

Lactose and Galactose Promote the Crystallization of Human Galectin-10

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
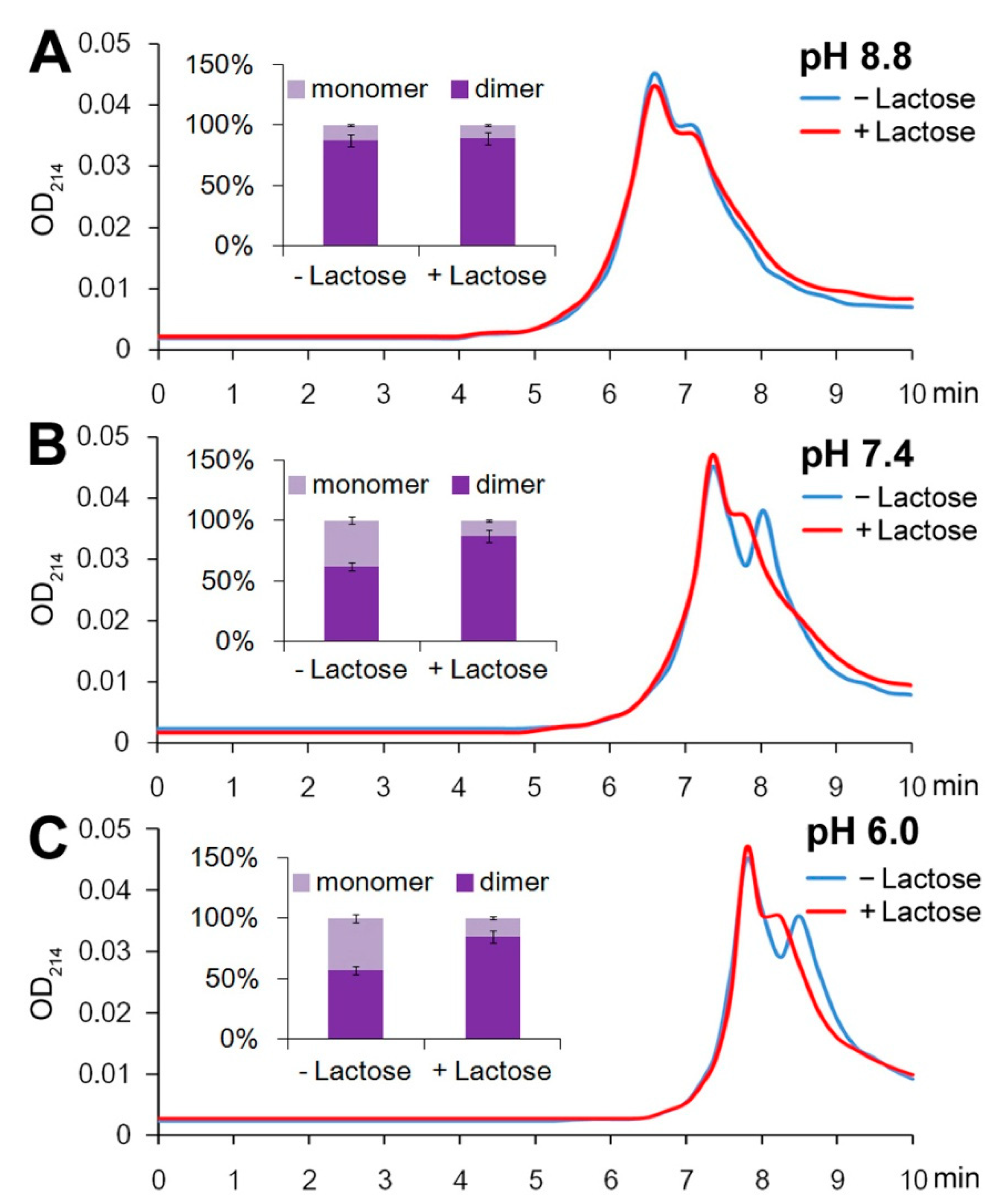
2.1. Gal-10 Is a Dimer at High Concentrations
2.2. More Dimers Formed upon Lactose/Galactose Binding
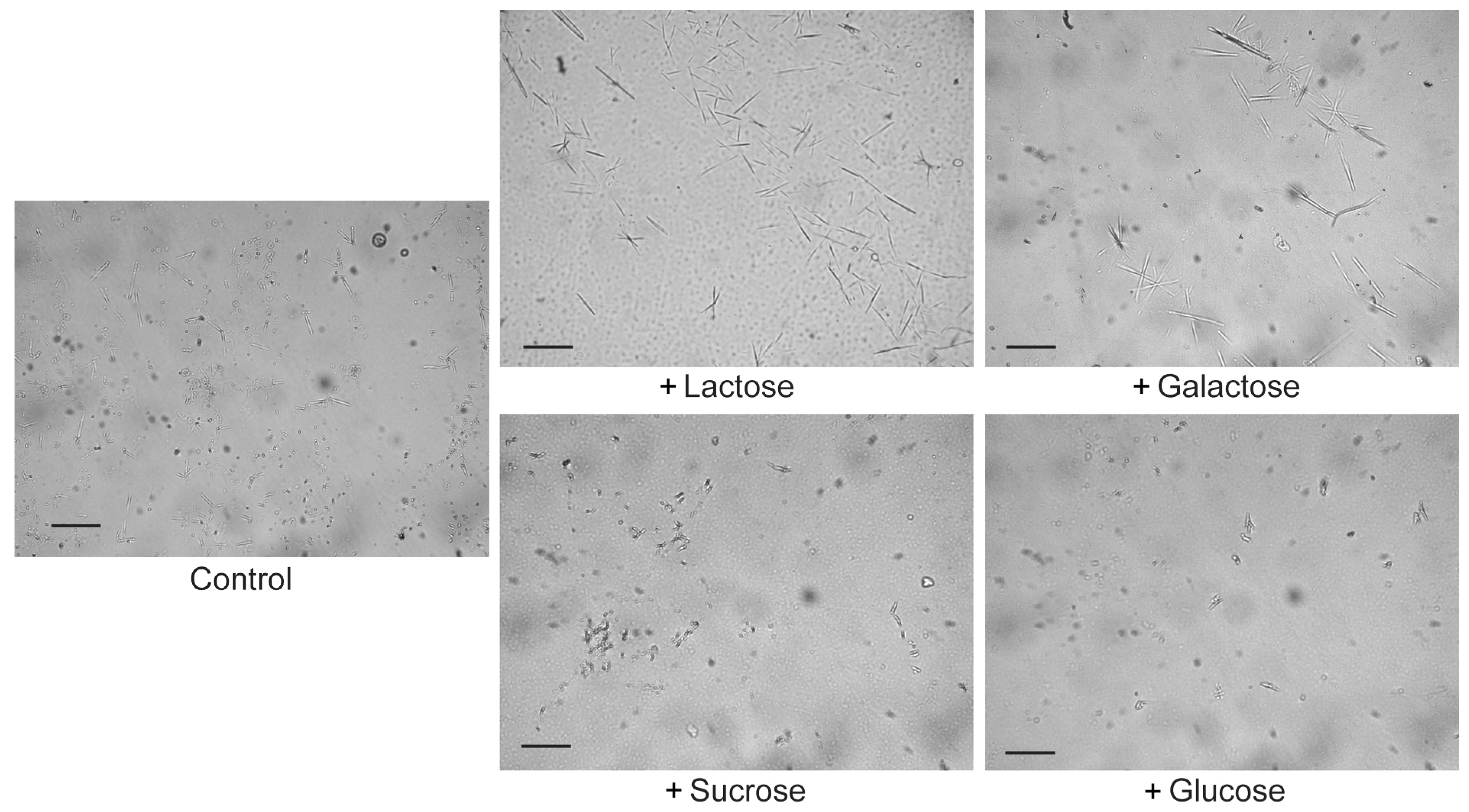
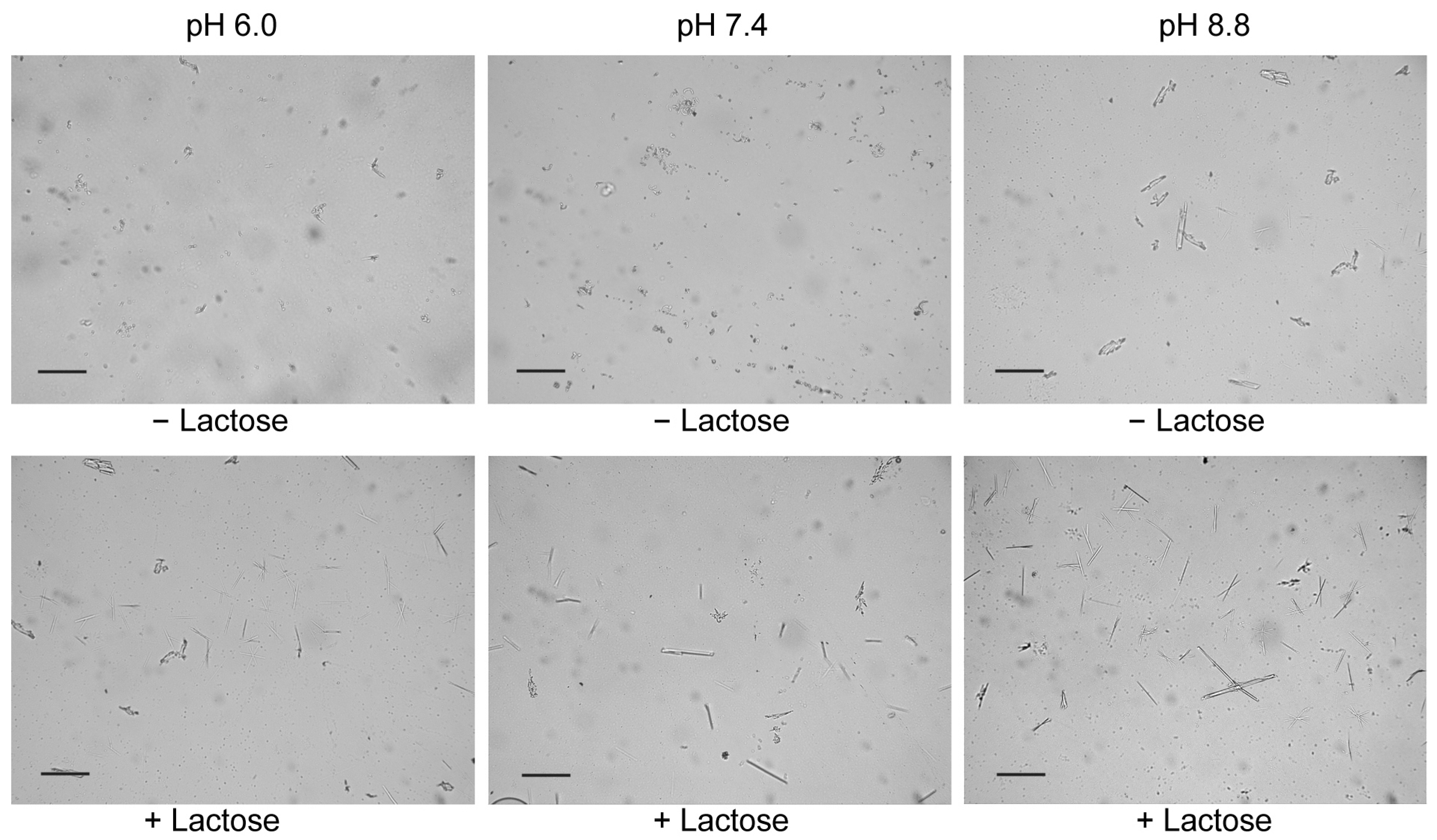
2.3. Lactose/Galactose Binding Promotes Gal-10 Crystallization
2.4. Surface Electrostatic Shift on the Gal-10 Monomer after Lactose Binding
3. Discussion
4. Materials and Methods
4.1. Human CLC Protein
4.2. Separation of the Monomers and Dimers of Gal-10 via Size-Exclusion Chromatography
4.3. Reversibility of Monomer–Dimer Equilibrium
4.4. Microscope Observation of Gal-10 Crystals
4.5. Homology Modeling of Gal-10
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Charcot, J.M.; Robin, C. Observation de leucocythemie. C. R. Seances Mem. Soc. Biol. Paris 1853, 5, 44–52. [Google Scholar]
- Leyden, E. Zur Kenntniss des Bronchial-Asthma. Virchows Arch. 1872, 54, 324–344. [Google Scholar] [CrossRef] [Green Version]
- Su, J. A brief history of Charcot-Leyden crystal protein/galectin-10 research. Molecules 2018, 23, 2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryborn, M.; Hallden, C.; Sall, T.; Cardell, L.O. CLC—A novel susceptibility gene for allergic rhinitis? Allergy 2010, 65, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Staribratova, D.; Belovejdov, V.; Staikov, D.; Dikov, D. Demonstration of Charcot-Leyden crystals in eosinophilic cystitis. Arch. Pathol. Lab Med. 2010, 134, 1420. [Google Scholar] [CrossRef]
- Noh, S.; Jin, S.; Park, C.O.; Lee, Y.S.; Lee, N.; Lee, J.; Shin, J.U.; Kim, S.H.; Yun, K.N.; Kim, J.Y.; et al. Elevated galectin-10 expression of IL-22 producing T cells in atopic dermatitis patients. J. Investig. Dermatol. 2015, 136, 328–331. [Google Scholar] [CrossRef] [Green Version]
- De Re, V.; Simula, M.P.; Cannizzaro, R.; Pavan, A.; De Zorzi, M.A.; Toffoli, G.; Canzonieri, V. Galectin-10, eosinophils, and celiac disease. Ann. N. Y. Acad. Sci. 2009, 1173, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Nyenhuis, S.M.; Alumkal, P.; Du, J.; Maybruck, B.T.; Vinicky, M.; Ackerman, S.J. Charcot-Leyden crystal protein/galectin-10 is a surrogate biomarker of eosinophilic airway inflammation in asthma. Biomark Med. 2019, 13, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Nashiro, T.; Hashimoto, H.; Horiuchi, H.; Usuki, K. Acute myeloid leukemia-associated Charcot-Leyden crystals in the bone marrow. Intern. Med. 2016, 55, 1821–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ågesen, T.H.; Berg, M.; Clancy, T.; Thiisevensen, E.; Cekaite, L.; Lind, G.E.; Nesland, J.M.; Bakka, A.; Mala, T.; Hauss, H.J.; et al. CLC and IFNAR1 are differentially expressed and a global immunity score is distinct between earlyand late-onset colorectal cancer. Genes Immun. 2011, 12, 653. [Google Scholar] [CrossRef] [Green Version]
- Lao, L.M.; Kumakiri, M.; Nakagawa, K.; Ishida, H.; Ishiguro, K.; Yanagihara, M.; Ueda, K. The ultrastructural findings of Charcot-Leyden crystals in stroma of mastocytoma. J. Dermatol. Sci. 1998, 17, 198–204. [Google Scholar] [CrossRef]
- Silver, G.K.; Simon, J.H. Charcot-Leyden crystals within a periapical lesion. J. Endod. 2000, 26, 679–681. [Google Scholar] [CrossRef]
- Taylor, G.; Ivey, A.; Milner, B.; Grimwade, D.; Culligan, D. Acute myeloid leukaemia with mutated NPM1 presenting with extensive bone marrow necrosis and Charcot-Leyden crystals. Int. J. Hematol. 2013, 98, 267–268. [Google Scholar] [CrossRef]
- Ackerman, S.J.; Corrette, S.E.; Rosenberg, H.F.; Bennett, J.C.; Mastrianni, D.M.; Nicholson-Weller, A.; Weller, P.F.; Chin, D.T.; Tenen, D.G. Molecular cloning and characterization of human eosinophil Charcot-Leyden crystal protein (lysophospholipase). Similarities to IgE binding proteins and the S-type animal lectin superfamily. J. Immunol. 1993, 150, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [CrossRef] [PubMed]
- Leonidas, D.D.; Elbert, B.L.; Zhou, Z.; Leffler, H.; Ackerman, S.J.; Acharya, K.R. Crystal structure of human Charcot-Leyden crystal protein, an eosinophil lysophospholipase, identifies it as a new member of the carbohydrate-binding family of galectins. Structure 1995, 3, 1379–1393. [Google Scholar] [CrossRef] [Green Version]
- Leffler, H.; Carlsson, S.; Hedlund, M.; Qian, Y.; Poirier, F. Introduction to galectins. Glycoconj. J. 2002, 19, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Gao, J.; Si, Y.; Cui, L.; Song, C.; Wang, Y.; Wu, R.; Tai, G.; Zhou, Y. Galectin-10: A new structural type of prototype galectin dimer and effects on saccharide ligand binding. Glycobiology 2018, 28, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Song, C.; Si, Y.; Cui, L.; Yang, T.; Li, Y.; Wang, H.; Tai, G.; Zhou, Y. Identification of key amino acid residues determining ligand binding specificity, homodimerization and cellular distribution of human galectin-10. Glycobiology 2019, 29, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Dyer, K.D.; Rosenberg, H.F. Eosinophil Charcot-Leyden crystal protein binds to beta-galactoside sugars. Life Sci. 1996, 58, 2073–2082. [Google Scholar] [CrossRef]
- Dvorak, A.M.; Furitsu, T.; Letourneau, L.; Ishizaka, T.; Ackerman, S.J. Mature eosinophils stimulated to develop in human cord blood mononuclear cell cultures supplemented with recombinant human interleukin-5. Part I. Piecemeal degranulation of specific granules and distribution of Charcot-Leyden crystal protein. Am. J. Pathol. 1991, 138, 69–82. [Google Scholar] [PubMed]
- Swaminathan, G.J.; Leonidas, D.D.; Savage, M.P.; Ackerman, S.J.; Acharya, K.R. Selective recognition of mannose by the human eosinophil Charcot-Leyden crystal protein (galectin-10): A crystallographic study at 1.8 A resolution. Biochemistry 1999, 38, 13837–13843. [Google Scholar] [CrossRef] [PubMed]
- Persson, E.K.; Verstraete, K.; Heyndrickx, I.; Gevaert, E.; Aegerter, H.; Percier, J.M.; Deswarte, K.; Verschueren, K.H.G.; Dansercoer, A.; Gras, D.; et al. Protein crystallization promotes type 2 immunity and is reversible by antibody treatment. Science 2019, 364, eaaw4295. [Google Scholar] [CrossRef] [PubMed]
- Itoh, A.; Nonaka, Y.; Nakakita, S.I.; Yoshida, H.; Nishi, N.; Nakamura, T.; Kamitori, S. Structures of human galectin-10/monosaccharide complexes demonstrate potential of monosaccharides as effectors in forming Charcot-Leyden crystals. Biochem. Biophys. Res. Commun. 2020, 525, 87–93. [Google Scholar] [CrossRef]
- Cho, M.; Cummings, R.D. Galectin-1, a beta-galactoside-binding lectin in Chinese hamster ovary cells. I. Physical and chemical characterization. J. Biol. Chem. 1995, 270, 5198–5206. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, S.J.; Liu, L.; Kwatia, M.A.; Savage, M.P.; Leonidas, D.D.; Swaminathan, G.J.; Acharya, K.R. Charcot-Leyden crystal protein (galectin-10) is not a dual function galectin with lysophospholipase activity but binds a lysophospholipase inhibitor in a novel structural fashion. J. Biol. Chem. 2002, 277, 14859–14868. [Google Scholar] [CrossRef] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Bertoni, M.; Kiefer, F.; Biasini, M.; Bordoli, L.; Schwede, T. Modeling protein quaternary structure of homo- and hetero-oligomers beyond binary interactions by homology. Sci. Rep. 2017, 7, 10480. [Google Scholar] [CrossRef] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.P.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository—New features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Svergun, D.; Barberato, C.; Koch, M.H.J. CRYSOL—A program to evaluate X-ray solution scattering of biological macromolecules from atomic coordinates. J. Appl. Crystallogr. 1995, 28, 768–773. [Google Scholar] [CrossRef]
- Rambo, R.P.; Tainer, J.A. Accurate assessment of mass, models and resolution by small-angle scattering. Nature 2013, 496, 477–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, E.A.; Jackson, D.J.; Heffler, E.; Mathur, S.K.; Bredenoord, A.J.; Pavord, I.D.; Akuthota, P.; Roufosse, F.; Rothenberg, M.E. Eosinophil knockout humans: Uncovering the role of eosinophils through eosinophil-directed biological therapies. Annu. Rev. Immunol. 2021, 39, 719–757. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W.; Sedgwick, J.B. Eosinophils in asthma. Ann. Allergy 1992, 68, 286–290. [Google Scholar] [PubMed]
- Welsh, R.A. The genesis of the Charcot-Leyden crystal in the eosinophilic leukocyte of man. Am. J. Pathol. 1959, 35, 1091–1103. [Google Scholar]
- Dor, P.J.; Ackerman, S.J.; Gleich, G.J. Charcot-Leyden crystal protein and eosinophil granule major basic protein in sputum of patients with respiratory diseases. Am. Rev. Respir. Dis. 1984, 130, 1072–1077. [Google Scholar] [CrossRef]
- Aegerter, H.; Smole, U.; Heyndrickx, I.; Verstraete, K.; Savvides, S.N.; Hammad, H.; Lambrecht, B.N. Charcot-Leyden crystals and other protein crystals driving type 2 immunity and allergy. Curr. Opin. Immunol. 2021, 72, 72–78. [Google Scholar] [CrossRef]
- Fulkerson, P.C.; Rothenberg, M.E. Targeting eosinophils in allergy, inflammation and beyond. Nat. Rev. Drug Discov. 2013, 12, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udell, I.J.; Gleich, G.J.; Allansmith, M.R.; Ackerman, S.J.; Abelson, M.B. Eosinophil granule major basic protein and Charcot-Leyden crystal protein in human tears. Am. J. Ophthalmol. 1981, 92, 824–828. [Google Scholar] [CrossRef]
- Ghafouri, B.; Irander, K.; Lindbom, J.; Tagesson, C.; Lindahl, M. Comparative proteomics of nasal fluid in seasonal allergic rhinitis. J. Proteome Res. 2006, 5, 330–338. [Google Scholar] [CrossRef]
- Arroyo-Hornero, R.; Aegerter, H.; Hamad, I.; Corte-Real, B.; Staes, K.; van der Woning, B.; Verstraete, K.; Savvides, S.N.; Lambrecht, B.N.; Kleinewietfeld, M. The Charcot-Leyden crystal protein galectin-10 is not a major determinant of human regulatory T-cell function. Allergy 2022, 77, 2818–2821. [Google Scholar] [CrossRef]
- Muir, A.B.; Ackerman, S.J.; Pan, Z.; Benitez, A.; Burger, C.; Spergel, J.M.; Furuta, G.T.; Rothman, J.; Wilkins, B.J.; Arnold, M.A.; et al. Esophageal remodeling in eosinophilic esophagitis: Relationships to luminal captured biomarkers of inflammation and periostin. J. Allergy Clin. Immunol. 2022, 150, 649–656.e5. [Google Scholar] [CrossRef]
- Gelardi, M.; Netti, G.S.; Giancaspro, R.; Spadaccino, F.; Pennella, A.; Fiore, V.; La Gatta, E.; Grilli, G.M.; Cassano, M.; Ranieri, E. Chronic rhinosinusitis with nasal polyposis (CRSwNP): The correlation between expression of Galectin-10 and Clinical-Cytological Grading (CCG). Am. J. Rhinol. Allergy 2022, 36, 229–237. [Google Scholar] [CrossRef]
- Chen, W.; Bai, Y.; Kong, W.; Luo, X.; Zeng, Y.; Chen, J.; Wang, X.; Wu, Q.; Chaudhuri, S.; Chen, J.; et al. Predictive significance of Charcot-Leyden crystal structures for nasal polyp recurrence. Clin. Transl. Allergy 2022, 12, e12212. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.C.N.; Wang, H.; Silva, T.P.; Imoto, Y.; Fujieda, S.; Fukuchi, M.; Miyabe, Y.; Hirokawa, M.; Ueki, S.; Weller, P.F. Galectin-10, the protein that forms Charcot-Leyden crystals, is not stored in granules but resides in the peripheral cytoplasm of human eosinophils. J. Leukoc. Biol. 2020, 108, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.R.; Tajudeen, B.A.; Brown, H.; Gattuso, P.; LoSavio, P.; Papagiannopoulos, P.; Batra, P.S.; Mahdavinia, M. Association of air pollutant exposure and sinonasal histopathology findings in chronic rhinosinusitis. Am. J. Rhinol. Allergy 2021, 35, 761–767. [Google Scholar] [CrossRef]
- Guo, C.L.; Wang, C.S.; Liu, Z. Clinical and biological markers in disease and biologics to treat chronic rhinosinusitis. Curr. Opin. Allergy Clin. Immunol. 2022, 22, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Delemarre, T.; Holtappels, G.; De Ruyck, N.; Zhang, N.; Nauwynck, H.; Bachert, C.; Gevaert, E. A substantial neutrophilic inflammation as regular part of severe type 2 chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2021, 147, 179–188.e2. [Google Scholar] [CrossRef]
- Poczobutt, J.M.; Mikosz, A.M.; Poirier, C.; Beatman, E.L.; Serban, K.A.; Gally, F.; Cao, D.; McCubbrey, A.L.; Cornell, C.F.; Schweitzer, K.S.; et al. Altered macrophage function associated with crystalline lung inflammation in acid sphingomyelinase deficiency. Am. J. Respir. Cell. Mol. Biol. 2021, 64, 629–640. [Google Scholar] [CrossRef]
- Weller, P.F.; Wang, H.; Melo, R.C.N. The Charcot-Leyden crystal protein revisited-A lysopalmitoylphospholipase and more. J. Leukoc. Biol. 2020, 108, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Grozdanovic, M.M.; Doyle, C.B.; Liu, L.; Maybruck, B.T.; Kwatia, M.A.; Thiyagarajan, N.; Acharya, K.R.; Ackerman, S.J. Charcot-Leyden crystal protein/galectin-10 interacts with cationic ribonucleases and is required for eosinophil granulogenesis. J. Allergy Clin. Immunol. 2020, 146, 377–389.e10. [Google Scholar] [CrossRef] [Green Version]
- Frosh, A.; Cruz, C.; Wellsted, D.; Stephens, J. Effect of a dairy diet on nasopharyngeal mucus secretion. Laryngoscope 2019, 129, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Maslova, E.; Halldorsson, T.I.; Strøm, M.; Olsen, S.F. Low-fat yoghurt intake in pregnancy associated with increased child asthma and allergic rhinitis risk: A prospective cohort study. J. Nutr. Sci. 2012, 1, e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Nagase, H.; Sugimoto, N.; Yamamoto, S.; Tanaka, A.; Fukunaga, K.; Atsuta, R.; Tagaya, E.; Hojo, M.; Gon, Y.; et al. Mepolizumab decreased the levels of serum galectin-10 and eosinophil cationic protein in asthma. Asia Pac. Allergy 2021, 11, e31. [Google Scholar] [CrossRef] [PubMed]
- Valadez-Vega, C.; Lugo-Magaña, O.; Betanzos-Cabrera, G.; Villagómez-Ibarra, J.R. Partial characterization of lectins purified from the Surco and Vara (Furrow and Rod) varieties of black Phaseolus vulgaris. Molecules 2022, 27, 8436. [Google Scholar] [CrossRef]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, T.; Chen, K.; McLaughlin, W.A.; Lu, B.; Wang, W. Computational analysis and prediction of the binding motif and protein interacting partners of the Abl SH3 domain. PLoS Comput. Biol. 2006, 2, e1. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.-F.; Jiang, S.-C.; Zhang, Z.-W.; Yang, X.-Y.; Li, Z.-L.; Hu, J.; Yuan, S. Lactose and Galactose Promote the Crystallization of Human Galectin-10. Molecules 2023, 28, 1979. https://doi.org/10.3390/molecules28041979
Fu Y-F, Jiang S-C, Zhang Z-W, Yang X-Y, Li Z-L, Hu J, Yuan S. Lactose and Galactose Promote the Crystallization of Human Galectin-10. Molecules. 2023; 28(4):1979. https://doi.org/10.3390/molecules28041979
Chicago/Turabian StyleFu, Yu-Fan, Si-Cong Jiang, Zhong-Wei Zhang, Xin-Yue Yang, Zi-Lin Li, Jing Hu, and Shu Yuan. 2023. "Lactose and Galactose Promote the Crystallization of Human Galectin-10" Molecules 28, no. 4: 1979. https://doi.org/10.3390/molecules28041979
APA StyleFu, Y. -F., Jiang, S. -C., Zhang, Z. -W., Yang, X. -Y., Li, Z. -L., Hu, J., & Yuan, S. (2023). Lactose and Galactose Promote the Crystallization of Human Galectin-10. Molecules, 28(4), 1979. https://doi.org/10.3390/molecules28041979