonda, C.B.; Joshi, A.R.; Sharma, S.S.; Roy, N. Role of nitric oxide synthases in cerebral ischemia. Curr. Res. Inf. Pharmaceut. Sci. 2011, 11, 50–56. [Google Scholar]
- Chen, Z.Q.; Mou, R.T.; Feng, D.X.; Wang, Z.; Chen, G. The role of nitric oxide in stroke. Med. Gas Res. 2017, 7, 194–203. [Google Scholar] [PubMed]
- Wang, Y.; Hong, F.; Yang, S. Role of nitric oxide in brain ischemia and reperfusion. Int. J. Mol. Sci. 2022, 23, 4243. [Google Scholar] [CrossRef]
- Zhang, J.; Han, Y.; Wang, Y.; Cheng, X.; Wang, C.-J. Neuronal nitric oxide synthase inhibition reduces brain damage by promoting collateral recruitment in a cerebral hypoxia-ischemia mice model. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3166–3172. [Google Scholar] [CrossRef]
- Charriaut-Marlangue, C.; Bonnin, P.; Gharib, A.; Leger, P.-L.; Villapol, S.; Pocard, M.; Gressens, P.; Renolleau, S.; Baud, O. Inhaled nitric oxide reduces brain damage by collateral recruitment in a neonatal stroke model. Stroke 2012, 43, 3078–3084. [Google Scholar] [CrossRef]
- Terpolilli, N.A.; Kim, S.-W.; Thal, S.C.; Kuebler, W.M.; Plesnila, N. Inhaled nitric oxide reduces secondary brain damage after traumatic brain injury in mice. J. Cereb. Blood Flow Metab. 2013, 33, 311–318. [Google Scholar] [CrossRef]
- Terpolilli, N.A.; Feiler, S.; Dienel, A.; Müller, F.; Heumos, N.; Friedrich, B.; Stover, J.; Thal, S.; Schöller, K.; Plesnila, N. Nitric oxide inhalation reduces brain damage, prevents mortality, and improves neurological outcome after subarachnoid hemorrhage by resolving early pial microvasospasms. J. Cereb. Blood Flow Metab. 2016, 36, 2096–2107. [Google Scholar] [CrossRef]
- Remizova, M.I.; Kochetygov, N.I.; Gerbout, K.A.; Lakomkin, V.L.; Timoshin, A.A.; Burgova, E.N.; Vanin, A.F. Effect of dinitrosyl iron complexes with glutathione on hemorrhagic shock followed by saline treatment. Eur. J. Pharmacol. 2011, 662, 40–46. [Google Scholar] [CrossRef]
- Terpolilli, N.A.; Moskowitz, M.A.; Plesnila, N. Nitric oxide: Considerations for the treatment of ischemic stroke. J. Cereb. Blood Flow Metab. 2012, 32, 1332–1346. [Google Scholar] [CrossRef] [PubMed]
- Manukhina, E.B.; Malyshev, I.Y.; Smirin, B.V.; Mashina, S.Y.; Saltykova, V.A.; Vanin, A.F. Production and storage of nitric oxide in adaptation to hypoxia. Nitric Oxide 1999, 3, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Bolanos, J.P.; Almeida, A. Roles of nitric oxide in brain hypoxia-ischemia. Biochim. Biophys. Acta 1999, 1411, 415–436. [Google Scholar] [CrossRef]
- Capizzi, A.; Woo, J.; Verduzco-Gutierrez, M. Traumatic brain injury: An overview of epidemiology, pathophysiology, and medical management. Med. Clin. N. Am. 2020, 104, 213–238. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, Y.E.V.; Gavrilova, S.A.; Koshelev, V.B. Mechanisms of development of acute ischemic brain injury: Clinical and experimental possibilities of its correction. Reg. Circ. Microcirc. 2021, 20, 5–19. (In Russian) [Google Scholar] [CrossRef]
- Jia, Z.; Zhu, H.; Li, J.; Wang, X.; Misra, H.; Li, Y. Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord 2012, 50, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Kozyreva, T.; Zovo, K.; Palumaa, P. Affinity gradients drive copper to cellular destinations. Nature 2010, 465, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef]
- Miller, A.-F. Superoxide dismutases: Ancient enzymes and new insights. FEBS Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.-F.; Teixeira, M.; Valentine, J.S. Superoxide dismutases and superoxide reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef]
- Sheng, Y.; Capri, J.; Waring, A.; Valentine, J.S.; Whitelegge, J. Exposure of solvent-inaccessible regions in the amyloidogenic protein human SOD1 determined by hydroxyl radical footprinting. J. Am. Soc. Mass Spectrom. 2019, 30, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed]
- Sehba, F.A.; Bederson, J.B. Mechanisms of acute brain injury after subarachnoid hemorrhage. Neurol. Res. 2006, 28, 381–398. [Google Scholar] [CrossRef]
- Cook, A.M.; Jones, G.M.; Hawryluk, G.W.J.; Mailloux, P.; McLaughlin, D.; Papangelou, A.; Samuel, S.; Tokumaru, S.; Venkatasubramanian, C.; Zacko, C.; et al. Guidelines for the acute treatment of cerebral edema in neurocritical care patients. Neurocrit. Care 2020, 32, 647–666. [Google Scholar] [CrossRef]
- Everitt, A.; Root, B.; Calnan, D.; Manwaring, P.; Bauer, D.; Halter, R. A bioimpedance-based monitor for real-time detection and identification of secondary brain injury. Sci. Rep. 2021, 11, 15454. [Google Scholar] [CrossRef] [PubMed]
- Murthy, T.; Bhatia, P.; Sandhu, K.; Prabhakar, T.; Gogna, R. Secondary brain injury: Prevention and intensive care management. Indian J. Neurotrauma 2005, 2, 7–12. [Google Scholar] [CrossRef]
- Che, X.; Fang, Y.; Si, X.; Wang, J.; Hu, X.; Reis, C.; Chen, S. The role of gaseous molecules in traumatic brain injury: An updated review. Front. Neurosci. 2018, 12, 392. [Google Scholar] [CrossRef]
- Andrianov, V.V.; Kulchitsky, V.A.; Yafarova, G.G.; Zamaro, A.S.; Tokalchik, Y.P.; Bazan, L.V.; Bogodvid, T.K.; Iyudin, V.S.; Pashkevich, S.G.; Dosina, M.O.; et al. Comparative study of the intensity of nitric oxide production and copper content in the hippocampus of rats after modeling hemorrhagic stroke and brain injury. Appl. Magn. Reson. 2021, 52, 1657–1669. [Google Scholar] [CrossRef]
- Crobeddu, E.; Pilloni, G.; Tardivo, V.; Fontanella, M.M.; Panciani, P.P.; Spena, G.; Fornaro, R.; Altieri, R.; Agnoletti, A.; Ajello, M.; et al. Role of nitric oxide and mechanisms involved in cerebral injury after subarachnoid hemorrhage: Is nitric oxide a possible answer to cerebral vasospasm? J. Neurosurg. Sci. 2016, 60, 385–391. [Google Scholar]
- Mikoyan, V.D.; Kubrina, L.N.; Serezhenkov, V.A.; Stukan, R.A.; Vanin, A.F. Complexes of Fe2+ with diethyldithiocarbamate or N-methyl-D-glucamine dithiocarbamate as traps of nitric oxide in animal tissues. Biochim. Biophys. Acta 1997, 1336, 225–234. [Google Scholar] [CrossRef]
- Van Faassen, E.E.; Koeners, M.P.; Joles, J.A.; Vanin, A.F. Detection of basal NO production in rat tissues using iron dithiocarbamate complexes. Nitric Oxide 2008, 18, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Jakubowska, M.A.; Pyka, J.; Michalczyk-Wetula, D.; Baczyński, K.; Cieśla, M.; Susz, A.; Ferdek, P.E.; Płonka, B.K.; Fiedor, L.; Płonka, P.M. Electron paramagnetic resonance spectroscopy reveals alterations in the redox state of endogenous copper and iron complexes in photodynamic stress-induced ischemic mouse liver. Redox Biol. 2020, 34, 101566. [Google Scholar] [CrossRef] [PubMed]
- Gainutdinov, K.L.; Kulchitsky, V.A.; Andrianov, V.V.; Yafarova, G.G.; Tokalchik, Y.P.; Zamaro, A.S.; Bazan, L.V.; Iyudin, V.S.; Bogodvid, T.K.; Pashkevich, S.G.; et al. Application of EPR spectroscopy to study the content of NO and copper in the frontal lobes, hippocampus and liver of rats after brain ischemia. Tech. Phys. 2022, 67, 311–316. [Google Scholar] [CrossRef]
- Ismailova, A.I.; Gnezdilov, O.I.; Muranova, L.N.; Obynochny, A.A.; Andrianov, V.V.; Gainutdinov, K.L.; Nasyrova, A.G.; Nigmatullina, R.R.; Rahmatullina, F.F.; Zefirov, A.L. ESR study of the nitric oxide production in tissues of animals under the external influence on the functioning of the cardiovascular and nervous systems. Appl. Magn. Reson. 2005, 28, 421–430. [Google Scholar] [CrossRef]
- AlRuwaili, R.; Al-Kuraishy, H.M.; Alruwaili, M.; Khalifa, A.K.; Alexiou, A.; Papadakis, M.; Saad, H.M.; Batiha, G. E-S. The potential therapeutic effect of phosphodiesterase 5 inhibitors in the acute ischemic stroke (AIS). Mol. Cell. Biochem. 2023, 3, 1–12. [Google Scholar] [CrossRef]
- Vanin, A.F. Dinitrosyl iron complexes with thiol-containing ligands as a working form of endogenous nitric oxide. Nitric Oxide 2016, 54, 15–29. [Google Scholar] [CrossRef]
- Maiese, K. The dynamics of cellular injury: Transformation into neuronal and vascular protection. Histol. Histopathol. 2001, 16, 633–644. [Google Scholar] [CrossRef]
- Guo, Z.-N.; Shao, A.; Tong, L.-S.; Sun, W.; Liu, J.; Yang, Y. The role of nitric oxide and sympathetic control in cerebral autoregulation in the setting of subarachnoid hemorrhage and traumatic brain injury. Mol. Neurobiol. 2016, 53, 3606–3615. [Google Scholar] [CrossRef]
- Donnan, G.A.; Fisher, M.; Macieod, M.; Davis, S.M. Stroke. Lancet 2008, 371, 1612–1623. [Google Scholar] [CrossRef]
- Reutov, V.P.; Samosudova, N.V.; Sorokina, E.G. A model of glutamate neurotoxicity and mechanisms of the development of the typical pathological process. Biophysics 2019, 64, 233–250. [Google Scholar] [CrossRef]
- Zhang, R.L.; Zhang, Z.G.; Chopp, M. Targeting nitric oxide in the subacute restorative treatment of ischemic stroke. Expert Opin. Investig. Drugs 2013, 22, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Vanin, A.F.; Huisman, A.; Van Faassen, E.E. Iron dithiocarbamate as spin trap for nitric oxide detection: Methods in Enzymology. Pitfalls Successes 2003, 359, 27–42. [Google Scholar]
- Hogg, N. Detection of nitric oxide by electron paramagnetic resonance spectroscopy. Free. Radic. Biol. Med. 2010, 49, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Vanin, A.F.; Mordvintcev, P.I.; Kleshchev, A.L. Appearance of nitrogen oxide in animal tissues in vivo. Stud. Biophys. 1984, 102, 135–143. [Google Scholar]
- Shanko, Y.; Zamaro, A.; Takalchik, S.Y.; Koulchitsky, S.; Pashkevich, S.; Panahova, E.; Navitskaya, V.; Dosina, M.; Denisov, A.; Bushuk, S.; et al. Mechanisms of neural network structures recovery in brain trauma. Biomed. J. Sci. Tech. Res. 2018, 7, 001567. [Google Scholar]
- Bogodvid, T.; Pashkevich, S.; Dosina, M.; Zamaro, A.; Takalchik, Y.; Yafarova, G.; Andrianov, V.; Denisov, A.; Loiko, D.; Gainutdinov, K.; et al. Effect of intranasal administration of mesenchymal stem cells on the approximate motor activity of rats after simulation of ischemic stroke. Eur. J. Clin. Investig. 2019, 49 (Suppl. S1), 161. [Google Scholar] [CrossRef]
- Shanko, Y.; Navitskaya, V.; Zamaro, A.; Krivenko, S.; Zafranskaya, M.; Pashkevich, S.; Koulchitsky, S.; Takalchik–Stukach, Y.; Denisov, A.; Kulchitsky, V. Prospects of perineural administration of autologous mesenchymal stem cells of adipose tissue in patients with cerebral infarction. Biomed. J. Sci. Tech. Res. 2018, 10, 3. [Google Scholar] [CrossRef]
- Vanin, A.; Poltorakov, A. NO spin trapping in biological systems. Front. Biosci. 2009, 14, 4427–4435. [Google Scholar] [CrossRef]
- Plonka, P.M.; Chlopicki, S.; Wisniewska, M.; Plonka, B.K. Kinetics of increased generation of NO in endotoxaemic rats as measured by EPR. Acta Biochim. Pol. 2003, 50, 807–813. [Google Scholar] [CrossRef]
- Kleschyov, A.L.; Wenzel, P.; Munzel, T. Electron paramagnetic resonance (EPR) spin trapping of biological nitric oxide. J. Chromatogr. B 2007, 851, 12–20. [Google Scholar] [CrossRef]
- Suzuki, Y.; Fujii, S.; Tominaga, T.; Yoshimoto, T.; Yoshimura, T.; Kamada, H. The origin of an EPR signal observed in dithiocarmate-loaded tissues Copper (II)-dithiocarmate complexes account for the narrow hyperfine lines. Biochim. Biophys. Acta 1997, 1335, 242–245. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrianov, V.V.; Kulchitsky, V.A.; Yafarova, G.G.; Bazan, L.V.; Bogodvid, T.K.; Deryabina, I.B.; Muranova, L.N.; Silantyeva, D.I.; Arslanov, A.I.; Paveliev, M.N.; et al. Investigation of NO Role in Neural Tissue in Brain and Spinal Cord Injury. Molecules 2023, 28, 7359. https://doi.org/10.3390/molecules28217359
Andrianov VV, Kulchitsky VA, Yafarova GG, Bazan LV, Bogodvid TK, Deryabina IB, Muranova LN, Silantyeva DI, Arslanov AI, Paveliev MN, et al. Investigation of NO Role in Neural Tissue in Brain and Spinal Cord Injury. Molecules. 2023; 28(21):7359. https://doi.org/10.3390/molecules28217359
Chicago/Turabian StyleAndrianov, Viacheslav V., Vladimir A. Kulchitsky, Guzel G. Yafarova, Leah V. Bazan, Tatiana K. Bogodvid, Irina B. Deryabina, Lyudmila N. Muranova, Dinara I. Silantyeva, Almaz I. Arslanov, Mikhail N. Paveliev, and et al. 2023. "Investigation of NO Role in Neural Tissue in Brain and Spinal Cord Injury" Molecules 28, no. 21: 7359. https://doi.org/10.3390/molecules28217359
APA StyleAndrianov, V. V., Kulchitsky, V. A., Yafarova, G. G., Bazan, L. V., Bogodvid, T. K., Deryabina, I. B., Muranova, L. N., Silantyeva, D. I., Arslanov, A. I., Paveliev, M. N., Fedorova, E. V., Filipovich, T. A., Nagibov, A. V., & Gainutdinov, K. L. (2023). Investigation of NO Role in Neural Tissue in Brain and Spinal Cord Injury. Molecules, 28(21), 7359. https://doi.org/10.3390/molecules28217359