

Structural Characterization and Properties of Modified Soybean Meal Protein via Solid-State Fermentation by Bacillus subtilis

Abstract
:
1. Introduction
2. Results
2.1. Optimization of the Fermentation Conditions
2.2. Protein Structure Analysis
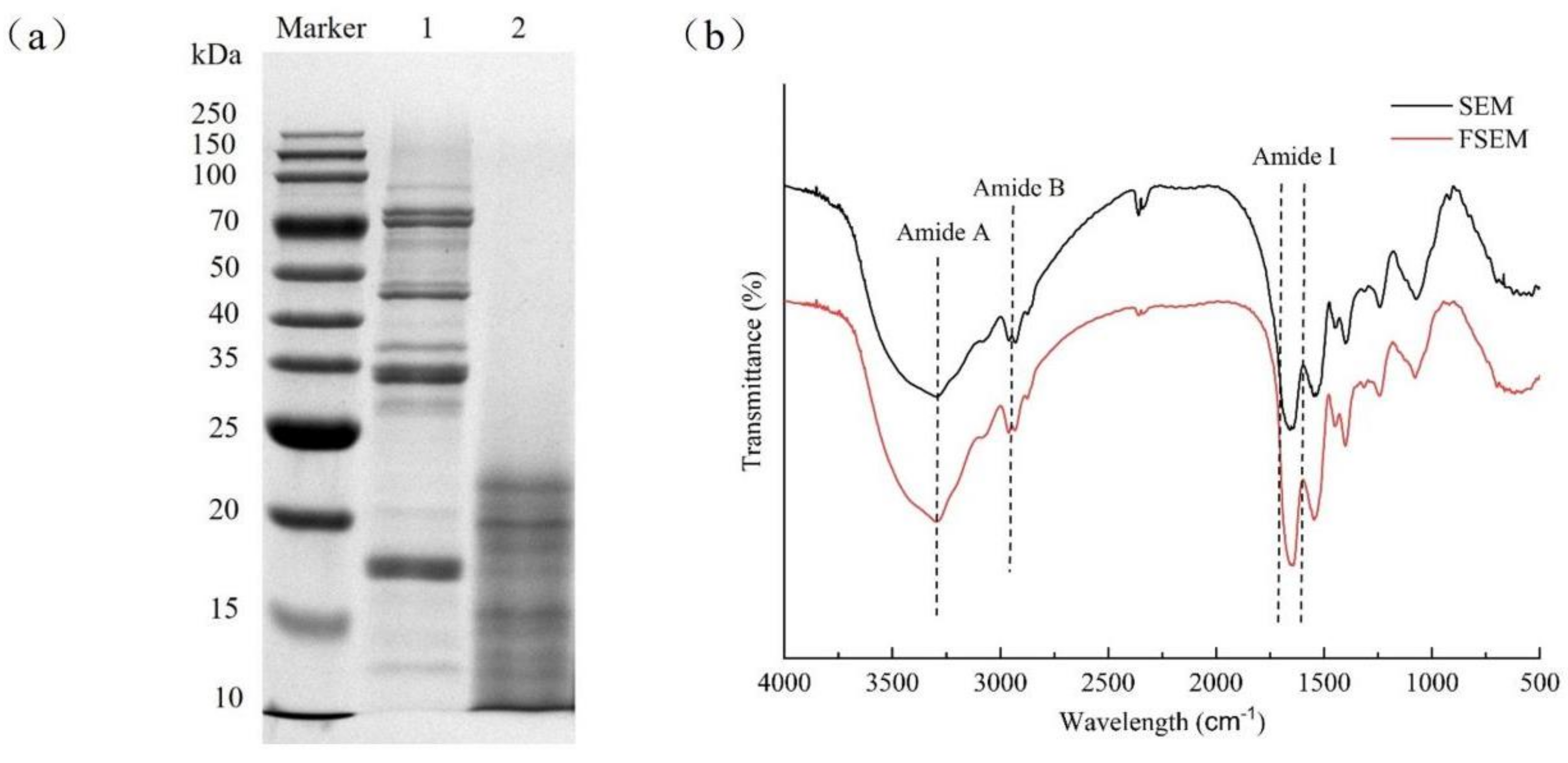
2.2.1. SDS-PAGE Protein Gel Electrophoresis
2.2.2. Fourier Transform Infrared Spectroscopy Assay
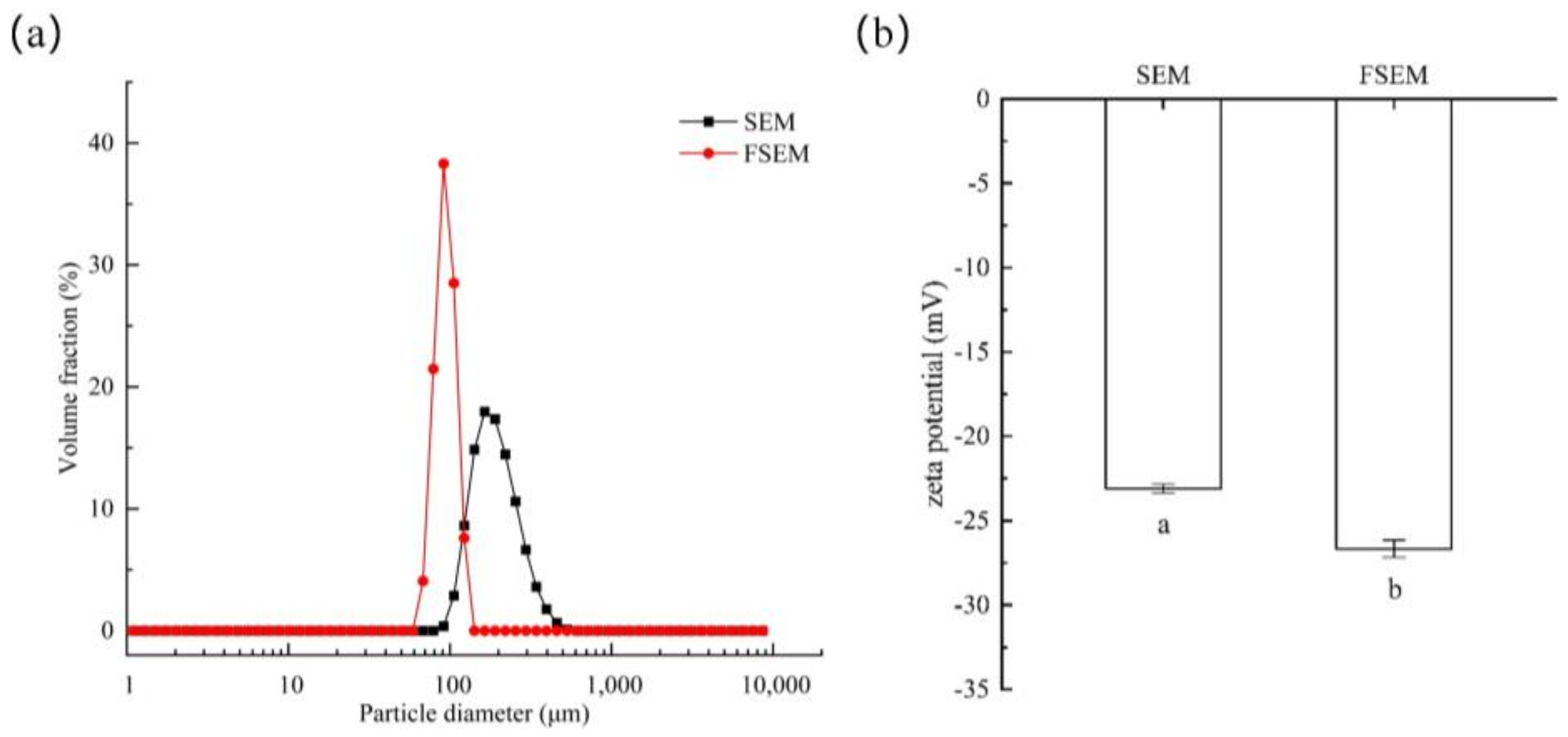
2.2.3. Particle Size Distribution
2.2.4. Zeta Potential
2.2.5. Scanning Electron Microscopy (SEM)
2.2.6. Confocal Laser Scanning Microscopy (CLSM)
2.3. Protein Properties Analysis
2.3.1. Solubility of Protein and Surface Hydrophobicity
2.3.2. Free Sulfhydryl Content
2.3.3. Antigenicity of Proteins
2.3.4. Binding Ability to Human Serum-Specific Antibodies (SIgE)
2.3.5. In Vitro Digestibility
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of the Culture Medium
4.3. Preparation of Fermented Soybean Meal
4.3.1. Strain Culture Method
4.3.2. RSM
4.4. Structural Analysis
4.4.1. Protein Preparation
4.4.2. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.4.3. FT-IR
4.4.4. Determination of Particle Size and Zeta Potential
4.4.5. SEM Analysis
4.4.6. CLSM Analysis
4.5. Protein Properties Analysis
4.5.1. Solubility of Protein
4.5.2. Surface Hydrophobicity
4.5.3. Free Sulfhydryl Group Content
4.5.4. Protein Antigenicity
4.5.5. Determination of Binding Ability to Human Serum-Specific Antibodies (SIgE)
4.5.6. In Vitro Digestibility
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, H.; Bian, Y.; Huang, L.; Lan, Q.; Ma, L.; Li, X.; Leng, X. Effects of replacing fish meal with fermented soybean meal on the growth performance, intestinal microbiota, morphology and disease resistance of largemouth bass (Micropterus salmoides). Aquac. Rep. 2022, 22, 100954. [Google Scholar] [CrossRef]
- Wu, P.; Guo, Y.; Golly, M.K.; Ma, H.; He, R.; Luo, S.; Zhang, C.; Zhang, L.; Zhu, J. Feasibility study on direct fermentation of soybean meal by Bacillus stearothermophilus under non-sterile conditions. J. Sci. Food Agric. 2019, 99, 3291–3298. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of Fermentation in Improving Nutritional Quality of Soybean Meal—A Review. Asian-Australas. J. Anim. Sci. 2015, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Raja, P.; Parthiban, M.; Ghadevaru, S. Isolation, molecular characterization, purification and production optimization of alpha galactosidase enzyme from Bacillus subtilis. Vet. Res. Int. 2020, 8, 88–93. [Google Scholar]
- Zheng, L.; Li, D.; Li, Z.L.; Kang, L.N.; Jiang, Y.Y.; Liu, X.Y.; Chi, Y.P.; Li, Y.Q.; Wang, J.H. Effects of Bacillus fermentation on the protein microstructure and anti-nutritional factors of soybean meal. Lett. Appl. Microbiol. 2017, 65, 520–526. [Google Scholar] [CrossRef]
- Yang, H.; Qu, J.; Zou, W.; Shen, W.; Chen, X. An overview and future prospects of recombinant protein production in Bacillus subtilis. Appl. Microbiol. Biotechnol. 2021, 105, 6607–6626. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, Z.; Yu, W.; Zheng, L.; Li, L.J.; Gu, W.; Xu, H.Y.; Wei, B.D.; Yan, X.G. Nutritional quality improvement of soybean meal by Bacillus velezensis and Lactobacillus plantarum during two-stage solid-state fermentation. AMB Express 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Ashaolu, T.J. Applications of soy protein hydrolysates in the emerging functional foods: A review. Int. J. Food Sci. Technol. 2020, 55, 421–428. [Google Scholar] [CrossRef]
- Hasan, M.N.; Sultan, M.Z.; Mar-E-Um, M. Significance of fermented food in nutrition and food science. J. Sci. Res. 2014, 6, 373–386. [Google Scholar] [CrossRef]
- Verni, M.; Rizzello, C.G.; Coda, R. Fermentation biotechnology applied to cereal industry by-products: Nutritional and functional insights. Front. Nutr. 2019, 6, 1–13. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 1–14. [Google Scholar] [CrossRef]
- Dai, C.; Hou, Y.; Xu, H.; Huang, L.; Dabbour, M.; Mintah, B.K.; He, R.H.; Ma, H. Effect of solid-state fermentation by three different Bacillus species on composition and protein structure of soybean meal. J. Sci. Food Agric. 2022, 102, 557–566. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Xu, Y.; Xiao, N.; Huang, J.Z.; Wu, S.S.; Zhou, Q.T.; Lai, Y.P.; Chen, J.P.; Li, P.; Du, B. Douchi ameliorates high-fat diet-induced hyperlipidaemia by regulation of intestinal microflora in rats. Int. J. Food Sci. Technol. 2022, 57, 2756–2769. [Google Scholar] [CrossRef]
- Czech, A.; Grela, E.R.; Kiesz, M. Dietary fermented rapeseed or/and soybean meal additives on performance and intestinal health of piglets. Sci. Rep. 2021, 11, 16952. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Silva, F.; Lemos, T.C.; Sandôra, D.; Monteiro, M.; Perrone, D. Fermentation of soybean meal improves isoflavone metabolism after soy biscuit consumption by adults. J. Sci. Food Agric. 2020, 100, 2991–2998. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, X.; Huang, J. Study on enzymatic hydrolysis of soybean β-conglycinin using alkaline protease from Bacillus subtilis ACCC 01746 and antigenicity of its hydrolysates. Grain Oil Sci. Technol. 2021, 4, 18–25. [Google Scholar] [CrossRef]
- Lu, F.; Alenyorege, E.A.; Ouyang, N.; Zhou, A.; Ma, H. Simulated natural and high temperature solid-state fermentation of soybean meal: A comparative study regarding microorganisms, functional properties and structural characteristics. LWT 2022, 159, 113125. [Google Scholar] [CrossRef]
- Shon, M.Y.; Kim, T.H.; Sung, N.J. Antioxidants and free radical scavenging activity of Phellinus baumii (Phellinus of Hymenochaetaceae) extracts. Food Chem. 2003, 82, 593–597. [Google Scholar] [CrossRef]
- Zhang, Y.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Dossou, S.; Wang, W.; Zhang, X.; Shadrack, R.S.; Mzengereza, K.; Zhu, K.H.; et al. Optimization of soybean meal fermentation for aqua-feed with Bacillus subtilis natto using the response surface methodology. Fermentation 2021, 7, 306. [Google Scholar] [CrossRef]
- Bailey, M.; Miller, B.G.; Telemo, E.; Stokes, C.R.; Bourne, F.J. Specific immunological unresponsiveness following active primary responses to proteins in the weaning diet of piglets. Int. Arch. Allergy Immunol. 1993, 101, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Yasar, S.; Tosun, R.; Sonmez, Z. Fungal fermentation inducing improved nutritional qualities associated with altered secondary protein structure of soybean meal determined by FTIR spectroscopy. Measurement 2020, 161, 107895. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Jiang, L.; Qi, B.; Zhou, L. Relationship between secondary structure and surface hydrophobicity of soybean protein isolate subjected to heat treatmen. J. Chem. 2014, 5, 475389. [Google Scholar] [CrossRef]
- Alrosan, M.; Tan, T.C.; Mat Easa, A.; Gammoh, S.; Alu’datt, M.H. Effects of fermentation on the quality, structure, and nonnutritive contents of lentil (Lens culinaris) proteins. J. Food Qual. 2021, 2021, 1–7. [Google Scholar] [CrossRef]
- Cheng, P.N.; Pham, J.D.; Nowick, J.S. The supramolecular chemistry of β-sheets. J. Am. Chem. Soc. 2013, 135, 5477–5492. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.C. Low-frequency motions in protein molecules. Beta-sheet and beta-barr. Biophys. J. 1985, 48, 289–297. [Google Scholar] [CrossRef]
- Salazar-Villanea, S.; Hendriks, W.H.; Bruininx, E.M.A.M.; Gruppen, H.; Van Der Poel, A.F.B. Protein structural changes during processing of vegetable feed ingredients used in swine diets: Implications for nutritional value. Nutr. Res. Rev. 2016, 29, 126–141. [Google Scholar] [CrossRef]
- Li, M.; Yang, R.; Feng, X.; Fan, X.; Liu, Y.; Xu, X.; Zhou, G.; Zhu, B.; Ullah, N.; Chen, L. Effects of low-frequency and high-intensity ultrasonic treatment combined with curdlan gels on the thermal gelling properties and structural properties of soy protein isolate. Food Hydrocoll. 2022, 127, 107506. [Google Scholar] [CrossRef]
- Tang, C.H.; Wang, X.Y.; Yang, X.Q.; Li, L. Formation of soluble aggregates from insoluble commercial soy protein isolate by means of ultrasonic treatment and their gelling properties. J. Food Eng. 2009, 92, 432–437. [Google Scholar] [CrossRef]
- Zhu, P.; Huang, W.; Chen, L. Develop and characterize thermally reversible transparent gels from pea protein isolate and study the gel formation mechanisms. Food Hydrocoll. 2022, 125, 107373. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, W.; Ding, X.; Wu, Z.; Li, Y. Effects of dual-frequency ultrasound with different energy irradiation modes on the structural and emulsifying properties of soy protein isolate. Food Bioprod. Process. 2020, 123, 419–426. [Google Scholar] [CrossRef]
- Song, Y.S.; Frías, J.; Martinez-Villaluenga, C.; Vidal-Valdeverde, C.; de Mejia, E.G. Immunoreactivity reduction of soybean meal by fermentation, effect on amino acid composition and antigenicity of commercial soy products. Food Chem. 2008, 108, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Zuo, L.L.; Cheng, Y.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Degradation of major allergens and allergenicity reduction of soybean meal through solid-state fermentation with microorganisms. Food Funct. 2018, 9, 1899–1909. [Google Scholar] [CrossRef]
- Ruan, S.; Li, Y.; Wang, Y.; Huang, S.; Luo, J.; Ma, H. Analysis in protein profile, antioxidant activity and structure-activity relationship based on ultrasound-assisted liquid-state fermentation of soybean meal with Bacillus subtilis. Ultrason. Sonochemistry 2020, 64, 104846. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Huang, Y.; Zhang, X.; Zeng, J.; Zou, J.; Zhang, L.; Gong, P. Texture analysis and physicochemical characteristics of fermented soymilk gel by different lactic acid bacteria. Food Hydrocoll. 2023, 136, 108252. [Google Scholar] [CrossRef]
- Jääskeläinen, A.S.; Holopainen-Mantila, U.; Tamminen, T.; Vuorinen, T. Endosperm and aleurone cell structure in barley and wheat as studied by optical and Raman microscopy. J. Cereal Sci. 2013, 57, 543–550. [Google Scholar] [CrossRef]
- Ma, S.; Wang, Z.; Tian, X.L.; Sun, B.H.; Huang, J.H.; Yan, J.Y.; Bao, Q.D.; Wang, X.X. Effect of synergistic fermentation of Lactobacillus plantarum and Saccharomyces cerevisiae on thermal properties of wheat bran dietary fiber-wheat starch system. Food Chem. 2022, 373, 131417. [Google Scholar] [CrossRef]
- Samoto, M.; Maebuchi, M.; Miyazaki, C.; Kugitani, H.; Kohno, M.; Hirotsuka, M.; Kito, M. Abundant proteins associated with lecithin in soy protein isolate. Food Chem. 2007, 102, 317–322. [Google Scholar] [CrossRef]
- Beveridge, T.T.S.J.; Toma, S.J.; Nakai, S. Determination of SH-and SS-groups in some food proteins using Ellman’s reagent. J. Food Sci. 1974, 39, 49–51. [Google Scholar] [CrossRef]
- Peñas, E.; Gomez, R.; Frias, J.; Baeza, M.L.; Vidal-Valverde, C. High hydrostatic pressure effects on immunoreactivity and nutritional quality of soybean products. Food Chem. 2011, 125, 423–429. [Google Scholar] [CrossRef]
- Tang, C.H. Functional properties and in vitro digestibility of buckwheat protein products: Influence of processing. J. Food Eng. 2007, 82, 568–576. [Google Scholar] [CrossRef]
- Teng, C.; Ma, K.D.; Ma, K.Y.; Feng, J.; Xu, L.J.; Li, Y. Effect of burdock on the regulation of gut microbiota in hyperglycemic rats and its in vitro digestion and fermentation characteristics. Food Biosci. 2023, 56, 103191. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Runs | Inoculation Amount (%) | Time (h) | Material-Liquid Ratio (g/mL) | Acid-Soluble Protein Content (g/100 g) |
|---|---|---|---|---|
| 1 | −1.00 | 0.00 | 1.00 | 7.0518 |
| 2 | 0.00 | 1.00 | −1.00 | 6.7463 |
| 3 | −1.00 | −1.00 | 0.00 | 6.6209 |
| 4 | 0.00 | 1.00 | 1.00 | 6.0381 |
| 5 | 0.00 | 0.00 | 0.00 | 12.7656 |
| 6 | −1.00 | 1.00 | 0.00 | 6.1621 |
| 7 | 0.00 | 0.00 | 0.00 | 12.7742 |
| 8 | 1.00 | −1.00 | 0.00 | 8.4825 |
| 9 | 0.00 | −1.00 | 1.00 | 6.4956 |
| 10 | 0.00 | −1.00 | −1.00 | 7.2193 |
| 11 | 0.00 | 0.00 | 0.00 | 12.9152 |
| 12 | 1.00 | 1.00 | 0.00 | 7.7903 |
| 13 | −1.00 | 0.00 | −1.00 | 6.8049 |
| 14 | 1.00 | 0.00 | 1.00 | 7.5611 |
| 15 | 1.00 | 0.00 | −1.00 | 8.8643 |
| 16 | 0.00 | 0.00 | 0.00 | 12.6207 |
| 17 | 0.00 | 0.00 | 0.00 | 12.8134 |
| Source | Sum of Squares | Df | Mean Square | F Value | p Value | Significance 1 |
|---|---|---|---|---|---|---|
| Model | 120.04 | 9 | 13.34 | 534.27 | <0.0001 | *** |
| A-Inoculation amount | 4.59 | 1 | 4.59 | 183.79 | <0.0001 | *** |
| B-Time | 0.54 | 1 | 0.54 | 21.69 | 0.0023 | ** |
| C-Material-liquid Ratio | 0.77 | 1 | 0.77 | 31.00 | 0.0008 | ** |
| AB | 0.014 | 1 | 0.014 | 0.55 | 0.4842 | |
| AC | 0.60 | 1 | 0.60 | 24.06 | 0.0017 | * |
| BC | 6.006 × 10−5 | 1 | 6.006 × 10−5 | 2.406 × 10−3 | 0.9622 | |
| A2 | 21.97 | 1 | 21.97 | 879.90 | <0.0001 | *** |
| B2 | 43.92 | 1 | 43.92 | 1759.36 | <0.0001 | *** |
| C2 | 35.98 | 1 | 35.98 | 1441.21 | <0.0001 | *** |
| Residual | 0.17 | 7 | 0.025 | |||
| Lack of Fit | 0.13 | 3 | 0.043 | 3.85 | 0.1130 | |
| Pure Error | 0.045 | 4 | 0.011 | |||
| Cor total | 120.22 | 16 | R2 = 0.9985 |
| Sample | α-Helix (%) | β-Sheet (g/100 g) | β-Turn (%) | Random Coil (%) | Ratio 1 (%) |
|---|---|---|---|---|---|
| SBM | 24.45 ± 0.18 a | 29.51 ± 0.67 a | 33.16 ± 0.10 a | 12.88 ± 0.79 a | 82.85 |
| FSBM | 16.15 ± 0.07 b | 32.59 ± 0.76 b | 35.87 ± 0.93 b | 15.39 ± 0.61 b | 48.55 |
| Levels | Inoculation Amount (%) | Time (h) | Material-Liquid Ratio (g/mL) |
|---|---|---|---|
| −1 | 9 | 24 | 1:0.4 |
| 0 | 12 | 48 | 1:0.6 |
| 1 | 15 | 72 | 1:0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, X.; Niu, H.; Sun, M.; Li, D.; Hua, M.; Wang, J.; Su, Y. Structural Characterization and Properties of Modified Soybean Meal Protein via Solid-State Fermentation by Bacillus subtilis. Molecules 2023, 28, 8015. https://doi.org/10.3390/molecules28248015
Miao X, Niu H, Sun M, Li D, Hua M, Wang J, Su Y. Structural Characterization and Properties of Modified Soybean Meal Protein via Solid-State Fermentation by Bacillus subtilis. Molecules. 2023; 28(24):8015. https://doi.org/10.3390/molecules28248015
Chicago/Turabian StyleMiao, Xinyu, Honghong Niu, Mubai Sun, Da Li, Mei Hua, Jinghui Wang, and Ying Su. 2023. "Structural Characterization and Properties of Modified Soybean Meal Protein via Solid-State Fermentation by Bacillus subtilis" Molecules 28, no. 24: 8015. https://doi.org/10.3390/molecules28248015
APA StyleMiao, X., Niu, H., Sun, M., Li, D., Hua, M., Wang, J., & Su, Y. (2023). Structural Characterization and Properties of Modified Soybean Meal Protein via Solid-State Fermentation by Bacillus subtilis. Molecules, 28(24), 8015. https://doi.org/10.3390/molecules28248015