

(Magnetic) Cross-Linked Enzyme Aggregates of Cellulase from T. reesei: A Stable and Efficient Biocatalyst




Abstract
:
1. Introduction
2. Results and Discussions
2.1. Selection of Precipitation Reagent
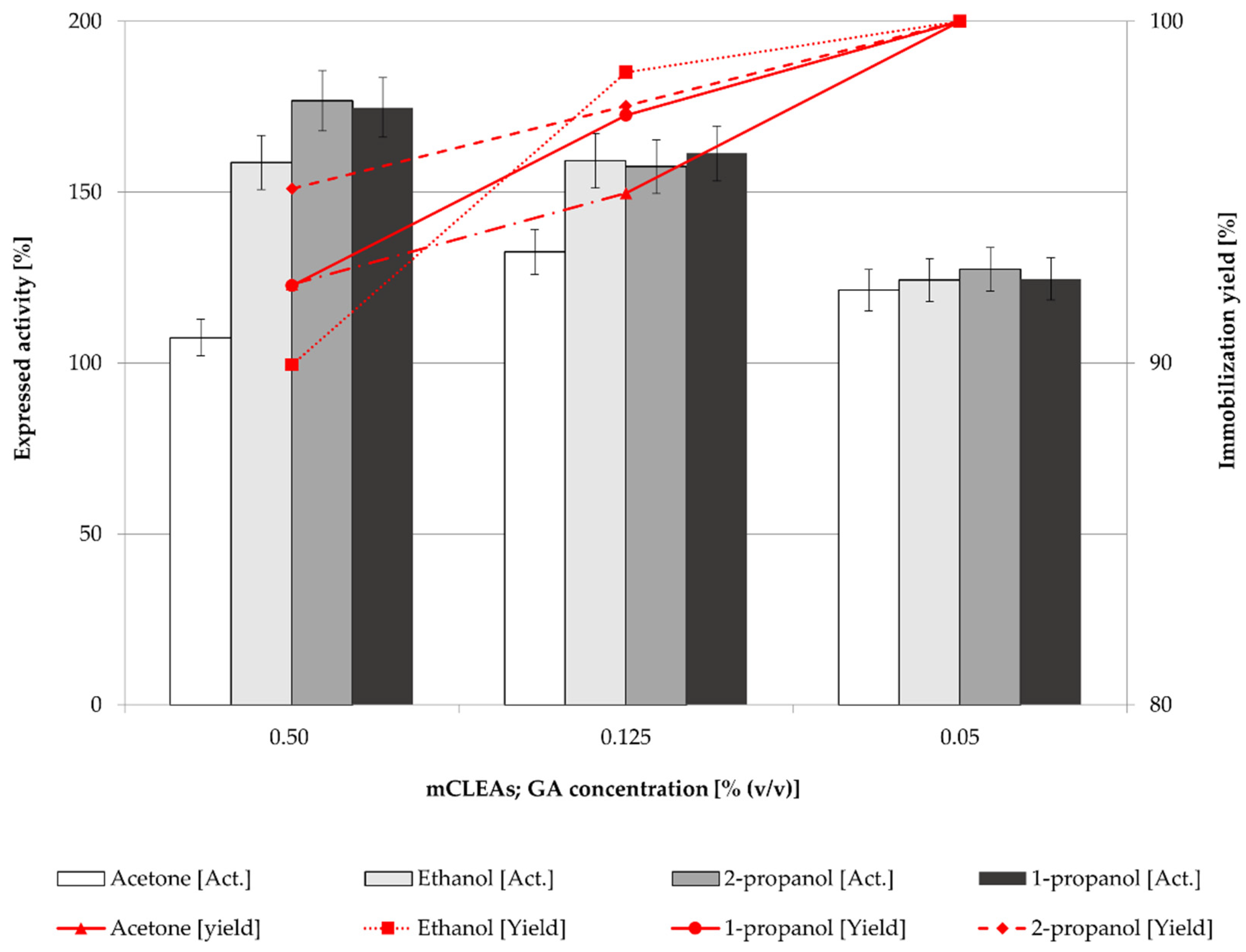
2.2. Effect of Cross-Linker (GA) Concentration
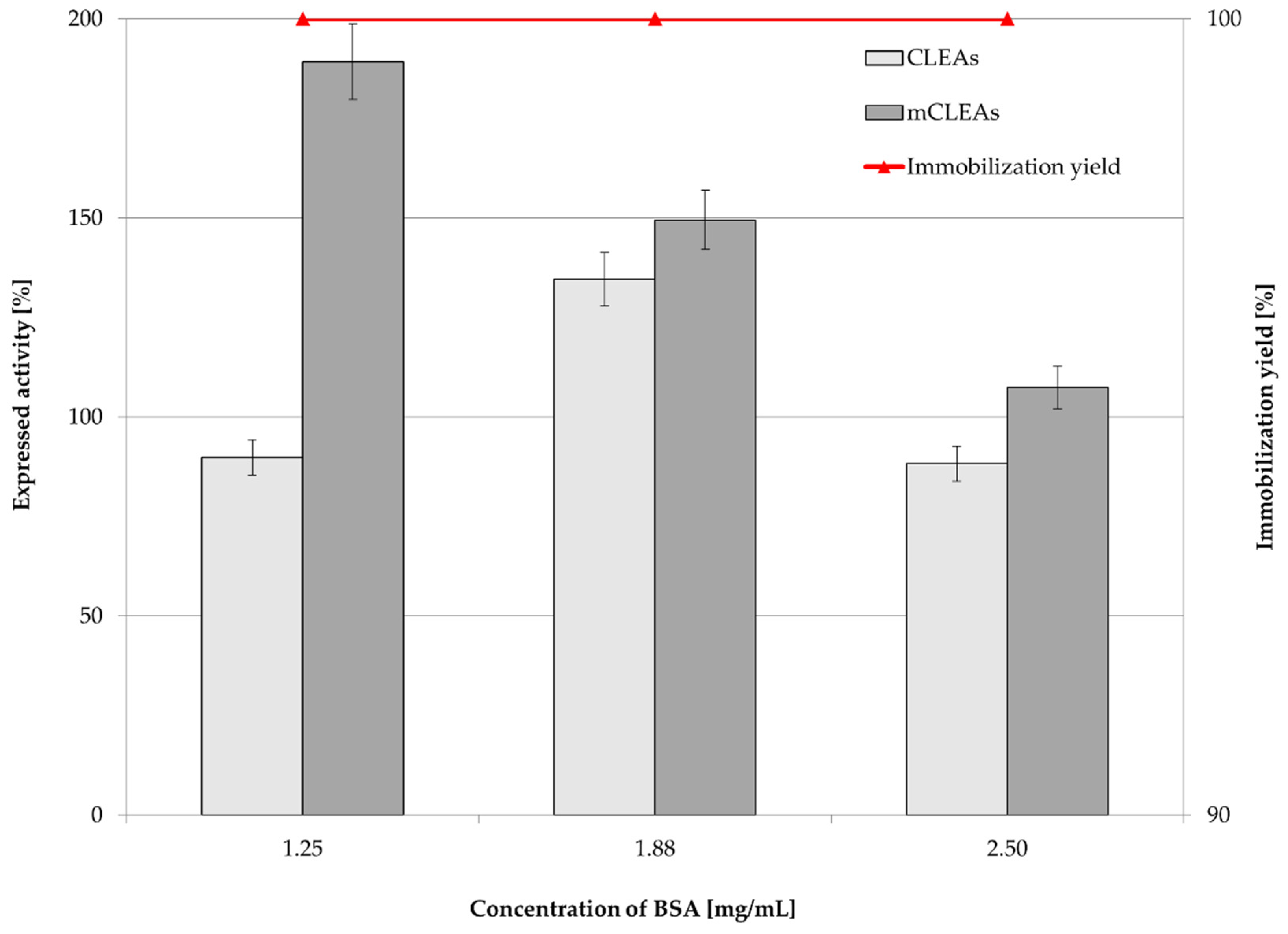
2.3. Effect of Proteic Feeder and Enzyme Concentration
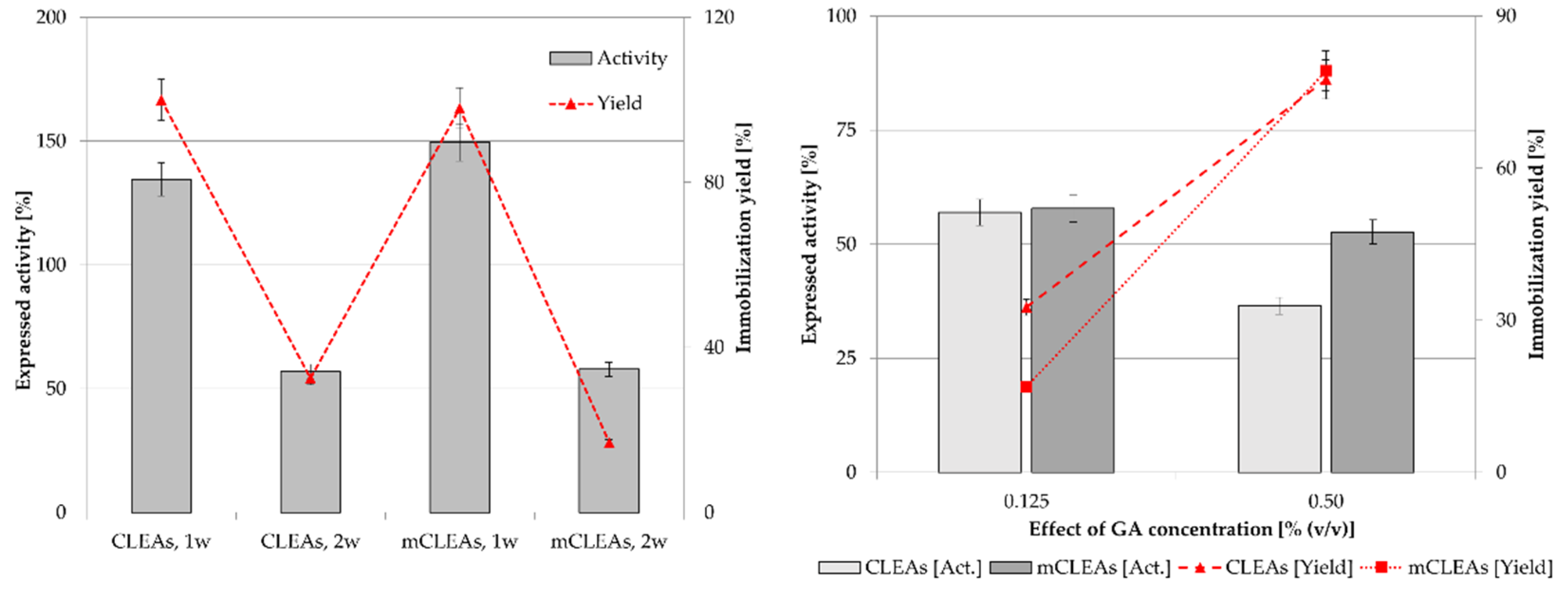
2.4. Effect of Increasing GA Concentration
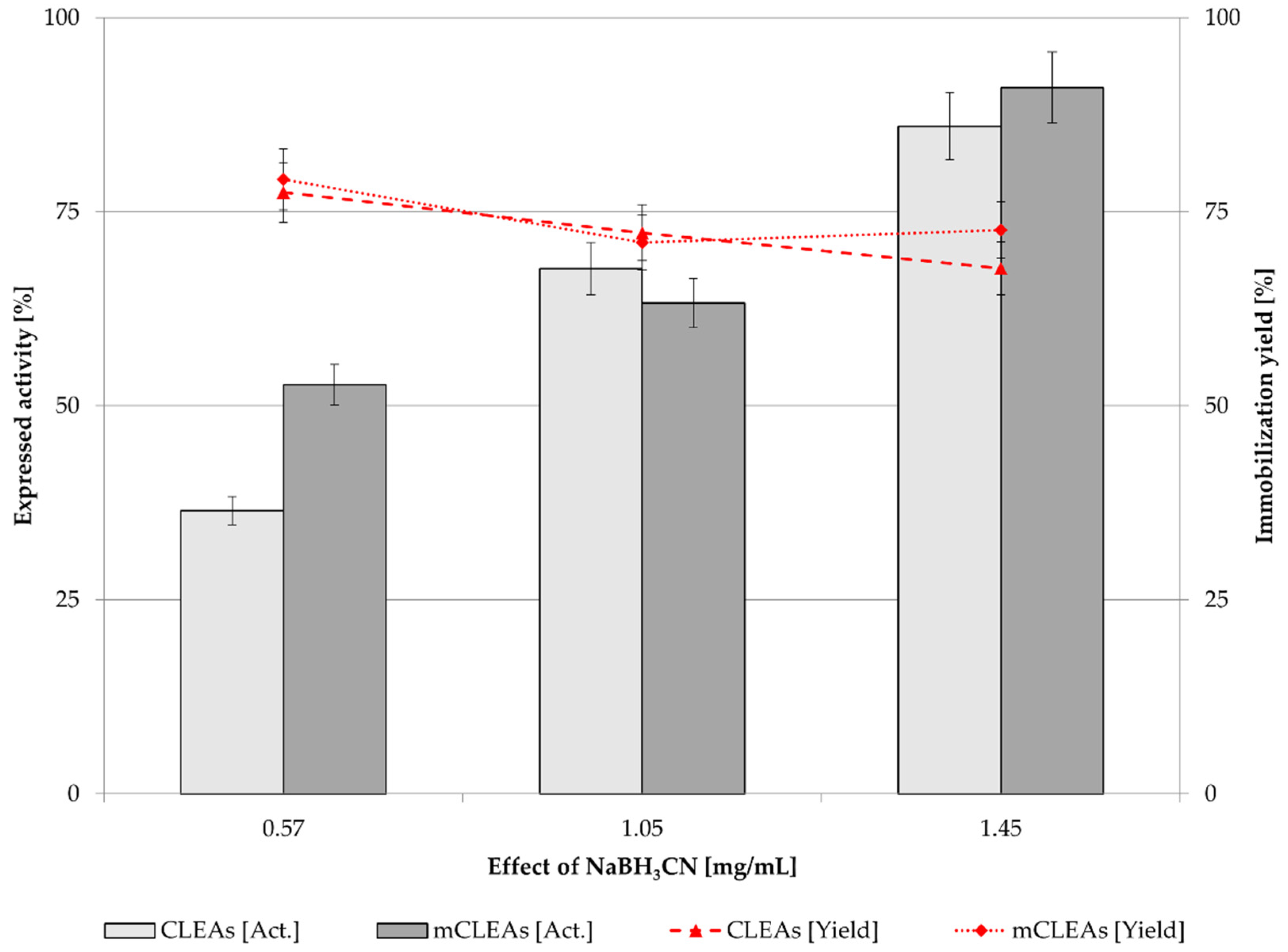
2.5. Effect of Reducing Agent Concentration
2.6. Reusability of CLEAs and mCLEAs
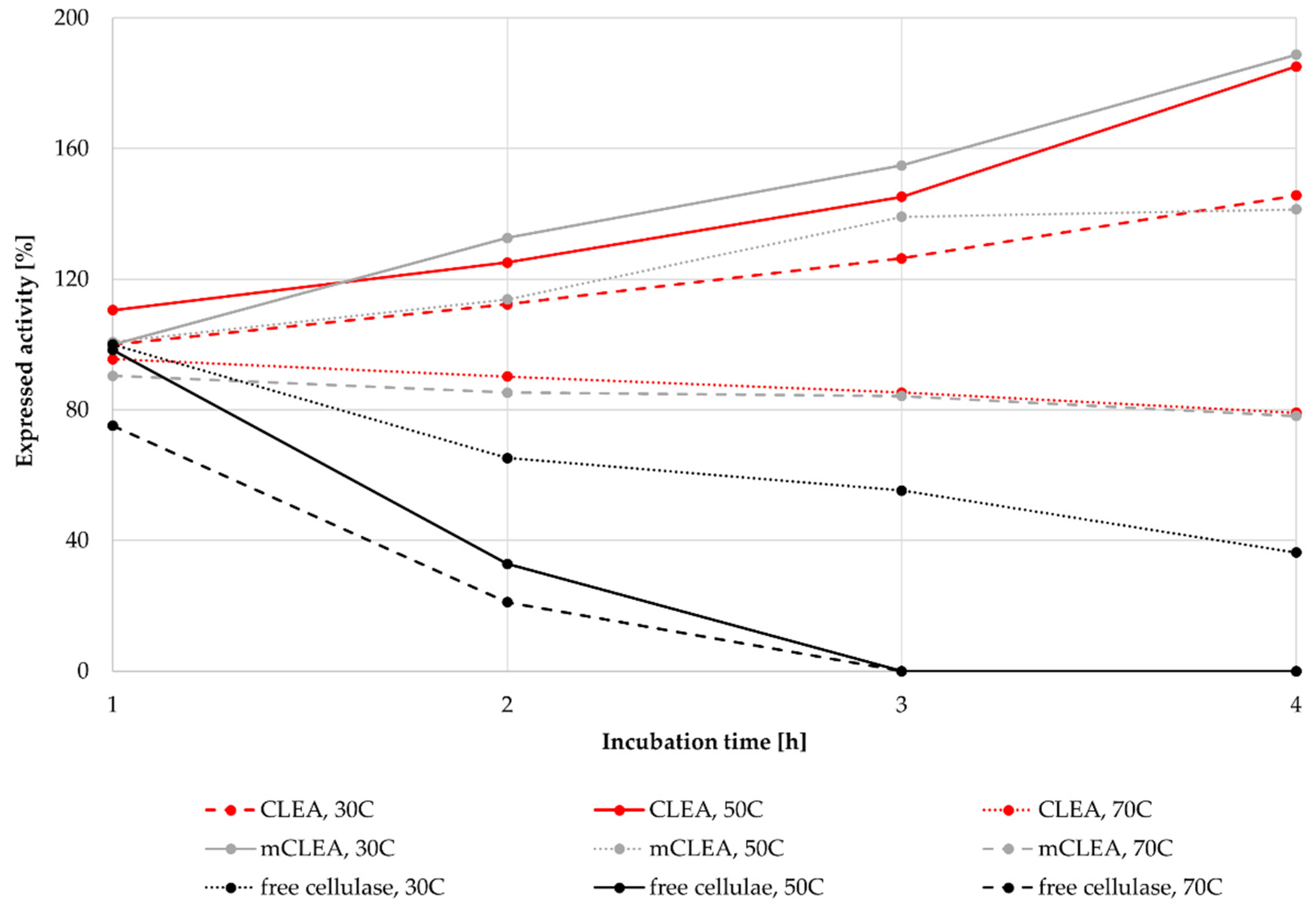
2.7. Thermal Stability of CLEAs and mCLEAs
2.8. Determination of Kinetic Parameters
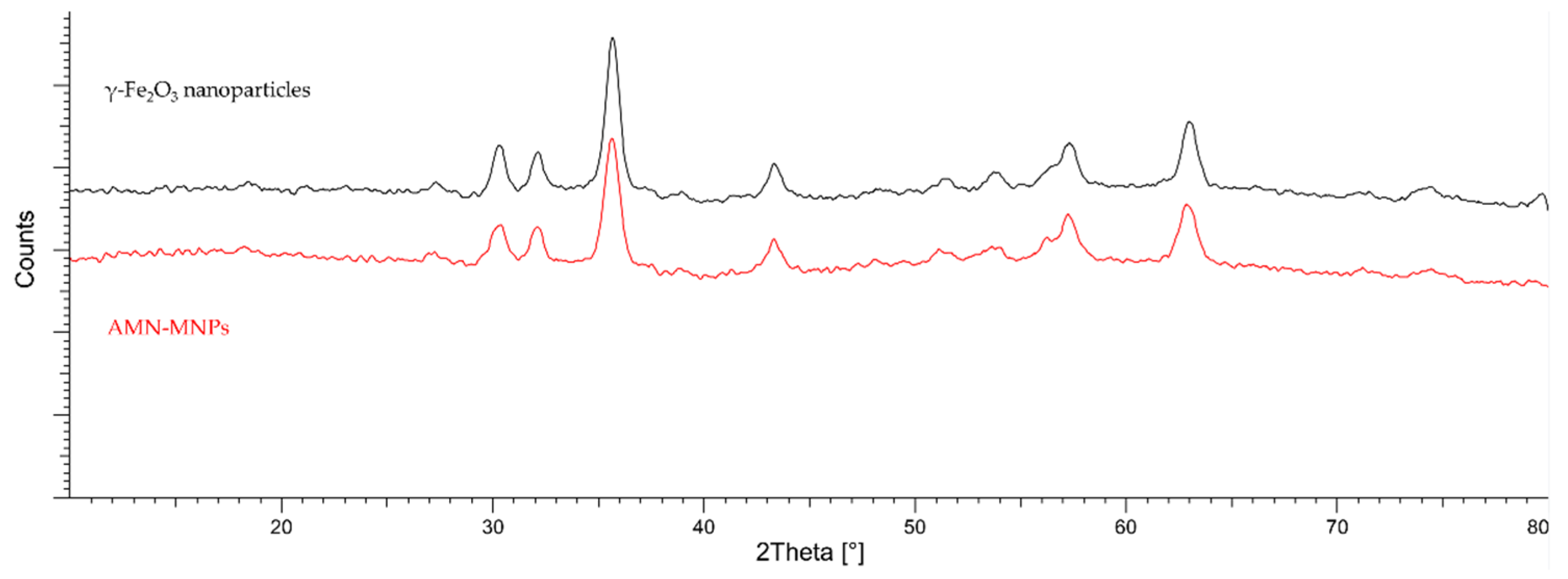
2.9. XRD Analysis
2.10. SEM Analysis
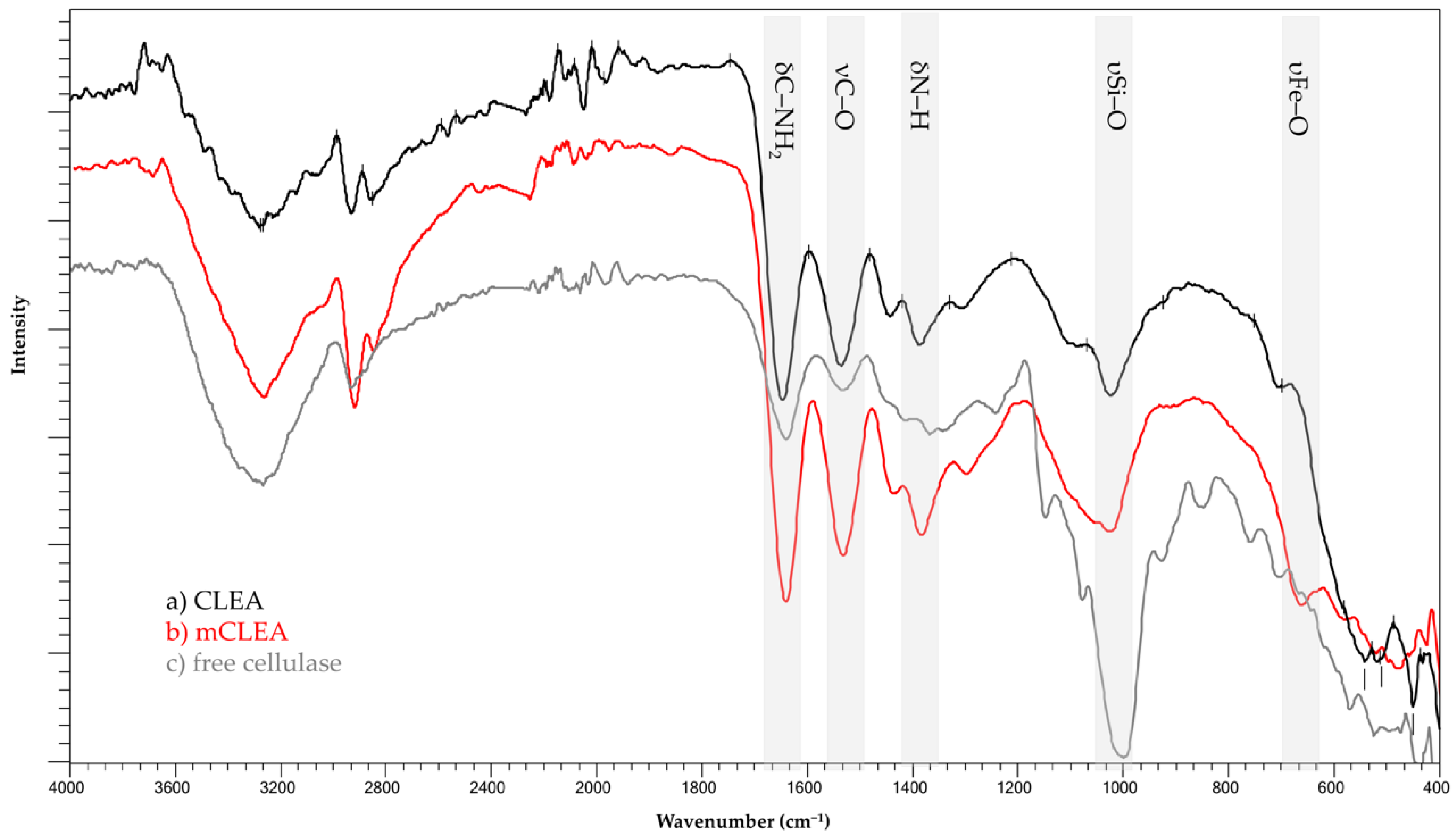
2.11. FT-IR Analysis
3. Materials and Methods
3.1. Materials
3.2. Preparation of Aminosilanized Magnetic Nanoparticles (AMN-MNPs)
3.3. Preparation of CLEAs and mCLEAs
3.3.1. Preparation of Enzyme Solution for CLEA
3.3.2. Preparation of Enzyme Solution for mCLEA
3.3.3. Enzyme Precipitation
3.3.4. Cross-Linking of CLEAs and mCLEAs
3.4. Determination of Immobilization Yield
- ci = concentration of cellulase initially used for reaction.
- cs = concentration of unbound cellulase, collected in the supernatant and in each purification (washing) cycle, respectively.
3.5. Cellulase Activity Assay
- activity of CLEAs or mCLEAs = activity of cross-linked cellulase or magnetic cross-linked cellulase measured by cellulase activity assay (U/mL).
- activity of free enzyme = activity of free cellulase measured by cellulase activity assay (U/mL).
3.6. Reusability Studies of Free Cellulase, CLEAs, and mCLEAs
3.7. Thermal Stability of Free Cellulase, CLEAs, and mCLEAs
3.8. Determination of Kinetic Parameters
3.9. X-ray Diffraction Analysis
3.10. Scanning Electron Microscopy Analysis
3.11. Fourier Transform Infrared Spectroscopy (FT-IR)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chen, N.; Chang, B.; Shi, N.; Yan, W.; Lu, F.; Liu, F. Cross-linked enzyme aggregates immobilization: Preparation, characterization, and applications. Crit. Rev. Biotechnol. 2022, 1–15. [Google Scholar] [CrossRef]
- Glasgow, E.; Meulen, K.V.; Kuch, N.; Fox, B.G. Multifunctional cellulases are potent, versatile tools for a renewable bioeconomy. Curr. Opin. Biotechnol. 2021, 67, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.P.; Himmel, M.E.; Mielenz, J.R. Outlook for cellulase improvement: Screening and selection strategies. Biotechnol. Adv. 2006, 24, 452–481. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Patel, S.K.S.; Gupta, R.K.; Otari, S.V.; Gao, H.; Lee, J.; Zhang, L. Enhanced Saccharification and Fermentation of Rice Straw by Reducing the Concentration of Phenolic Compounds Using an Immobilized Enzyme Cocktail. Biotechnol. J. 2019, 14, e1800468. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Wu, K.C.-W.; Saha, B. Emerging strategies for breaking the 3D amorphous network of lignin. Catal. Sci. Technol. 2014, 4, 3785–3799. [Google Scholar] [CrossRef]
- Dutta, S.; Wu, K.C.-W. Enzymatic breakdown of biomass: Enzyme active sites, immobilization, and biofuel production. Green Chem. 2014, 16, 4615–4626. [Google Scholar] [CrossRef]
- Rong, G.-X.; Lv, Z.-X.; Pan, Z.-J.; Zhang, S.-L.; Deng, P. Covalent Immobilization of Cellulase onto Amino and Graphene Oxide Functionalized Magnetic Fe2O3/Fe3O4@SiO2 Nanocomposites. J. Nanosci. Nanotechnol. 2021, 21, 4749–4757. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Das, D.; Kim, S.C.; Cho, B.-K.; Kalia, V.C.; Lee, J.-K. Integrating strategies for sustainable conversion of waste biomass into dark-fermentative hydrogen and value-added products. Renew. Sustain. Energy Rev. 2021, 150, 111491. [Google Scholar] [CrossRef]
- Patel, S.K.; Gupta, R.K.; Das, D.; Lee, J.-K.; Kalia, V.C. Continuous biohydrogen production from poplar biomass hydrolysate by a defined bacterial mixture immobilized on lignocellulosic materials under non-sterile conditions. J. Clean. Prod. 2021, 287, 125037. [Google Scholar] [CrossRef]
- Chi, M.-C.; Huang, Y.-F.; Lu, B.-Y.; Lin, M.-G.; Wang, T.-F.; Lin, L.-L. Magnetic Cross-Linked Enzyme Aggregates of a Transpeptidase-Specialized Variant (N450D) of Bacillus licheniformis γ-Glutamyl Transpeptidase: An Efficient and Stable Biocatalyst for l-Theanine Synthesis. Catalysts 2021, 11, 243. [Google Scholar] [CrossRef]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters necessary to define an immobilized enzyme preparation. Process. Biochem. 2019, 90, 66–80. [Google Scholar] [CrossRef]
- Morellon-Sterling, R.; Tavano, O.; Bolivar, J.M.; Berenguer-Murcia, Á.; Vela-Gutiérrez, G.; Sabir, J.S.; Tacias-Pascacio, V.G.; Fernandez-Lafuente, R. A review on the immobilization of pepsin: A Lys-poor enzyme that is unstable at alkaline pH values. Int. J. Biol. Macromol. 2022, 210, 682–702. [Google Scholar] [CrossRef]
- Sampaio, C.S.; Angelotti, J.A.; Fernandez-Lafuente, R.; Hirata, D.B. Lipase immobilization via cross-linked enzyme aggregates: Problems and prospects—A review. Int. J. Biol. Macromol. 2022, 215, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Castañeda-Valbuena, D.; Berenguer-Murcia, Á.; Kamli, M.R.; Tavano, O.; Fernandez-Lafuente, R. Immobilization of papain: A review. Int. J. Biol. Macromol. 2021, 188, 94–113. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Virgen-Ortííz, J.J.; dos Santos, J.C.S.; Berenguer-Murcia, Á.; Alcantara, A.R.; Barbosa, O.; Ortiz, C.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports: Immobilization mechanism, advantages, problems, and solutions. Biotechnol. Adv. 2019, 37, 746–770. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Strategies for the one-step immobilization–purification of enzymes as industrial biocatalysts. Biotechnol. Adv. 2015, 33, 435–456. [Google Scholar] [CrossRef] [Green Version]
- Cowan, D.A.; Fernandez-Lafuente, R. Enhancing the functional properties of thermophilic enzymes by chemical modification and immobilization. Enzym. Microb. Technol. 2011, 49, 326–346. [Google Scholar] [CrossRef]
- Hernandez, K.; Fernandez-Lafuente, R. Control of protein immobilization: Coupling immobilization and site-directed mutagenesis to improve biocatalyst or biosensor performance. Enzym. Microb. Technol. 2011, 48, 107–122. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Berenguer-Murcia, Á.; Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Stabilization of enzymes via immobilization: Multipoint covalent attachment and other stabilization strategies. Biotechnol. Adv. 2021, 52, 107821. [Google Scholar] [CrossRef]
- Kalyankar, P.; Zhu, Y.; Keeffe, M.O.; Cuinn, G.O.; FitzGerald, R.J. Substrate specificity of glutamyl endopeptidase (GE): Hydrolysis studies with a bovine α-casein preparation. Food Chem. 2013, 136, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and Biotechnological Aspects of Microbial Proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butré, C.I.; Sforza, S.; Gruppen, H.; Wierenga, P.A. Introducing enzyme selectivity: A quantitative parameter to describe enzymatic protein hydrolysis. Anal. Bioanal. Chem. 2014, 406, 5827–5841. [Google Scholar] [CrossRef] [PubMed]
- Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. Immobilization as a Strategy for Improving Enzyme Properties-Application to Oxidoreductases. Molecules 2014, 19, 8995–9018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.K.; Anwar, M.Z.; Kumar, A.; Otari, S.V.; Pagolu, R.T.; Kim, S.-Y.; Kim, I.-W.; Lee, J.-K. Fe2O3 yolk-shell particle-based laccase biosensor for efficient detection of 2,6-dimethoxyphenol. Biochem. Eng. J. 2018, 132, 1–8. [Google Scholar] [CrossRef]
- Patel, S.K.; Otari, S.V.; Li, J.; Kim, D.R.; Kim, S.C.; Cho, B.-K.; Kalia, V.C.; Kang, Y.C.; Lee, J.-K. Synthesis of cross-linked protein-metal hybrid nanoflowers and its application in repeated batch decolorization of synthetic dyes. J. Hazard. Mater. 2018, 347, 442–450. [Google Scholar] [CrossRef]
- Khoshnevisan, K.; Bordbar, A.-K.; Zare, D.; Davoodi, D.; Noruzi, M.; Barkhi, M.; Tabatabaei, M. Immobilization of cellulase enzyme on superparamagnetic nanoparticles and determination of its activity and stability. Chem. Eng. J. 2011, 171, 669–673. [Google Scholar] [CrossRef]
- Roessl, U.; Nahalka, J.; Nidetzky, B. Carrier-free immobilized enzymes for biocatalysis. Biotechnol. Lett. 2010, 32, 341–350. [Google Scholar] [CrossRef]
- Sheldon, R.A. Enzyme Immobilization: The Quest for Optimum Performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Schoevaart, R.; Wolbers, M.; Golubovic, M.; Ottens, M.; Kieboom, A.; van Rantwijk, F.; van der Wielen, L.; Sheldon, R. Preparation, optimization, and structures of cross-linked enzyme aggregates (CLEAs). Biotechnol. Bioeng. 2004, 87, 754–762. [Google Scholar] [CrossRef]
- Ju, H.; Jang, E.; Ryu, B.H.; Kim, T.D. Characterization and preparation of highly stable aggregates of a novel type of hydrolase (BL28) from Bacillus licheniformis. Bioresour. Technol. 2013, 128, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Li, C.; Jiao, X.; Jia, S.; Jiang, Y.; Bilal, M.; Cui, J. Recent progress in multienzymes co-immobilization and multienzyme system applications. Chem. Eng. J. 2019, 373, 1254–1278. [Google Scholar] [CrossRef]
- Mehde, A.A. Development of magnetic cross-linked peroxidase aggregates on starch as enhancement template and their application for decolorization. Int. J. Biol. Macromol. 2019, 131, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Cheng, H.; Chen, G.; Ju, F.; Fernández-Lucas, J.; Zdarta, J.; Jesionowski, T.; Bilal, M. Designing multifunctional biocatalytic cascade system by multi-enzyme co-immobilization on biopolymers and nanostructured materials. Int. J. Biol. Macromol. 2023, 227, 535–550. [Google Scholar] [CrossRef]
- Sheldon, R.A. CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts 2019, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Del Arco, J.; Del Arco, J.; Alcántara, A.R.; Alcántara, A.R.; Fernández-Lafuente, R.; Fernández-Lafuente, R.; Fernández-Lucas, J.; Fernández-Lucas, J. Magnetic micro-macro biocatalysts applied to industrial bioprocesses. Bioresour. Technol. 2021, 322, 124547. [Google Scholar] [CrossRef]
- Cruz, J.; Barbosa, O.; Rodrigues, R.C.; Fernandez-Lafuente, R.; Torres, R.; Ortiz, C. Optimized preparation of CALB-CLEAs by response surface methodology: The necessity to employ a feeder to have an effective crosslinking. J. Mol. Catal. B Enzym. 2012, 80, 7–14. [Google Scholar] [CrossRef]
- López-Gallego, F.; Betancor, L.; Hidalgo, A.; Alonso, N.; Fernández-Lafuente, R.; Guisán, J.M. Co-aggregation of Enzymes and Polyethyleneimine: A Simple Method to Prepare Stable and Immobilized Derivatives of Glutaryl Acylase. Biomacromolecules 2005, 6, 1839–1842. [Google Scholar] [CrossRef]
- Shah, S.; Sharma, A.; Gupta, M.N. Preparation of cross-linked enzyme aggregates by using bovine serum albumin as a proteic feeder. Anal. Biochem. 2006, 351, 207–213. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Pletschke, B.I. Magnetic cross-linked enzyme aggregates (CLEAs): A novel concept towards carrier free immobilization of lignocellulolytic enzymes. Enzym. Microb. Technol. 2014, 61–62, 17–27. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Choi, S.H.; Kang, Y.C.; Lee, J.-K. Eco-Friendly Composite of Fe3O4-Reduced Graphene Oxide Particles for Efficient Enzyme Immobilization. ACS Appl. Mater. Interfaces 2017, 9, 2213–2222. [Google Scholar] [CrossRef] [PubMed]
- Arana-Peña, S.; Carballares, D.; Morellon-Sterlling, R.; Berenguer-Murcia, Á.; Alcántara, A.R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Enzyme co-immobilization: Always the biocatalyst designers’ choice…or not? Biotechnol. Adv. 2020, 51, 107584. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Hussain, N.; Américo-Pinheiro, J.H.P.; Almulaiky, Y.Q.; Iqbal, H.M. Multi-enzyme co-immobilized nano-assemblies: Bringing enzymes together for expanding bio-catalysis scope to meet biotechnological challenges. Int. J. Biol. Macromol. 2021, 186, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, R.C.; Gupta, R.; Singh, A. Microbial Cellulases and Their Industrial Applications. Available online: https://www.hindawi.com/journals/er/2011/280696/ (accessed on 9 April 2018).
- Rodríguez-Zúñiga, U.F.; Cannella, D.; Giordano, R.D.C.; Giordano, R.D.L.C.; Jørgensen, H.; Felby, C. Lignocellulose pretreatment technologies affect the level of enzymatic cellulose oxidation by LPMO. Green Chem. 2015, 17, 2896–2903. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.K.; Kim, Y.; Ximenes, E.; Ladisch, M.R. Effect of liquid hot water pretreatment severity on properties of hardwood lignin and enzymatic hydrolysis of cellulose. Biotechnol. Bioeng. 2014, 112, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xue, Y.; Wu, X. Characterization of a Novel Thermostable Chitin-Binding Domain and Its Application in Immobilization of a Multifunctional Hemicellulase. J. Agric. Food Chem. 2013, 61, 3074–3081. [Google Scholar] [CrossRef]
- Hartmann, M.; Kostrov, X. Immobilization of enzymes on porous silicas—Benefits and challenges. Chem. Soc. Rev. 2013, 42, 6277–6289. [Google Scholar] [CrossRef]
- Yuan, Y.; Luan, X.; Rana, X.; Hassan, M.E.; Dou, D. Covalent immobilization of cellulase in application of biotransformation of ginsenoside Rb1. J. Mol. Catal. B Enzym. 2016, 133, S525–S532. [Google Scholar] [CrossRef]
- Cui, J.; Cui, L.; Jia, S.; Su, Z.; Zhang, S. Hybrid Cross-Linked Lipase Aggregates with Magnetic Nanoparticles: A Robust and Recyclable Biocatalysis for the Epoxidation of Oleic Acid. J. Agric. Food Chem. 2016, 64, 7179–7187. [Google Scholar] [CrossRef]
- Tang, C.; Saquing, C.D.; Morton, S.W.; Glatz, B.N.; Kelly, R.M.; Khan, S.A. Cross-linked Polymer Nanofibers for Hyperthermophilic Enzyme Immobilization: Approaches to Improve Enzyme Performance. ACS Appl. Mater. Interfaces 2014, 6, 11899–11906. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.H.-Y.; Jang, J.; Wu, K.C.-W. Cellulase immobilized mesoporous silica nanocatalysts for efficient cellulose-to-glucose conversion. Green Chem. 2011, 13, 2844. [Google Scholar] [CrossRef]
- Pota, G.; Salerno, A.S.; Costantini, A.; Silvestri, B.; Passaro, J.; Califano, V. Co-immobilization of Cellulase and β-Glucosidase into Mesoporous Silica Nanoparticles for the Hydrolysis of Cellulose Extracted from Eriobotrya japonica Leaves. Langmuir 2022, 38, 5481–5493. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-C.; Dutta, S.; Wu, K.C.-W. Integrated, Cascading Enzyme-/Chemocatalytic Cellulose Conversion using Catalysts based on Mesoporous Silica Nanoparticles. Chemsuschem 2014, 7, 3241–3246. [Google Scholar] [CrossRef]
- Ozyilmaz, E.; Alhiali, A.; Caglar, O.; Yilmaz, M. Preparation of regenerable magnetic nanoparticles for cellulase immobilization: Improvement of enzymatic activity and stability. Biotechnol. Prog. 2021, 37, e3145. [Google Scholar] [CrossRef]
- Desai, M.P.; Pawar, K.D. Immobilization of cellulase on iron tolerant Pseudomonas stutzeri biosynthesized photocatalytically active magnetic nanoparticles for increased thermal stability. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 106, 110169. [Google Scholar] [CrossRef]
- Huang, W.; Pan, S.; Li, Y.; Yu, L.; Liu, R. Immobilization and characterization of cellulase on hydroxy and aldehyde functionalized magnetic Fe2O3/Fe3O4 nanocomposites prepared via a novel rapid combustion process. Int. J. Biol. Macromol. 2020, 162, 845–852. [Google Scholar] [CrossRef]
- Perwez, M.; Mazumder, J.; Sardar, M. Preparation and characterization of reusable magnetic combi-CLEA of cellulase and hemicellulase. Enzym. Microb. Technol. 2019, 131, 109389. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gong, X.; Yang, G.; Li, Q.; Huang, J.; Zhou, N.; Jia, X. Cross-linked enzyme aggregates (CLEAs) of cellulase with improved catalytic activity, adaptability and reusability. Bioprocess Biosyst. Eng. 2022, 45, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, S.; Liu, X. An overview of algae bioethanol production. Int. J. Energy Res. 2014, 38, 965–977. [Google Scholar] [CrossRef]
- Pérez de los Ríos, A.; Hernández-Fernández, F.J.; Henríquez, P.A.Z.; Missoun, F.; Hernández-Fernández, J.; Ortiz-Martínez, V.; Salar-García, M.J.; Blanco, L.J.L.; Godínez, C. Keys for Bioethanol Production Processes by Fermentation and Ionic Liquid Extraction. ACS Sustain. Chem. Eng. 2017, 5, 6986–6993. [Google Scholar] [CrossRef]
- Podrepšek, G.H.; Knez, Ž.; Leitgeb, M. Different preparation methods and characterization of magnetic maghemite coated with chitosan. J. Nanoparticle Res. 2013, 15, 1–12. [Google Scholar] [CrossRef]
- Leitgeb, M.; Heržič, K.; Podrepšek, G.H.; Hojski, A.; Crnjac, A.; Knez, Z. Toxicity of magnetic chitosan micro and nanoparticles as carriers for biologically active substances. Acta Chim. Slov. 2014, 61, 145–152. [Google Scholar] [PubMed]
- Križnik, L.; Vasić, K.; Knez, Ž.; Leitgeb, M. Hyper-activation of ß-galactosidase from Aspergillus oryzae via immobilization onto amino-silane and chitosan magnetic maghemite nanoparticles. J. Clean. Prod. 2018, 179, 225–234. [Google Scholar] [CrossRef]
- Peirce, S.; Russo, M.E.; Isticato, R.; Lafuente, R.F.; Salatino, P.; Marzocchella, A. Structure and activity of magnetic cross-linked enzyme aggregates of bovine carbonic anhydrase as promoters of enzymatic CO 2 capture. Biochem. Eng. J. 2017, 127, 188–195. [Google Scholar] [CrossRef]
- Agyei, D.; He, L. Evaluation of cross-linked enzyme aggregates of Lactobacillus cell-envelope proteinases, for protein degradation. Food Bioprod. Process. 2015, 94, 59–69. [Google Scholar] [CrossRef]
- Chen, N.; Chang, B.; Shi, N.; Lu, F.; Liu, F. Robust and recyclable cross-linked enzyme aggregates of sucrose isomerase for isomaltulose production. Food Chem. 2023, 399, 134000. [Google Scholar] [CrossRef]
- Rahman, N.H.A.; Jaafar, N.R.; Murad, A.M.A.; Abu Bakar, F.D.; Annuar, N.A.S.; Illias, R.M. Novel cross-linked enzyme aggregates of levanase from Bacillus lehensis G1 for short-chain fructooligosaccharides synthesis: Developmental, physicochemical, kinetic and thermodynamic properties. Int. J. Biol. Macromol. 2020, 159, 577–589. [Google Scholar] [CrossRef]
- Jannat, M.; Yang, K.-L. A Millifluidic Device with Embedded Cross-Linked Enzyme Aggregates for Degradation of H2O2. ACS Appl. Mater. Interfaces 2020, 12, 6768–6775. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Yang, K.-L. Uniform cross-linked cellulase aggregates prepared in millifluidic reactors. J. Colloid Interface Sci. 2014, 428, 146–151. [Google Scholar] [CrossRef]
- Amadi, O.; Awodiran, I.; Moneke, A.; Nwagu, T.; Egong, J.; Chukwu, G. Concurrent production of cellulase, xylanase, pectinase and immobilization by combined Cross-linked enzyme aggregate strategy- advancing tri-enzyme biocatalysis. Bioresour. Technol. Rep. 2022, 18, 101019. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (CLEAs). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Jung, D.-H.; Jung, J.H.; Seo, D.-H.; Ha, S.-J.; Kweon, D.-K.; Park, C.-S. One-pot bioconversion of sucrose to trehalose using enzymatic sequential reactions in combined cross-linked enzyme aggregates. Bioresour. Technol. 2013, 130, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Lucena, G.N.; dos Santos, C.C.; Pinto, G.C.; Piazza, R.D.; Guedes, W.; Júnior, M.J.; de Paula, A.V.; Marques, R.F.C. Synthesis and characterization of magnetic cross-linked enzyme aggregate and its evaluation of the alternating magnetic field (AMF) effects in the catalytic activity. J. Magn. Magn. Mater. 2020, 516, 167326. [Google Scholar] [CrossRef]
- Gajšek, M.; Jančič, U.; Vasić, K.; Knez, Ž.; Leitgeb, M. Enhanced activity of immobilized transglutaminase for cleaner production technologies. J. Clean. Prod. 2019, 240, 118218. [Google Scholar] [CrossRef]
- Torres, M.G.; Foresti, M.; Ferreira, M. Effect of different parameters on the hydrolytic activity of cross-linked enzyme aggregates (CLEAs) of lipase from Thermomyces lanuginosa. Biochem. Eng. J. 2013, 72, 18–23. [Google Scholar] [CrossRef]
- Galvis, M.; Barbosa, O.; Ruiz, M.; Cruz, J.; Ortiz, C.; Torres, R.; Fernandez-Lafuente, R. Chemical amination of lipase B from Candida antarctica is an efficient solution for the preparation of crosslinked enzyme aggregates. Process. Biochem. 2012, 47, 2373–2378. [Google Scholar] [CrossRef]
- Yu, H.; Chen, H.; Wang, X.; Yang, Y.; Ching, C. Cross-linked enzyme aggregates (CLEAs) with controlled particles: Application to Candida rugosa lipase. J. Mol. Catal. B Enzym. 2006, 43, 124–127. [Google Scholar] [CrossRef]
- Li, B.; Dong, S.L.; Xie, X.L.; Xu, Z.B.; Li, L. Preparation and Properties of Cross-Linked Enzyme Aggregates of Cellulase. Adv. Mater. Res. 2012, 581–582, 257–260. [Google Scholar] [CrossRef]
- Abraham, R.E.; Verma, M.L.; Barrow, C.J.; Puri, M. Suitability of magnetic nanoparticle immobilised cellulases in enhancing enzymatic saccharification of pretreated hemp biomass. Biotechnol. Biofuels 2014, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Eldin, M.M.; EL-Aassar, M.R.; EL-Zatahry, A.A.; EL-Sabbah, M.M.B. β-Galactosidase Immobilization onto Poly (Acrylonitrile-Co-Methyl Methacrylate) Nanoparticles. Trends Biomater. Artif. Organs. 2015, 29, 193–200. [Google Scholar]
- Mateo, C.; Palomo, J.M.; van Langen, L.M.; van Rantwijk, F.; Sheldon, R.A. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol. Bioeng. 2004, 86, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.; Mauguen, Y.; Prangé, T. Revisiting glutaraldehyde cross-linking: The case of the Arg–Lys intermolecular doublet. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Šulek, F.; Fernández, D.P.; Knez, Ž.; Habulin, M.; Sheldon, R.A. Immobilization of horseradish peroxidase as crosslinked enzyme aggregates (CLEAs). Process. Biochem. 2011, 46, 765–769. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, G.; Chougle, R.; Nainegali, B.; Desai, S.; Joshi, A.; Kambale, S.; Kamat, P.; Haripurkar, R.; Jadhav, S.; et al. Preparation of stable cross-linked enzyme aggregates (CLEAs) of NADH-dependent nitrate reductase and its use for silver nanoparticle synthesis from silver nitrate. Catal. Commun. 2014, 53, 62–66. [Google Scholar] [CrossRef]
- Wang, H.; Huang, J.; Wang, C.; Li, D.; Ding, L.; Han, Y. Immobilization of glucose oxidase using CoFe2O4/SiO2 nanoparticles as carrier. Appl. Surf. Sci. 2011, 257, 5739–5745. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, W.; Yang, Z.; Yang, X.; Wang, N.; Yu, X. Novel Magnetic Cross-Linked Cellulase Aggregates with a Potential Application in Lignocellulosic Biomass Bioconversion. Molecules 2017, 22, 269. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEA® s): Stable and recyclable biocatalysts. Biochem. Soc. Trans. 2007, 35, 1583–1587. [Google Scholar] [CrossRef] [Green Version]
- Khorshidi, K.J.; Lenjannezhadian, H.; Jamalan, M.; Zeinali, M. Preparation and characterization of nanomagnetic cross-linked cellulase aggregates for cellulose bioconversion. J. Chem. Technol. Biotechnol. 2014, 91, 539–546. [Google Scholar] [CrossRef]
- Tufvesson, P.; Fu, W.; Jensen, J.S.; Woodley, J. Process considerations for the scale-up and implementation of biocatalysis. Food Bioprod. Process. 2010, 88, 3–11. [Google Scholar] [CrossRef]
- Basso, A.; Serban, S. Industrial applications of immobilized enzymes—A review. Mol. Catal. 2019, 479, 110607. [Google Scholar] [CrossRef]
- Gokhale, A.A.; Lu, J.; Lee, I. Immobilization of cellulase on magnetoresponsive graphene nano-supports. J. Mol. Catal. B Enzym. 2013, 90, 76–86. [Google Scholar] [CrossRef]
- Kumar, S.; Mohan, U.; Kamble, A.L.; Pawar, S.; Banerjee, U.C. Cross-linked enzyme aggregates of recombinant Pseudomonas putida nitrilase for enantioselective nitrile hydrolysis. Bioresour. Technol. 2010, 101, 6856–6858. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Cao, M.; Wen, H.; Tan, Z.; Jia, S.; Cui, J. Biodegradation of polyvinyl alcohol using cross-linked enzyme aggregates of degrading enzymes from Bacillus niacini. Int. J. Biol. Macromol. 2019, 124, 10–16. [Google Scholar] [CrossRef]
- Shuddhodana; Gupta, M.N.; Bisaria, V.S. Effectiveness of cross-linked enzyme aggregates of cellulolytic enzymes in hydrolyzing wheat straw. J. Biosci. Bioeng. 2018, 126, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, B.; Pal, A.; Jain, V. Improved Enzyme Catalytic Characteristics upon Glutaraldehyde Cross-Linking of Alginate Entrapped Xylanase Isolated from Aspergillus flavus MTCC 9390. Enzym. Res. 2015, 2015, 210784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šulek, F.; Drofenik, M.; Habulin, M.; Knez, Ž. Surface functionalization of silica-coated magnetic nanoparticles for covalent attachment of cholesterol oxidase. J. Magn. Magn. Mater. 2010, 322, 179–185. [Google Scholar] [CrossRef]
- Sánchez-Ramírez, J.; Martínez-Hernández, J.L.; Segura-Ceniceros, P.; López, G.; Saade, H.; Medina-Morales, M.A.; Ramos-González, R.; Aguilar, C.N.; Ilyina, A. Cellulases immobilization on chitosan-coated magnetic nanoparticles: Application for Agave Atrovirens lignocellulosic biomass hydrolysis. Bioprocess Biosyst. Eng. 2016, 40, 9–22. [Google Scholar] [CrossRef]
- Bohara, R.A.; Thorat, N.D.; Pawar, S.H. Immobilization of cellulase on functionalized cobalt ferrite nanoparticles. Korean J. Chem. Eng. 2015, 33, 216–222. [Google Scholar] [CrossRef]
- Zang, L.; Qiu, J.; Wu, X.; Zhang, W.; Sakai, E.; Wei, Y. Preparation of Magnetic Chitosan Nanoparticles as Support for Cellulase Immobilization. Ind. Eng. Chem. Res. 2014, 53, 3448–3454. [Google Scholar] [CrossRef]
- Šulek, F.; Knez, Ž.; Habulin, M. Immobilization of cholesterol oxidase to finely dispersed silica-coated maghemite nanoparticles based magnetic fluid. Appl. Surf. Sci. 2010, 256, 4596–4600. [Google Scholar] [CrossRef]
- Podrepsek, G.H.; Primozic, M.; Knez, Z.; Habulin, M. Immobilization of cellulase for industrial production. Chem. Eng. 2012, 27, 235–240. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Storms, R.; Tsang, A. Microplate-based carboxymethylcellulose assay for endoglucanase activity. Anal. Biochem. 2005, 342, 176–178. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activity CLEAs | Activity mCLEAs | Yield CLEAs | Yield mCLEAs | |
|---|---|---|---|---|
| 0.50% (w/w) GA | 181.71 | 174.79 | 99.76 | 99.69 |
| 0.125% (w/w) GA | 152.34 | 161.33 | 99.96 | 99.89 |
| 0.05% (w/w) GA | 87.15 | 124.63 | 100 | 100 |
| Sample | Vmax [µmol/min] | KM [mM] |
|---|---|---|
| Free cellulase | 8.86 ± 0.0041 | 0.012 ± 0.0018 |
| CLEAs | 1.12 ± 0.0012 | 0.055 ± 0.0102 |
| mCLEAs | 1.71 ± 0.0023 | 0.037 ± 0.0012 |
| Sample | dXRD [nm] |
|---|---|
| γ-Fe2O3 | 11.2 |
| AMN-MNPS | 12.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ifko, D.; Vasić, K.; Knez, Ž.; Leitgeb, M. (Magnetic) Cross-Linked Enzyme Aggregates of Cellulase from T. reesei: A Stable and Efficient Biocatalyst. Molecules 2023, 28, 1305. https://doi.org/10.3390/molecules28031305
Ifko D, Vasić K, Knez Ž, Leitgeb M. (Magnetic) Cross-Linked Enzyme Aggregates of Cellulase from T. reesei: A Stable and Efficient Biocatalyst. Molecules. 2023; 28(3):1305. https://doi.org/10.3390/molecules28031305
Chicago/Turabian StyleIfko, Dušica, Katja Vasić, Željko Knez, and Maja Leitgeb. 2023. "(Magnetic) Cross-Linked Enzyme Aggregates of Cellulase from T. reesei: A Stable and Efficient Biocatalyst" Molecules 28, no. 3: 1305. https://doi.org/10.3390/molecules28031305
APA StyleIfko, D., Vasić, K., Knez, Ž., & Leitgeb, M. (2023). (Magnetic) Cross-Linked Enzyme Aggregates of Cellulase from T. reesei: A Stable and Efficient Biocatalyst. Molecules, 28(3), 1305. https://doi.org/10.3390/molecules28031305