


Strictinin, a Major Ingredient in Yunnan Kucha Tea Possessing Inhibitory Activity on the Infection of Mouse Hepatitis Virus to Mouse L Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
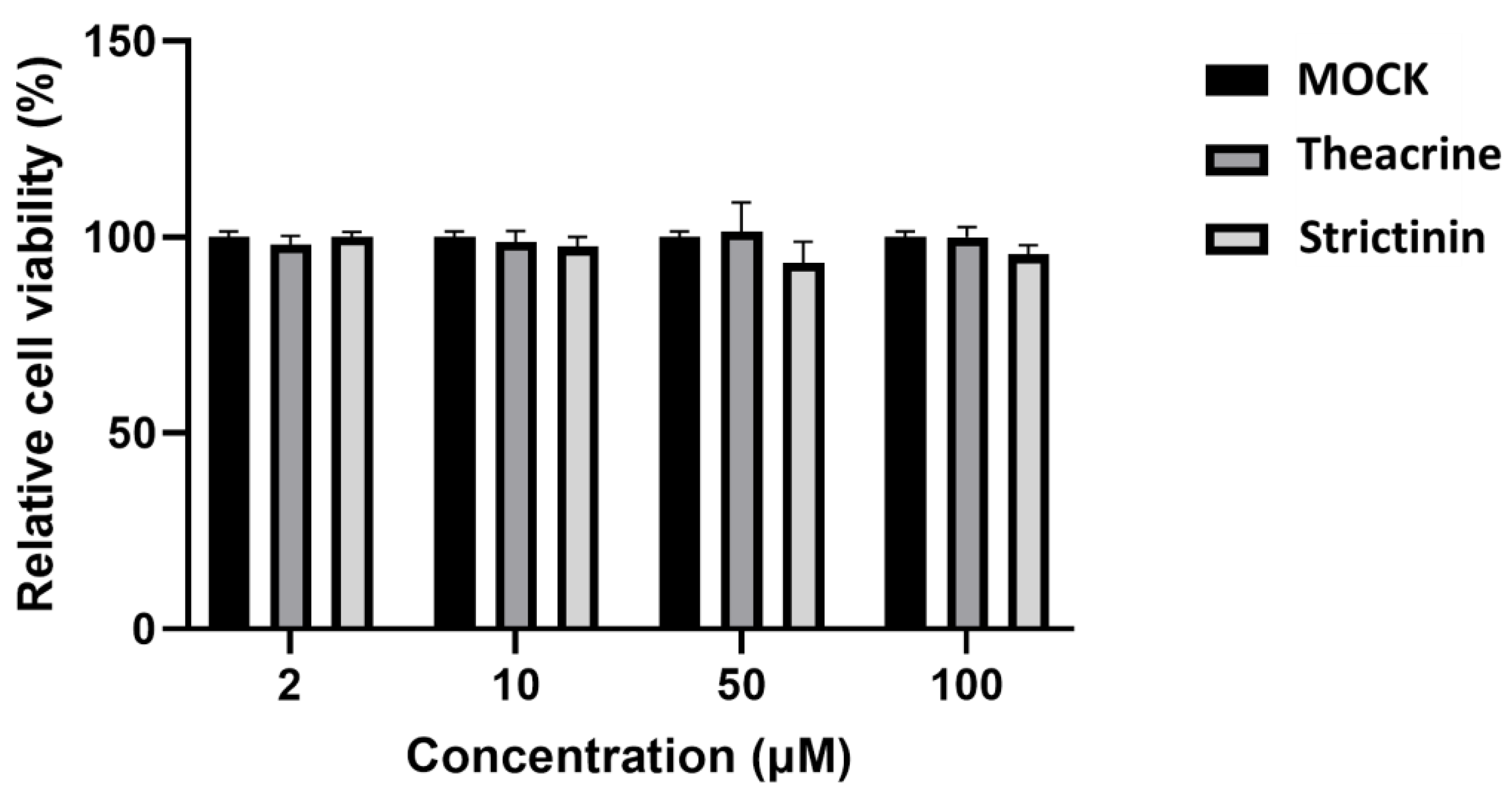
2.1. Cytotoxicity of Theacrine and Strictinin to Mouse L Cells
2.2. Effects of Theacrine and Strictinin on MHV Infection of Mouse L Cells
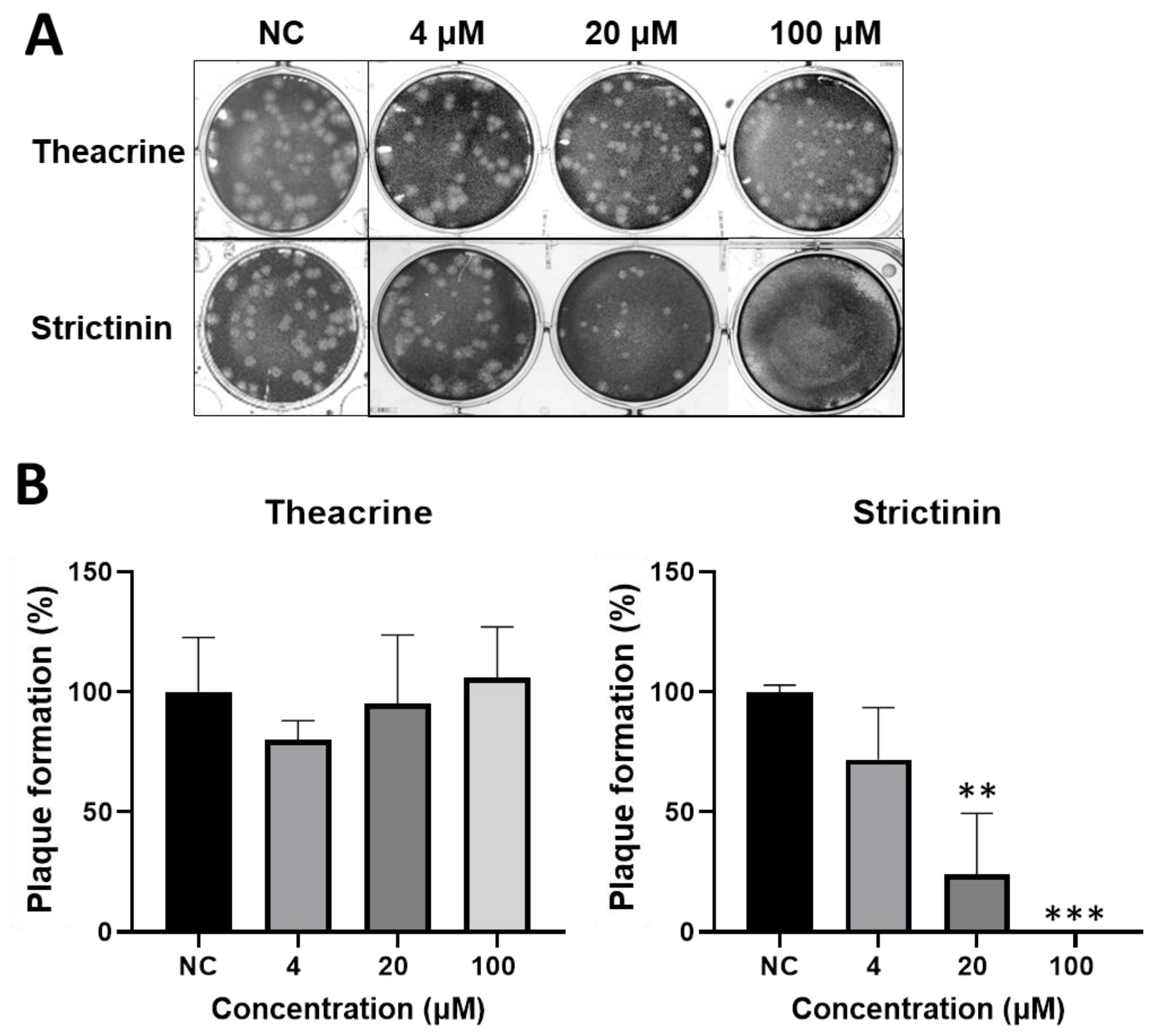
2.2.1. Inhibitory Activity of Theacrine and Strictinin on Plaque Formation
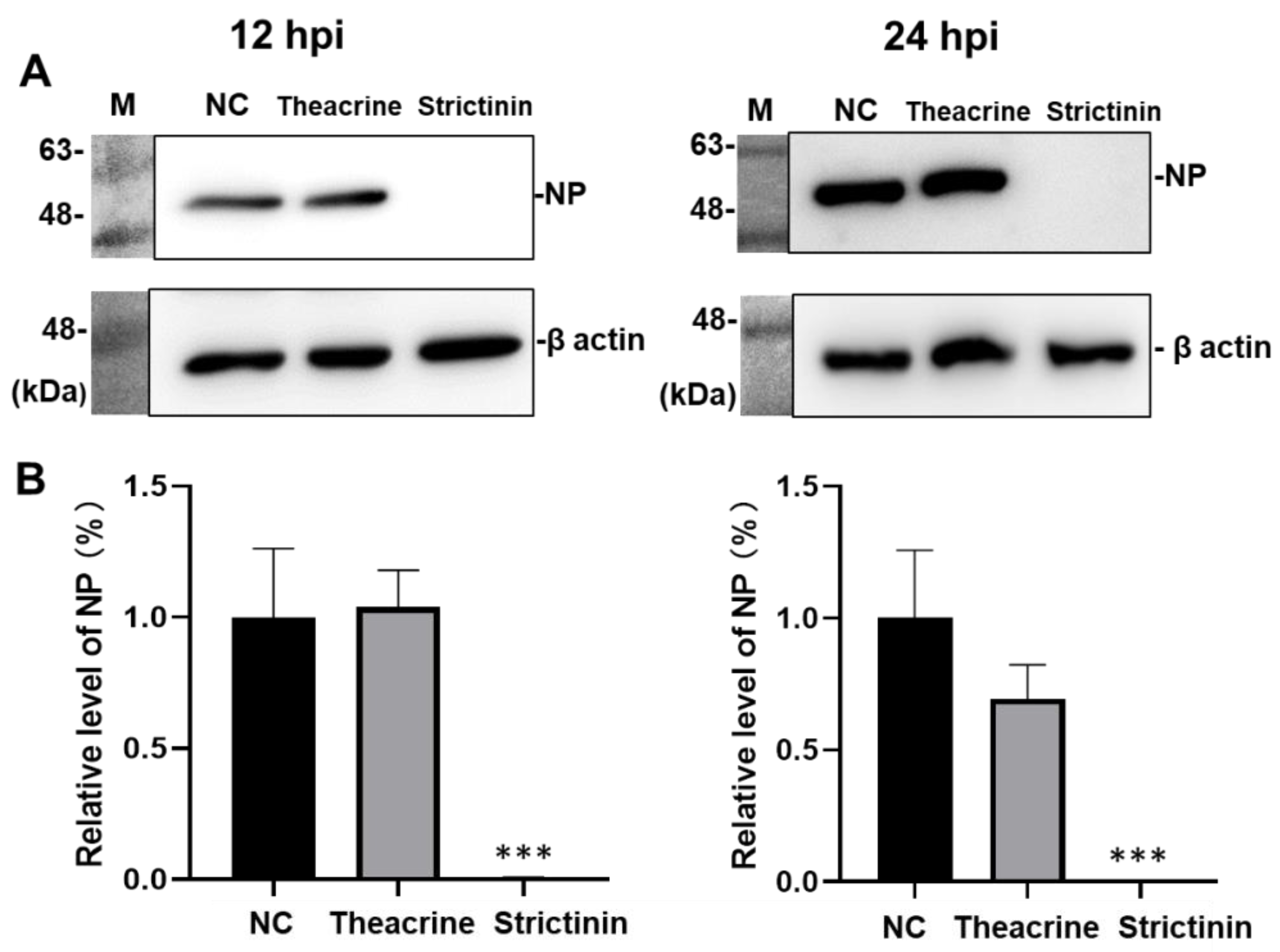
2.2.2. Effects of Theacrine and Strictinin on the Level of Nucleocapsid Protein
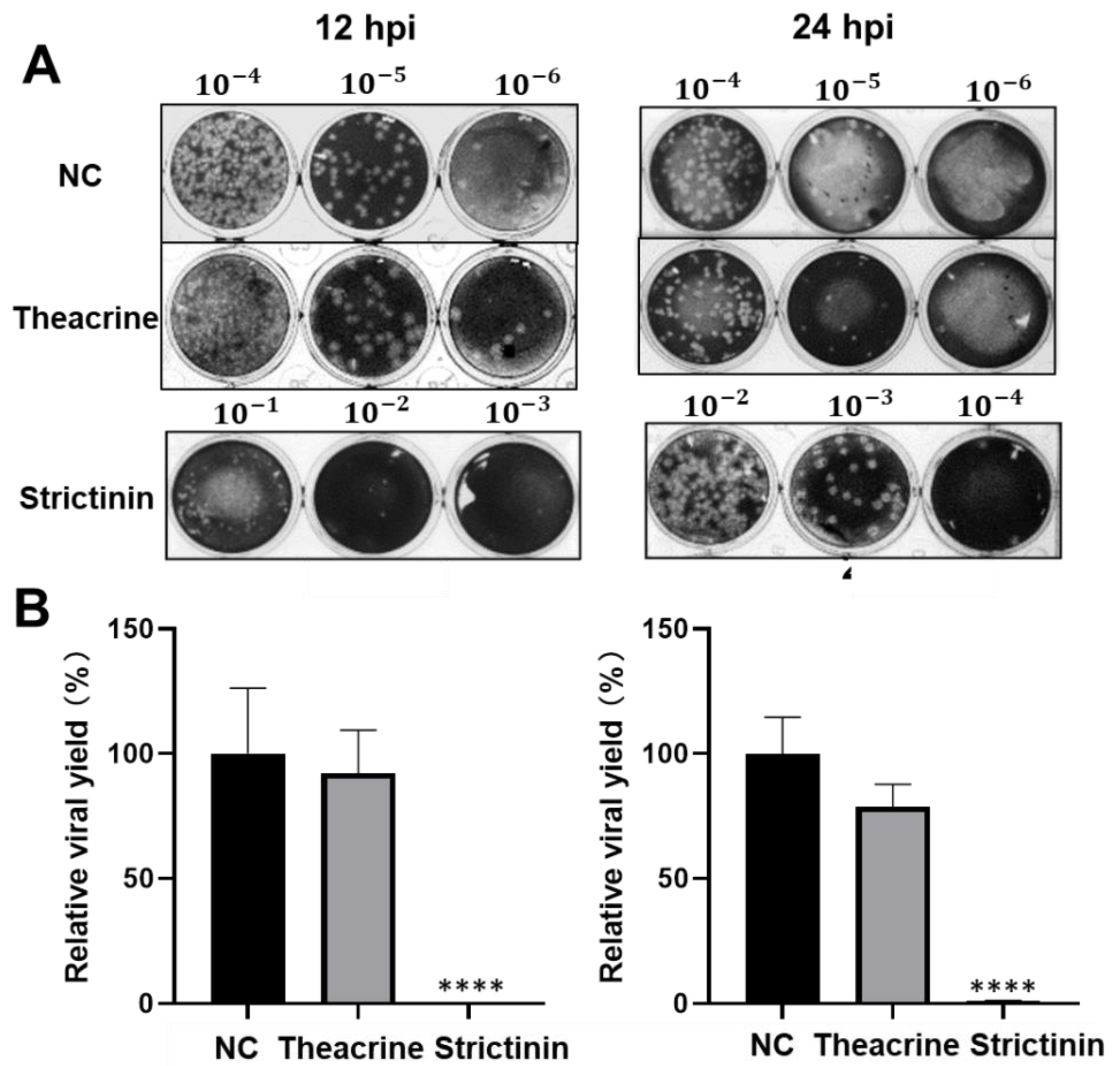
2.2.3. Effects of Theacrine and Strictinin on Viral Progeny Production
2.3. Comparison of Effects of Remdesivir and Strictinin on MHV Infection of Mouse L Cells
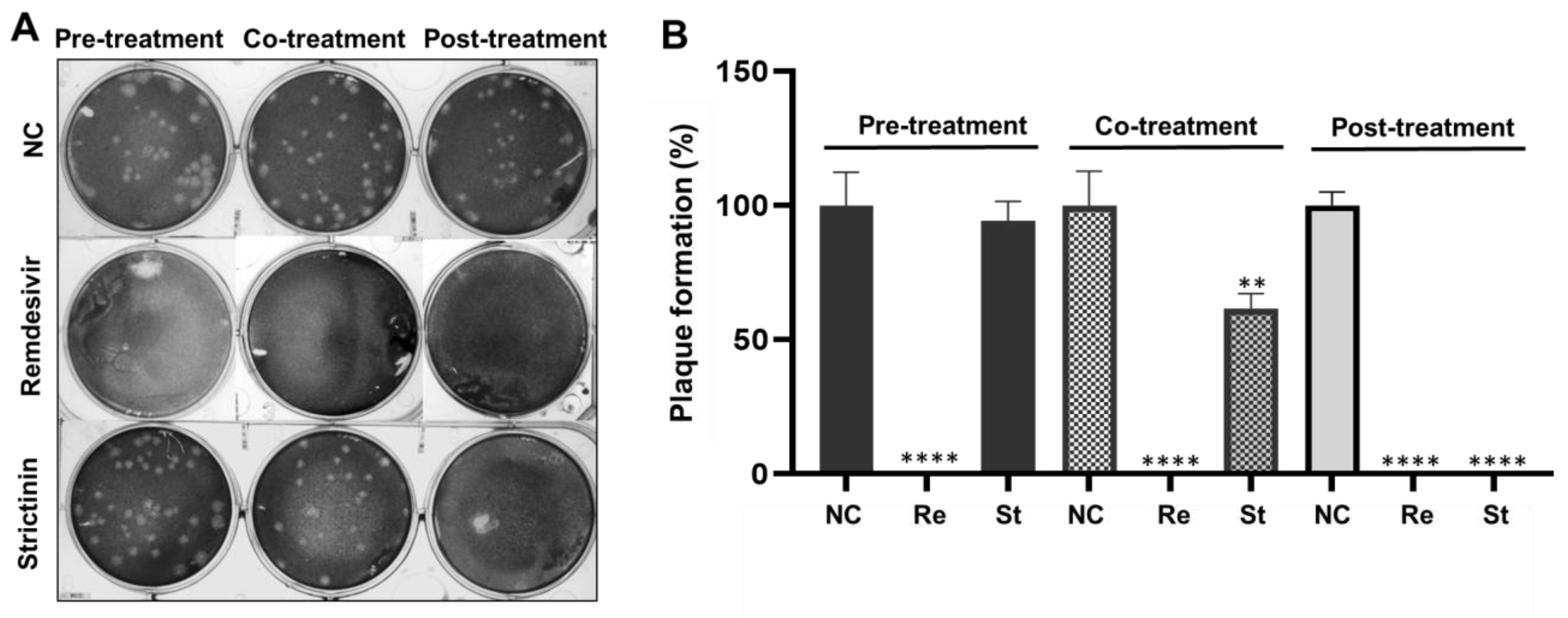
2.3.1. Inhibitory Activity of Remdesivir and Strictinin on Plaque Formation
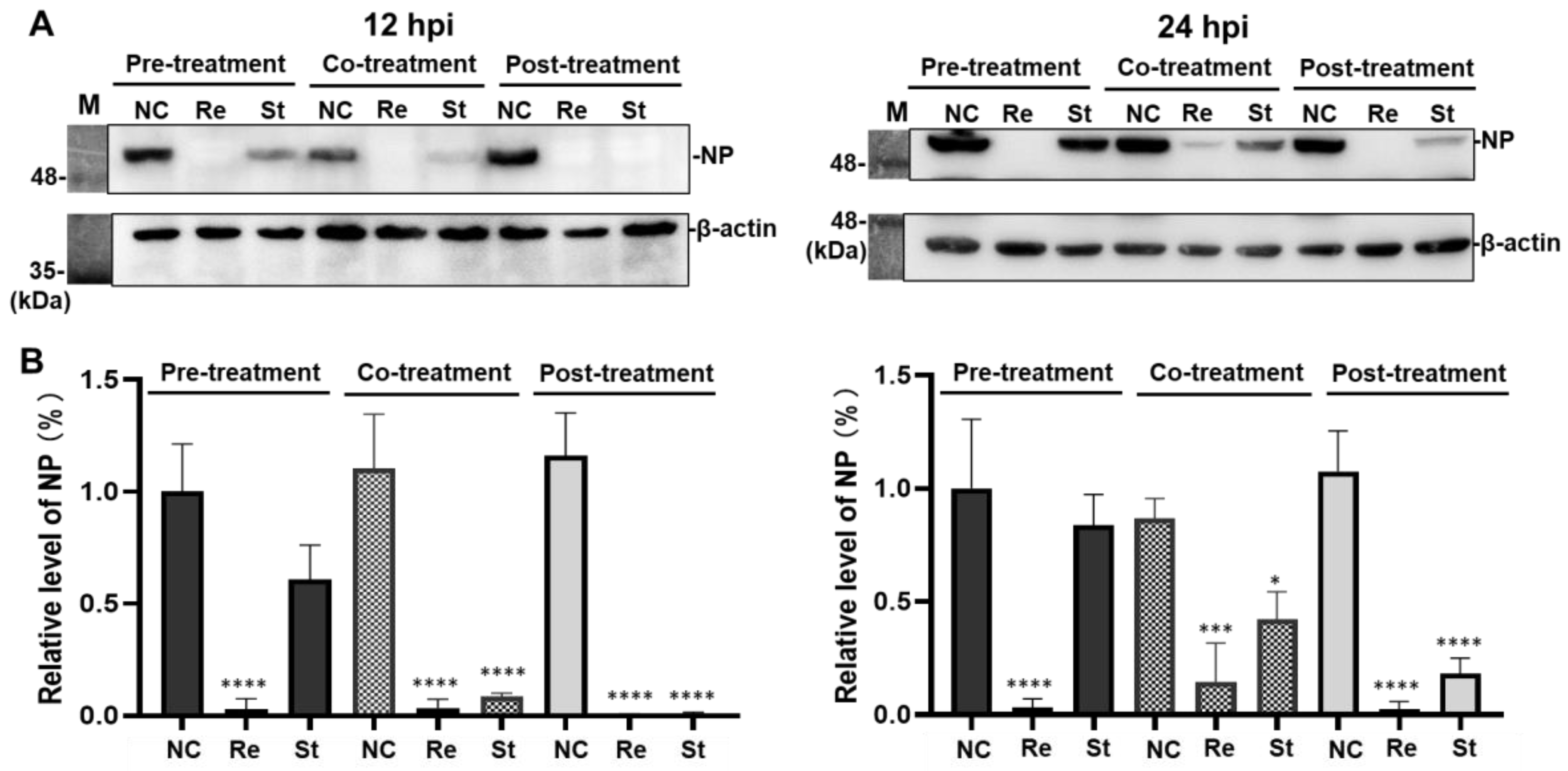
2.3.2. Effects of Remdesivir and Strictinin on the Level of Nucleocapsid Protein
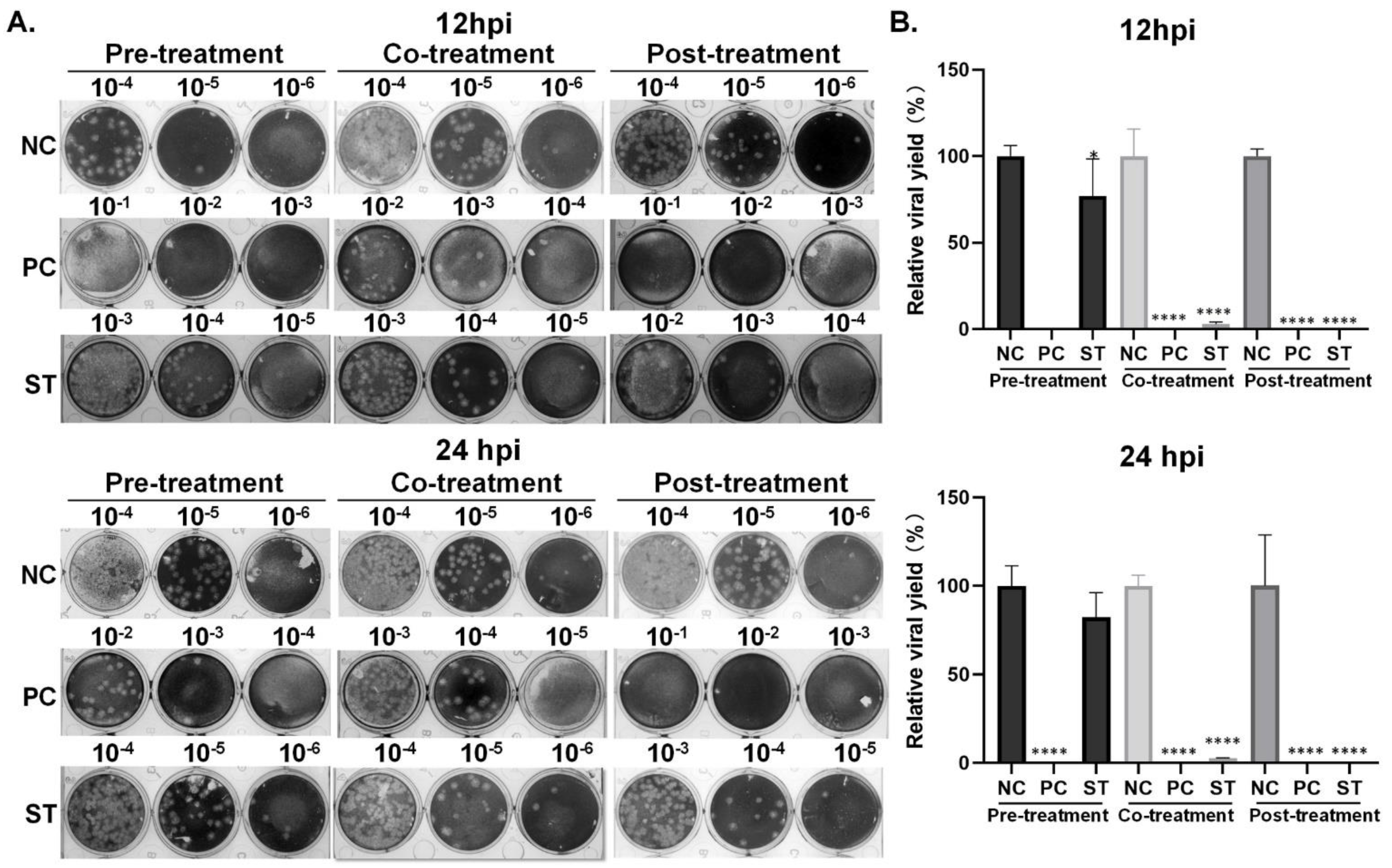
2.3.3. Effects of Remdesivir and Strictinin on Viral Progeny Production
3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Test Compounds
4.3. Cytotoxicity Test
4.4. Viral Infection of Mouse L Cells with Different Treatments of Test Compounds
4.5. Plaque Reduction Assay
4.6. Western Blot Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parashar, U.D.; Anderson, L.J. Severe acute respiratory syndrome: Review and lessons of the 2003 outbreak. Int. J. Epidemiol. 2004, 33, 628–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Wu, T.; Liu, Q.; Yang, Z. The SARS-CoV-2 outbreak: What we know. Int. J. Infect. Dis. 2020, 94, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Sakayori, M.; Hagiwara, E.; Baba, T.; Kitamura, H.; Sekine, A.; Ikeda, S.; Tabata, E.; Yamada, S.; Fujimoto, K.; Ogura, T. Incidence of acute exacerbation in patients with interstitial lung disease after COVID-19 vaccination. J. Infect. Chemother. 2023, 29, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, H.; Yang, L.; Wang, Q. Progress on COVID-19 chemotherapeutics discovery and novel technology. Molecules 2022, 27, 8257. [Google Scholar] [CrossRef]
- Sahin, G.; Akbal-Dagistan, O.; Culha, M.; Erturk, A.; Basarir, N.S.; Sancar, S.; Yildiz-Pekoz, A. Antivirals and the potential benefits of orally inhaled drug administration in COVID-19 treatment. J. Pharm. Sci. 2022, 111, 2652–2661. [Google Scholar] [CrossRef] [PubMed]
- Duarte, N.B.A.; Takahashi, J.A. Plant spices as a source of antimicrobial synergic molecules to treat bacterial and viral co-infections. Molecules 2022, 27, 8210. [Google Scholar] [CrossRef]
- Ren, Y.; Frank, T.; Meyer, G.; Lei, J.; Grebenc, J.R.; Slaughter, R.; Gao, Y.G.; Kinghorn, A.D. Potential benefits of black chokeberry (Aronia melanocarpa) fruits and their constituents in improving human health. Molecules 2022, 27, 7823. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, M.; Dinda, S.; Chakraborty, M. Some natural compounds and their analogues having potent anti- SARS-CoV-2 and anti-proteases activities as lead molecules in drug discovery for COVID-19. Eur. J. Med. Chem. Rep. 2022, 6, 100079. [Google Scholar] [CrossRef]
- Begum, R.; Mamun-Or-Rashid, A.N.M.; Lucy, T.T.; Pramanik, M.K.; Sil, B.K.; Mukerjee, N.; Tagde, P.; Yagi, M.; Yonei, Y. Potential therapeutic approach of melatonin against omicron and some other variants of SARS-CoV-2. Molecules 2022, 27, 6934. [Google Scholar] [CrossRef]
- Goc, A.; Rath, M.; Niedzwiecki, A. Composition of naturally occurring compounds decreases activity of Omicron and SARS-CoV-2 RdRp complex. Eur. J. Microbiol. Immunol. 2022, 12, 39–45. [Google Scholar] [CrossRef]
- Elsebai, M.F.; Albalawi, M.A. Essential Oils and COVID-19. Molecules 2022, 27, 7893. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.Y.; Jeon, S.; Ko, M.; Du, Y.E.; Son, S.R.; Jang, D.S.; Kim, S.; Lee, C.J. Lancemaside A from Codonopsis lanceolata: Studies on antiviral activity and mechanism of action against SARS-CoV-2 and its variants of concern. Antimicrob. Agents Chemother. 2022, 66, e0120122. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Zhang, M.; Xu, Q.; Song, F.; Wang, L.; Gai, S.; Tang, H.; Wang, S.; Zhou, L.; Li, H. Exploration of molecular targets and mechanisms of Chinese medicinal formula Acacia Catechu -Scutellariae Radix in the treatment of COVID-19 by a systems pharmacology strategy. Phytother. Res. 2022, 36, 4210–4229. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Zhao, M.; Zhang, H.; Liu, C. The chemical and pharmacological research progress on a kind of Chinese herbal medicine, Fructus Malvae. Molecules 2022, 27, 5678. [Google Scholar] [CrossRef]
- Petitjean, S.J.L.; Lecocq, M.; Lelong, C.; Denis, R.; Defrère, S.; Mariage, P.A.; Alsteens, D.; Pilette, C. Salvia miltiorrhiza Bunge as a potential natural compound against COVID-19. Cells 2022, 11, 1311. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Yen, G.C.; Wang, B.S.; Chiu, C.K.; Yen, W.J.; Chang, L.W.; Duh, P.D. Antimutagenic and antimicrobial activities of pu-erh tea. LWT Food Sci. Technol. 2007, 40, 506–512. [Google Scholar] [CrossRef]
- Hou, Y.; Shao, W.; Xiao, R.; Xu, K.; Ma, Z.; Johnstone, B.H.; Du, Y. Pu-erh tea aqueous extracts lower atherosclerotic risk factors in a rat hyperlipidemia model. Exp. Gerontol. 2009, 44, 434–439. [Google Scholar] [CrossRef]
- Lin, P.R.; Kuo, P.C.; Li, Y.C.; Jhuo, C.F.; Hsu, W.L.; Tzen, J.T.C. Theacrine and strictinin, two major ingredients for the anti-influenza activity of Yunnan Kucha tea. J. Ethnopharmacol. 2020, 262, 113190. [Google Scholar] [CrossRef]
- Feduccia, A.A.; Wang, Y.; Simms, J.A.; Yi, H.Y.; Li, R.; Bjeldanes, L.; Ye, C.; Bartlett, S.E. Locomotor activation by theacrine, a purine alkaloid structurally similar to caffeine: Involvement of adenosine and dopamine receptors. Pharmacol. Biochem. Behav. 2012, 102, 241–248. [Google Scholar] [CrossRef]
- Qiao, H.; Ye, X.; Bai, X.; He, J.; Li, T.; Zhang, J.; Zhang, W.; Xu, J. Theacrine: A purine alkaloid from Camellia assamica var. kucha with a hypnotic property via the adenosine system. Neurosci. Lett. 2017, 659, 48–53. [Google Scholar] [CrossRef]
- Xu, J.K.; Kurihara, H.; Zhao, L.; Yao, X.S. Theacrine, a special purine alkaloid with sedative and hypnotic properties from Cammelia assamica var. kucha in mice. J. Asian Nat. Prod. Res. 2007, 9, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.Y.; Xiang, J.; Wang, Z.S.; Jin, J.; Wang, Y.Q.; Li, Q.S.; Li, D.; Fang, Z.T.; Lu, J.L.; Ye, J.H.; et al. Theacrine from Camellia kucha and Its Health Beneficial Effects. Front. Nutr. 2020, 7, 596823. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.H.; Zhai, Y.J.; Wu, Y.P.; Xie, G.; Wang, G.E.; Mao, Z.F.; Hu, H.H.; Luo, X.H.; Sun, W.Y.; Liang, L.; et al. Theacrine, a potent antidepressant purine alkaloid from a special Chinese tea, promotes adult hippocampal neurogenesis in stressed mice. J. Agric. Food Chem. 2021, 69, 7016–7027. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Yang, M.H.; Baek, S.H.; Nam, D.; Jung, S.H.; Ahn, K.S. Theacrine attenuates epithelial mesenchymal transition in human breast cancer MDA-MB-231 cells. Phytother. Res. 2019, 33, 1934–1942. [Google Scholar] [CrossRef]
- Jhuo, C.F.; Hsu, Y.Y.; Chen, W.Y.; Tzen, J.T.C. Attenuation of tumor development in mammary carcinoma rats by theacrine, an antagonist of adenosine 2A receptor. Molecules 2021, 26, 7455. [Google Scholar] [CrossRef]
- Saha, R.K.; Takahashi, T.; Kurebayashi, Y.; Fukushima, K.; Minami, A.; Kinbara, N.; Ichitani, M.; Sagesaka, Y.M.; Suzuki, T. Antiviral effect of strictinin on influenza virus replication. Antivir. Res. 2010, 88, 10–18. [Google Scholar] [CrossRef]
- Chen, G.H.; Lin, Y.L.; Hsu, W.L.; Hsieh, S.K.; Tzen, J.T.C. Significant elevation of antiviral activity of strictinin from Pu’er tea after thermal degradation to ellagic acid and gallic acid. J. Food Drug Anal. 2015, 23, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, S.K.; Xu, J.R.; Lin, N.H.; Li, Y.C.; Chen, G.H.; Kuo, P.C.; Chen, W.Y.; Tzen, J.T.C. Antibacterial and laxative activities of strictinin isolated from Pu’er tea (Camellia sinensis). J. Food Drug Anal. 2016, 24, 722–729. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.Y.; Wang, M.M.C.; Hsieh, S.K.; Hsieh, M.H.; Chen, W.Y.; Tzen, J.T.C. Pancreatic lipase inhibition of strictinin isolated from Pu’er tea (Cammelia sinensis) and its anti-obesity effects in C57BL6 mice. J. Func. Foods 2018, 48, 1–8. [Google Scholar] [CrossRef]
- Lin, P.Y.; Jhuo, C.F.; Lin, N.H.; Chen, W.Y.; Tzen, J.T.C. Assessing anti-psoriatic effects of bitter Pu’er tea and its three major compounds, strictinin, theacrine and epigallocatechin gallate in Iimiquimod-treated mice. Compounds 2022, 2, 293–306. [Google Scholar] [CrossRef]
- Salazar-García, M.; Acosta-Contreras, S.; Rodríguez-Martínez, G.; Cruz-Rangel, A.; Flores-Alanis, A.; Patiño-López, G.; Luna-Pineda, V.M. Pseudotyped vesicular stomatitis virus-severe acute respiratory syndrome-coronavirus-2 spike for the study of variants, vaccines, and therapeutics against coronavirus disease 2019. Front. Microbiol. 2022, 12, 817200. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Bertsch, P.M.; Bibby, K.; Haramoto, E.; Hewitt, J.; Huygens, F.; Gyawali, P.; Korajkic, A.; Riddell, S.; Sherchan, S.P.; et al. Decay of SARS-CoV-2 and surrogate murine hepatitis virus RNA in untreated wastewater to inform application in wastewater-based epidemiology. Environ. Res. 2020, 191, 110092. [Google Scholar] [CrossRef]
- Featherstone, A.B.; Brown, A.C.; Chitlapilly Dass, S. Murine hepatitis virus, a biosafety level 2 model for SARS-CoV-2, can remain viable on meat and meat packaging materials for at least 48 hours. Microbiol. Spectr. 2022, 10, e0186222. [Google Scholar] [CrossRef] [PubMed]
- Körner, R.W.; Majjouti, M.; Alcazar, M.A.A.; Mahabir, E. Of mice and men: The coronavirus MHV and mouse models as a translational approach to understand SARS-CoV-2. Viruses 2020, 12, 880. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.L.; Tan, K.S.W.; Chu, J.J.H.; Chow, V.T. Combination treatment with remdesivir and ivermectin exerts highly synergistic and potent antiviral activity against murine coronavirus infection. Front. Cell Infect. Microbiol. 2021, 11, 700502. [Google Scholar] [CrossRef]
- Tian, L.; Pang, Z.; Li, M.; Lou, F.; An, X.; Zhu, S.; Song, L.; Tong, Y.; Fan, H.; Fan, J. Molnupiravir and its antiviral activity against COVID-19. Front. Immunol. 2022, 13, 855496. [Google Scholar] [CrossRef]
- Landete, J. Ellagitannins, ellagic acid and their derived metabolites: A review about source, metabolism, functions and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Falade, V.A.; Adelusi, T.I.; Adedotun, I.O.; Abdul-Hammed, M.; Lawal, T.A.; Agboluaje, S.A. In silico investigation of saponins and tannins as potential inhibitors of SARS-CoV-2 main protease (M pro). In Silico Pharmacol. 2021, 9, 9. [Google Scholar] [CrossRef]
- Khalifa, I.; Zhu, W.; Mohammed, H.H.H.; Dutta, K.; Li, C. Tannins inhibit SARS-CoV-2 through binding with catalytic dyad residues of 3CLpro: An in silico approach with 19 structural different hydrolysable tannins. J. Food Biochem. 2020, 44, e13432. [Google Scholar] [CrossRef]
- Chen, C.N.; Lin, C.P.; Huang, K.K.; Chen, W.C.; Hsieh, H.P.; Liang, P.H.; Hsu, J.T.A. Inhibition of SARS-CoV 3C-like protease activity by theaflavin-3, 3’-digallate (TF3). J. Evid. Based Complement. Altern. Med. 2005, 2, 209–215. [Google Scholar] [CrossRef]
- Deaville, E.R.; Green, R.J.; Mueller-Harvey, I.; Willoughby, I.; Frazier, R.A.J. Hydrolyzable tannin structures influence relative globular and random coil protein binding strengths. J. Agric. Food Chem. 2007, 55, 4554–4561. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Chen, T.Y.; Lin, S.C.; Chung, C.Y.; Lin, T.C.; Wang, G.H.; Anderson, R.; Lin, C.C.; Richardson, C.D. Broad-spectrum antiviral activity of chebulagic acid and punicalagin against viruses that use glycosaminoglycans for entry. BMC Microbiol. 2013, 13, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, R.; Cooper, L.; Chen, Z.; Lee, H.; Rong, L.; Cui, Q. Discovery of chebulagic acid and punicalagin as novel allosteric inhibitors of SARS-CoV-2 3CLpro. Antivir. Res. 2021, 190, 105075. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, C.C.; Schang, L.M. A small molecule inhibits virion attachment to heparan sulfate- or sialic acid-containing glycans. J. Virol. 2014, 88, 7806–7817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.Y.; Shien, J.H.; Tiley, L.; Chiou, S.S.; Wang, S.Y.; Chang, T.J.; Lee, Y.J.; Chan, K.W.; Hsu, W.L. Curcumin inhibits influenza virus infection and haemagglutination activity. Food Chem. 2010, 119, 1346–1351. [Google Scholar] [CrossRef]
- Richart, S.M.; Li, Y.L.; Mizushina, Y.; Chang, Y.Y.; Chung, T.Y.; Chen, G.H.; Tzen, J.T.; Shia, K.S.; Hsu, W.L. Synergic effect of curcumin and its structural analogue (Monoacetylcurcumin) on anti-influenza virus infection. J. Food Drug Anal. 2018, 26, 1015–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.T.; Kuan, C.Y.; Liao, G.R.; Albrecht, R.A.; Tseng, Y.Y.; Hsu, Y.C.; Ou, S.C.; Hsu, W.L. A variant NS1 protein from H5N2 avian influenza virus suppresses PKR activation and promotes replication and virulence in mammals. Emerg. Microbes Infect. 2022, 11, 2291–2303. [Google Scholar] [CrossRef]
- Khanolkar, A.; Pewe, L.; Tifrea, D.; Perlman, S.; Harty, J.T. Developing MHV-1 infection as an animal model for Severe Acute Respiratory Syndrome (SARS). J. Immunol. 2007, 178, S42. [Google Scholar] [CrossRef]
- De Albuquerque, N.; Baig, E.; Ma, X.; Zhang, J.; He, W.; Rowe, A.; Habal, M.; Liu, M.; Shalev, I.; Downey, G.P.; et al. Murine hepatitis virus strain 1 produces a clinically relevant model of severe acute respiratory syndrome in A/J mice. J. Virol. 2006, 80, 10382–10394. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, E.-C.; Hsu, W.-L.; Tzen, J.T.C. Strictinin, a Major Ingredient in Yunnan Kucha Tea Possessing Inhibitory Activity on the Infection of Mouse Hepatitis Virus to Mouse L Cells. Molecules 2023, 28, 1080. https://doi.org/10.3390/molecules28031080
Tu E-C, Hsu W-L, Tzen JTC. Strictinin, a Major Ingredient in Yunnan Kucha Tea Possessing Inhibitory Activity on the Infection of Mouse Hepatitis Virus to Mouse L Cells. Molecules. 2023; 28(3):1080. https://doi.org/10.3390/molecules28031080
Chicago/Turabian StyleTu, Erh-Chuang, Wei-Li Hsu, and Jason T. C. Tzen. 2023. "Strictinin, a Major Ingredient in Yunnan Kucha Tea Possessing Inhibitory Activity on the Infection of Mouse Hepatitis Virus to Mouse L Cells" Molecules 28, no. 3: 1080. https://doi.org/10.3390/molecules28031080
APA StyleTu, E. -C., Hsu, W. -L., & Tzen, J. T. C. (2023). Strictinin, a Major Ingredient in Yunnan Kucha Tea Possessing Inhibitory Activity on the Infection of Mouse Hepatitis Virus to Mouse L Cells. Molecules, 28(3), 1080. https://doi.org/10.3390/molecules28031080