


Analysis of the Qualitative and Quantitative Content of the Phenolic Compounds of Selected Moss Species under NaCl Stress

 , ,
, ,  ,
,  and
and 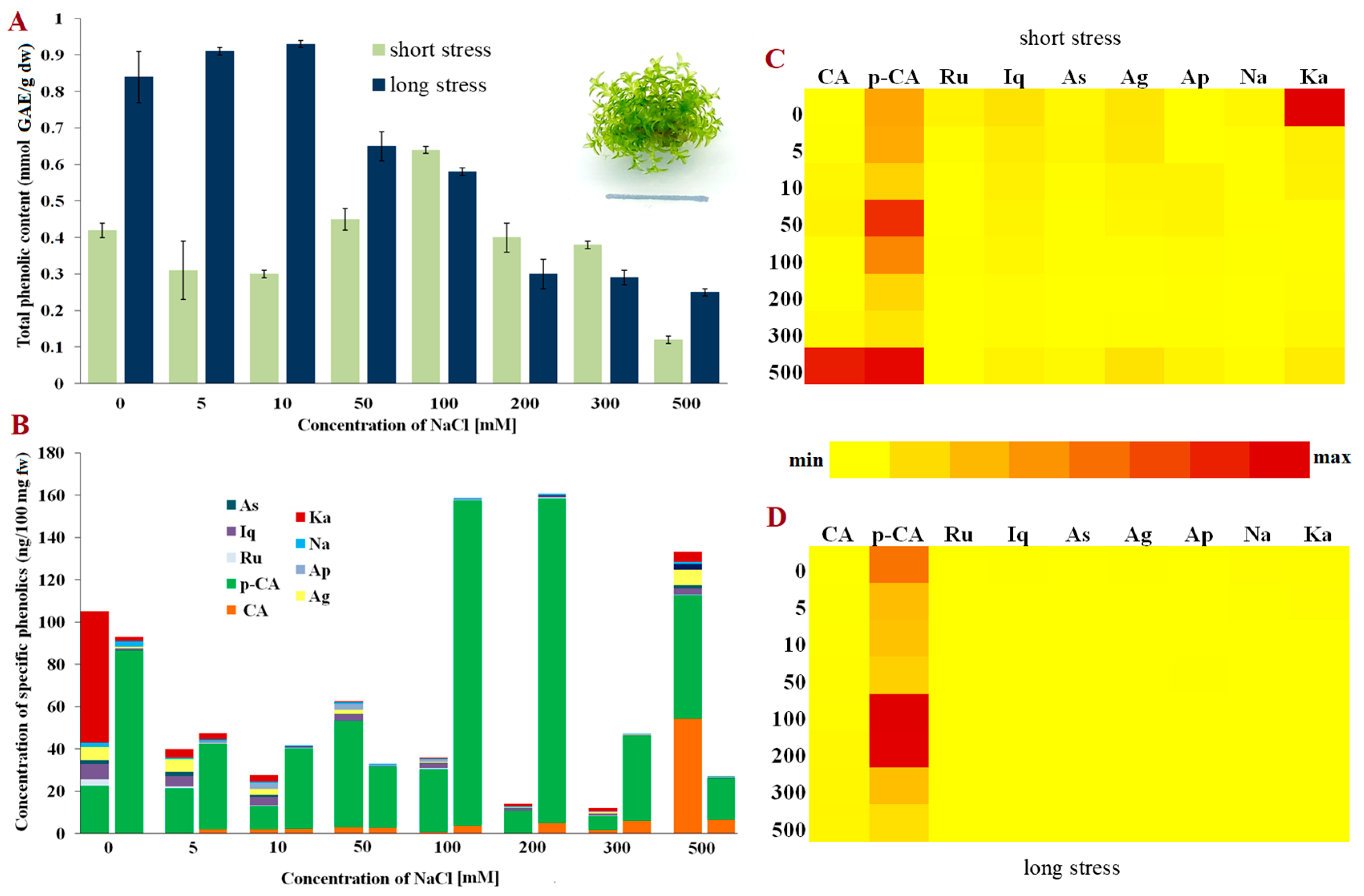{kind=link}
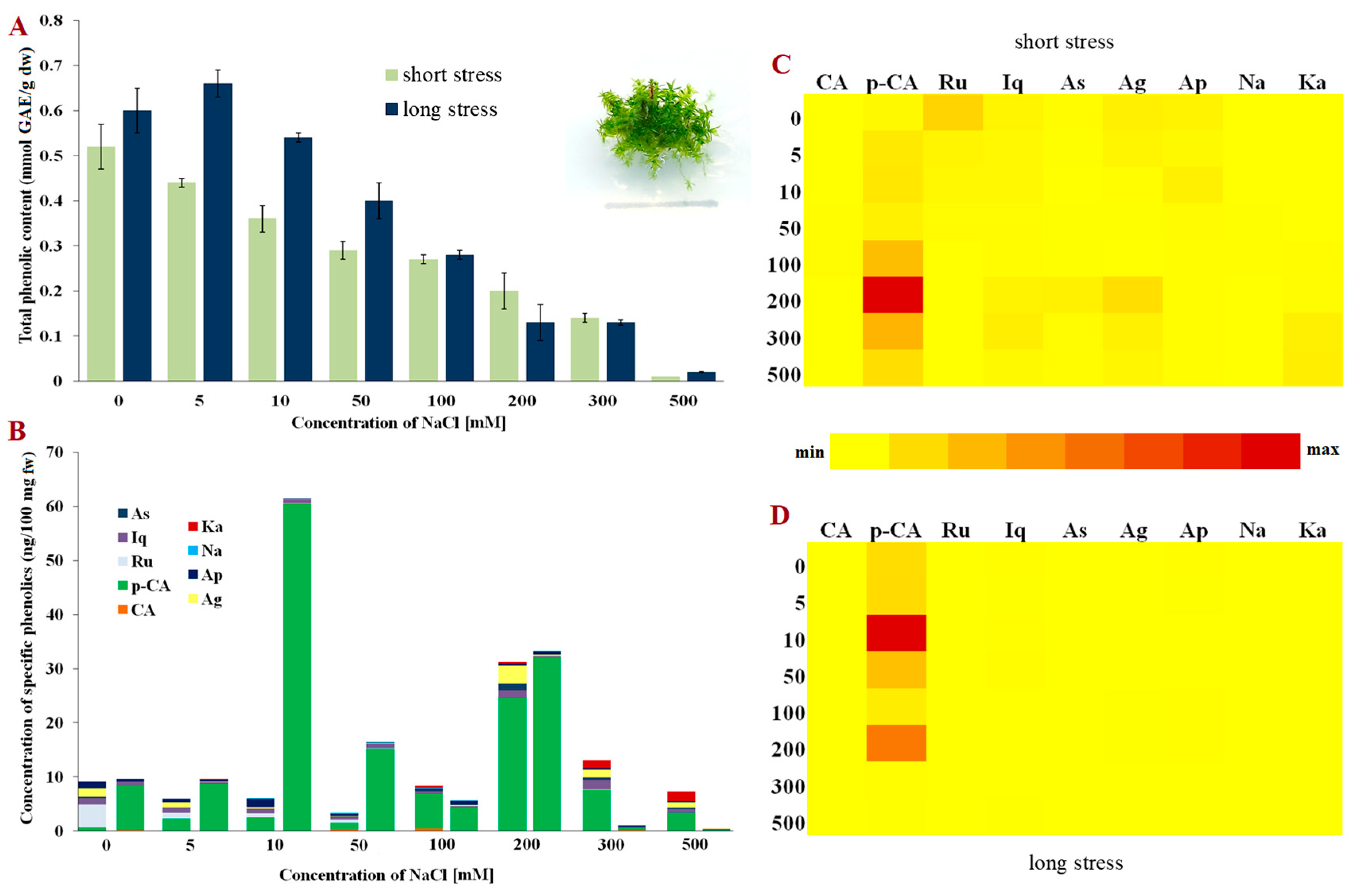{kind=link}
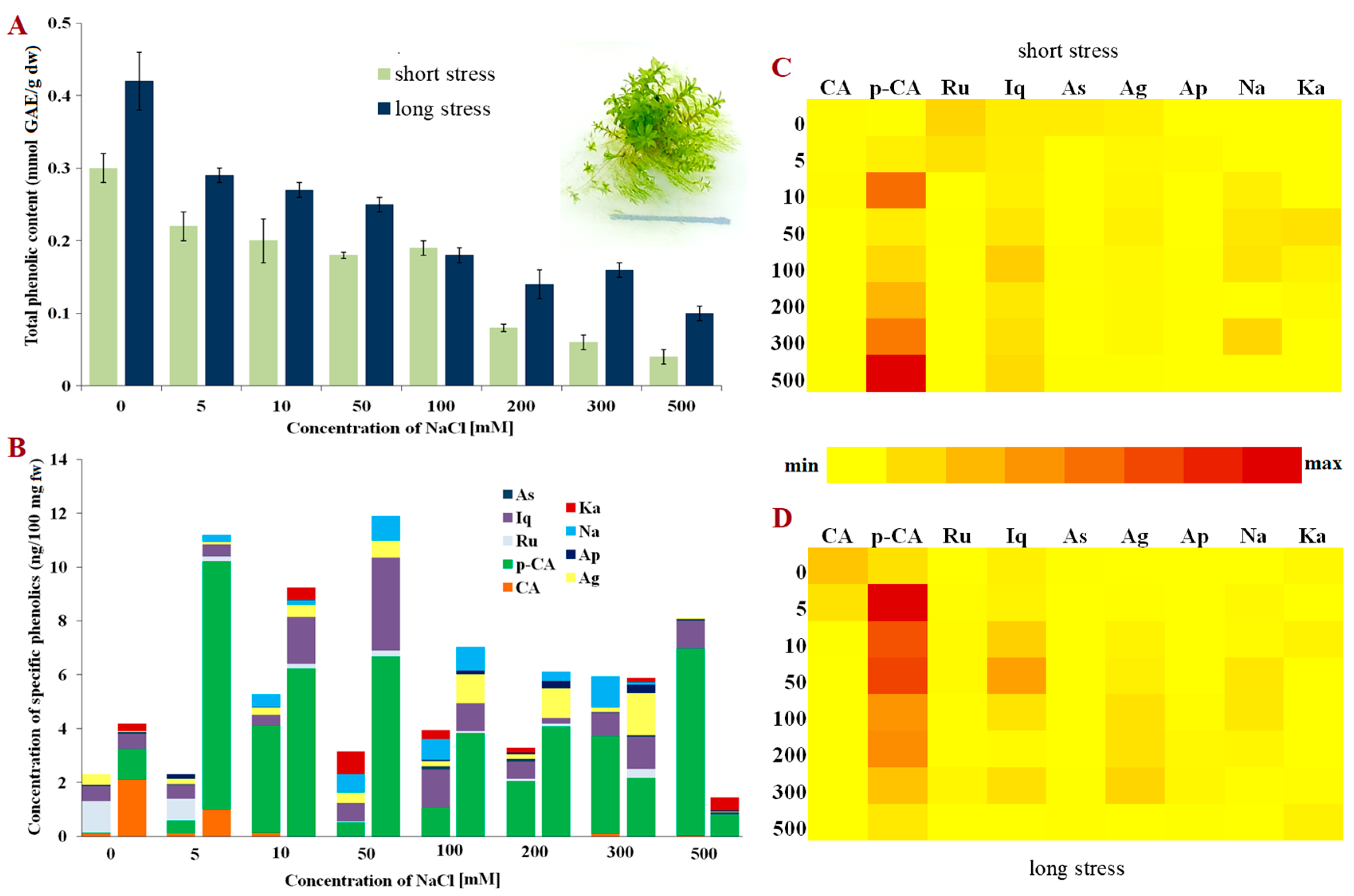{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Quantification of the Total Phenolic Content
4.3. UHPLC/-HESI-MS/MS Analysis of the Phenolic Compounds
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Hodgetts, N.; Calix, M.; Englefield, E.; Fettes, N.; Garcia Criado, M.; Patin, L.; Nieto, A.; Bergamini, A.; Bisang, I.; Baisheva, E.; et al. A Miniature World in Decline: European Red List of Mosses, Liverworts and Hornworts; IUCN: Brussels, Belgium, 2019; ISBN 9782831719931/9782831719948. [Google Scholar]
- Ćosić, M.; Vujičić, M.M.; Sabovljević, M.S.; Sabovljević, A.D. What do we know on salt stress in bryophytes? Plant Biosys. 2019, 153, 478–489. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Sabovljević, M.; Sabovljević, A. Contribution to the coastal bryophytes of the Northern Mediterranean: Are there halophytes among bryophytes? Phytol. Balc. 2007, 13, 131–135. [Google Scholar]
- Sabovljević, M.; Papp, B.; Sabovljević, A.; Vujičić, M.; Szurdoki, E.; Segarra-Moragues, J.G. In vitro micropropagation of rare and endangered moss Entosthodon hungaricus (Funariaceae). Biosci. J. 2012, 28, 632–640. [Google Scholar]
- Cuming, A.C.; Cho, S.H.; Kamisugi, Y.; Graham, H.; Quatrano, R.S. Microarray analysis of transcriptional responses to abscisic acid and osmotic, salt, and drought stress in the moss, Physcomitrella patens. New Phytol. 2007, 176, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Richardt, S.; Timmerhaus, G.; Lang, D.; Qudeimat, E.; Correa, L.; Reski, R.; Rensing, S.A.; Frank, W. Microarray analysis of the moss Physcomitrella patens reveals evolutionarily conserved transcriptional regulation of salt stress and abscisic acid signaling. Plant Mol. Biol. 2010, 72, 27–45. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Yang, P. Proteomic studies of the abiotic stresses response in model moss—Physcomitrella patens. Front. Plant Sci. 2012, 3, 258. [Google Scholar] [CrossRef]
- Gao, C.; Xing, D.; Li, L.; Zhang, L. Implication of reactive oxygen species and mitochondrial dysfunction in the early stages of plant programmed cell death induced by ultraviolet-C overexposure. Planta 2008, 227, 755–767. [Google Scholar] [CrossRef]
- Nagae, M.; Nakata, M.; Takahashi, Y. Identification of negative cis-acting elements in response to copper in the chloroplastic iron superoxide dismutase gene of the moss Barbula unguiculata. Plant Physiol. 2008, 146, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, Y. Biologically active compounds from bryophytes. Pure Appl. Chem. 2007, 79, 557–580. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Vujičić, M.; Wang, X.; Garraffo, M.; Bewley, C.A.; Sabovljević, A. Production of the macrocyclic bis-bibenzyls in axenically farmed and wild liverwort Marchantia polymorpha L. subsp. ruderalis Bischl. et Boisselier. Plant Biosyst. 2017, 151, 414–418. [Google Scholar] [CrossRef]
- Gathori, D.; Chaturvedi, P. Bryophytes: A potential source of antioxidants. In Bryophytes; Sabovljević, M.S., Sabovljević, A.D., Eds.; IntechOpen: London, UK, 2019; pp. 53–64. [Google Scholar] [CrossRef] [Green Version]
- Ludwiczuk, A.; Asakawa, Y. Bryophytes as a source of bioactive volatile terpenoids—A review. Food Chem. Toxicol. 2019, 132, 110649. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, Y. Bryophytes: Chemical diversity, synthesis and biotechnology. A review. Flavour Fragr. J. 2011, 26, 318–320. [Google Scholar] [CrossRef]
- He, X.; Sun, Y.; Zhu, R.-L. The oil bodies of liverworts: Unique and important organelles in land plants. Crit. Rev. Plant Sci. 2013, 32, 293–302. [Google Scholar] [CrossRef]
- Resemann, H.; Lewandowska, M.; Mann, J.G.; Feussner, I. Membrane lipids, waxes and oxylipins in the moss model organism Physcomitrella patens. Plant Cell Physiol. 2019, 60, 1166–1175. [Google Scholar] [CrossRef]
- Ćosić, M.V.; Janošević, D.A.; Oaldje, M.M.; Vujičić, M.M.; Lang, I.; Sabovljević, M.S.; Sabovljević, A.D. Terpenoid evidences within three selected bryophyte species under salt stress as inferred by histochemical analyses. Flora 2021, 285, 151956. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J.D. Bioactivities of phenolics by focusing on suppression of chronic diseases: A review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef] [PubMed]
- Chobot, V.; Kubicová, L.; Nabbout, S.; Jahodář, L.; Vytlačilova, J. Antioxidant and free radical scavenging activities of five moss species. Fitoterapia 2006, 77, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F.; Toyota, M.; Hashimoto, T.; Tori, M.; Fukuyama, Y.; Harinantenaina, L. Bryophytes: Bio- and chemical diversity, bioactivity and chemosystematics. Heterocycles 2009, 77, 99–150. [Google Scholar] [CrossRef] [PubMed]
- Sgherri, C.; Cosi, E.; Navarri-Izzo, F. Phenols and antioxidative status of Raphanus sativus grown in copper excess. Physiol. Plant. 2003, 118, 21–28. [Google Scholar] [CrossRef]
- Aslanbaba, B.; Yilmaz, S.; Tonguc-Yayinta, Ö. Total phenol content and antioxidant activity of mosses from Yenice forest (Ida mountain). J. Sci. Perspect. 2017, 1, 1–12. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Joganath, I.B.; Crozier, A. Dietary flavonoids and phenolic compounds. In Plant Phenolics and Human Health; Fraga, G., Ed.; Wiley: Hoboken, NJ, USA, 2010; pp. 1–50. [Google Scholar]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Bogdanović, M.; Ilić, M.; Živković, S.; Sabovljević, A.; Grubišić, D.; Sabovljević, M. Comparative study on the effects of NaCl on selected moss and fern representatives. Aust. J. Bot. 2011, 59, 734–740. [Google Scholar] [CrossRef]
- Sharma, A.; Slathia, S.; Gupta, D.; Handa, N.; Choudhary, S.P.; Langer, A.; Bhardwaj, R. Antifungal and antioxidant profile of ethnomedicinally important liverworts (Pellia endivaefolia and Plagiochasma appendiculatum) used by indigenous tribes of district Reasi: North West Himalayas. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 85, 571–579. [Google Scholar] [CrossRef]
- Ćosić, M.; Vujičić, M.M.; Sabovljević, M.S.; Sabovljević, A.D. Effects of salt on selected bryophyte species tested under controlled conditions. Bot. Serb. 2020, 44, 27–35. [Google Scholar] [CrossRef]
- Ćosić, M.; Vujičić, M.M.; Sabovljević, M.S.; Sabovljević, A.D. Effects of ABA and NaCl on physiological responses in selected bryophyte species. Botany 2020, 98, 639–650. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Nikolić, N.; Vujičić, M.; Sinžar-Sekulić, J.; Pantović, J.; Papp, B.; Sabovljević, A. Ecology, distribution, propagation in vitro, ex situ conservation and native population strengthening of rare and threatened halophyte moss Entosthodon hungaricus in Serbia. Wulfenia 2018, 25, 117–130. [Google Scholar]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Phenols, polyphenols and tannins: An overview. In Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; Crozier, A., Ashihara, H., Eds.; Blackwell: Oxford, UK, 2006; pp. 1–24. [Google Scholar]
- Asada, K.; Takahashi, M. Production and scavenging of active oxygen in photosynthesis. In Photoinhibition; Kyle, D.J., Osmond, C.B., Arntzem, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; pp. 227–287. [Google Scholar]
- Rezazadeh, A.; Ghasemneshaz, A.; Barani, M.; Telmadarrehei, T. Effect of salinity on phenolic composition and antioxidant activity of artichoke (Cynara scolymus L.) leaves. J. Med. Plant Res. 2012, 6, 245–252. [Google Scholar] [CrossRef]
- Kim, H.J.; Fonseca, J.M.; Choi, J.H.; Kubota, C.; Kwon, D.Y. Salt in irrigation water affects the nutritional and visual properties of Romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef]
- Yuan, G.; Wang, X.; Guo, R.; Wang, Q. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts. Food Chem. 2010, 121, 1014–1019. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Muzolf-Panek, M.; Goliński, P. Phenolic content changes in plants under salt stress. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M., Prasad, M., Eds.; Springer: New York, NY, USA, 2013; pp. 283–314. [Google Scholar] [CrossRef]
- Commisso, M.; Guarino, F.; Marchi, L.; Muto, A.; Piro, A.; Degola, F. Bryo-activities: A review on how bryophytes are contributing to arsenal of natural bioactive compounds against fungi. Plants 2021, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Erxleben, A.; Gessler, A.; Vervilet-Scheebaum, M.; Reski, R. Metabolite profiling of the moss Physcomitrella patens reveals evolutionary conservation of osmoprotective substances. Plant Cell Rep. 2012, 31, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Mendez, J.; Sanz-Cabanilles, F. Cinnamic Acid Esters in Anthoceros Species. Phytochemistry 1979, 18, 1409. [Google Scholar] [CrossRef]
- Jocković, N.; Andrade, P.B.; Valentão, P.; Sabovljević, M. HPLC-DAD of Phenolics in Bryophytes Lunularia cruciata, Brachytheciastrum velutinum and Kindbergia praelonga. J. Serb. Chem. Soc. 2008, 73, 1161–1167. [Google Scholar] [CrossRef]
- Frank, W.; Ratnadewi, D.; Reski, R. Physcomitrella patens is highly tolerant against drought, salt and osmotic stress. Planta 2005, 220, 384–394. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Chemical constituents of bryophytes: Bio- and chemical diversity, biological activity, and chemosystematics. In Progress in the Chemistry of Organic Natural Products; Kinghorn, A.D., Falk, H., Kobayashi, J., Eds.; Springer: Vienna, Austria, 2013; Volume 95, pp. 25–561. [Google Scholar]
- Davies, K.M.; Jibran, R.; Zhou, Y.; Albert, N.W.; Brummell, D.A.; Jordan, B.R.; Bowman, J.L.; Schwinn, K.E. The Evolution of Flavonoid Biosynthesis: A Bryophyte Perspective. Front. Plant Sci. 2020, 11, 7. [Google Scholar] [CrossRef]
- Horn, A.; Pascal, A.; Lončarević, I.; Volpatto Marques, R.; Lu, Y.; Miguel, S.; Bourgaud, F.; Thorsteinsdóttir, M.; Cronberg, N.; Becker, J.D.; et al. Natural products from bryophytes: From basic biology to biotechnological applications. Crit. Rev. Plant Sci. 2021, 40, 191–217. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A. Distribution of cyclic and acyclic bis-bibenzyls in the marchantiophyta (liverworts), ferns and higher plants and their biological activities, biosynthesis, and total synthesis (Dedicated to Professor Dr. Ei-Ichi Negishi on the Occasion of His 77th Birthday). Heterocycles Int. J. Rev. Commun. Heterocycl. Chem. 2012, 86, 891–917. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 3, 681–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Ćosić, M.V.; Sabovljević, M.S.; Papp, B.; Giba, Z.S.; Šinzar-Sekulić, J.B.; Sabovljević, A.D.; Vujičić, M.M. Micropropagation of rare bryo-halophyte Hennediella heimii. Bot. Serb. 2022, 46, 187–195. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Ćosić, M.V.; Jadranin, B.Z.; Pantović, J.P.; Giba, Z.S.; Vujičić, M.M.; Sabovljević, A.D. The conservation physiology of bryophytes. Plants 2022, 11, 1282. [Google Scholar] [CrossRef]
- Sabovljević, A.; Sabovljević, M.; Jocković, N. In vitro culture and secondary metabolite isolation in bryophytes. Methods Mol. Biol. 2009, 547, 117–128. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Mišić, D.; Šiler, B.; Gašić, U.; Avramov, S.; Živković, S.; Nestorović Živković, J.; Milutinović, M.; Tešić, Z. Simultaneous UHPLC/DAD/(+/?) HESI-MS/MS analysis of phenolic acids and nepetalactones in methanol extracts of Nepeta species: A possible application in chemotaxonomic studies. Phytochem. Anal. 2015, 26, 72–85. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ćosić, M.V.; Mišić, D.M.; Jakovljević, K.M.; Giba, Z.S.; Sabovljević, A.D.; Sabovljević, M.S.; Vujičić, M.M. Analysis of the Qualitative and Quantitative Content of the Phenolic Compounds of Selected Moss Species under NaCl Stress. Molecules 2023, 28, 1794. https://doi.org/10.3390/molecules28041794
Ćosić MV, Mišić DM, Jakovljević KM, Giba ZS, Sabovljević AD, Sabovljević MS, Vujičić MM. Analysis of the Qualitative and Quantitative Content of the Phenolic Compounds of Selected Moss Species under NaCl Stress. Molecules. 2023; 28(4):1794. https://doi.org/10.3390/molecules28041794
Chicago/Turabian StyleĆosić, Marija V., Danijela M. Mišić, Ksenija M. Jakovljević, Zlatko S. Giba, Aneta D. Sabovljević, Marko S. Sabovljević, and Milorad M. Vujičić. 2023. "Analysis of the Qualitative and Quantitative Content of the Phenolic Compounds of Selected Moss Species under NaCl Stress" Molecules 28, no. 4: 1794. https://doi.org/10.3390/molecules28041794
APA StyleĆosić, M. V., Mišić, D. M., Jakovljević, K. M., Giba, Z. S., Sabovljević, A. D., Sabovljević, M. S., & Vujičić, M. M. (2023). Analysis of the Qualitative and Quantitative Content of the Phenolic Compounds of Selected Moss Species under NaCl Stress. Molecules, 28(4), 1794. https://doi.org/10.3390/molecules28041794